











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El conocimiento relacionado con las lombrices de tierra del neotrópico es todavía incipiente. En América del Sur se ha avanzado, pero se conoce poco de la presencia y distribución de muchas de las familias que se han reconocido, especialmente Rhinodrilidae. Esta familia, cuenta con la mayoría de especies nativas y se caracteriza por la presencia de 1 molleja en el segmento 6, un número variable de glándulas calcíferas en cada género, oscilan desde 1 par hasta 8 y que se localizan después de la molleja y adheridas al esófago. En el caso de la región amazónica (Amazonía sensu-stricto; Eva y Huber, 2005) se han encontrado 202 especies nativas, de las cuales la mayoría fueron colectadas en Brasil (124) (Feijoo et al., 2017), mientras que en Colombia los valores siguen siendo bajos (14 especies incluyendo las 5 de este estudio) debido a las pocas colectas realizadas en la Amazonía colombiana. Uno de los géneros de Rhinodrilidae que se destaca por el alto número de especies es Rhinodrilus Perrier, 1872, holándrico con 3 pares de glándulas calcíferas entre los segmentos 7-9, para el que se conocen 49 especies descritas. De ellas, 27 se han encontrado en Brasil (10 amazónicas), 11 en Venezuela (2 de Amazonía), 3 amazónicas en Perú, 4 amazónicas en Colombia (este estudio), 3 en Trinidad (una de ellas con distribución compartida con Venezuela) y 1 en Argentina. Otro género importante es Diachaeta (Benham, 1887), metándrico con 3 pares de glándulas calcíferas entre los segmentos 7-9 y que, a su vez, se subdivide en los subgéneros Amazo con 12 especies y Diachaeta con 9. Diachaeta (Amazo) se distribuye en Brasil (7 especies en la Amazonía y 1 en el Mato Grosso), Venezuela (3 en Amazonía), 1 amazónica en Colombia (este estudio) y 1 en Argentina. Diachaeta (Diachaeta) se ha encontrado en Brasil (3 especies amazónicas) y en las Antillas menores (4 distribuidas en Barbados, Bonaire y Jamaica) (Brown y Fragoso, 2007).
De acuerdo con los registros para las lombrices de tierra de América Latina, hasta el momento no se habían registrado especies de los géneros Rhinodrilus y Diachaeta en Colombia (Brown y Fragoso, 2007; Brown et al., 2013; Feijoo y Celis, 2010, 2011, 2012). Por lo anterior, en este artículo se describen por primera vez para Colombia 5 especies de estos 2 géneros provenientes de la Amazonía colombiana.
Materiales y métodos
El área de muestreo se ubicó en la llamada “tierra firme” caracterizada por suelos no inundables del orden de los Inceptisoles y Oxisoles (USDA, 1999), de los departamentos de Amazonas (municipio de Leticia, corregimientos de La Chorrera, Puerto Arica y Puerto Alegría), Putumayo (municipio de Puerto Leguízamo) y Caquetá (municipios de Florencia y Morelia). En los sitios se utilizó la metodología Tropical Soil Biology and Fertility-TSBF (Anderson e Ingram, 1993), con colectas manuales en el mantillo y en los estratos 0-10, 10-20, 20-30 cm de profundidad en 5 monolitos de 25 × 25 × 30 cm por cada lugar distribuidos al azar en un área de 60 × 60 m2.
Las lombrices fueron separadas manualmente en recipientes plásticos, para luego ser fijadas y conservadas en alcohol al 95%. Los especímenes fueron examinados mediante observación externa utilizando un estereomicroscopio Nikon (SMZ800), se usaron números arábigos para ubicar los segmentos, intersegmentos y septos internos o disepimentos, mismos que fueron enumerados desde el peristomio hasta el periprocto en la región posterior. La longitud fue medida, el diámetro antes del clitelo (región preclitelar, segmento 8), dentro del clitelo (región intraclitelar, segmentos 18 al 20) y después del clitelo (posclitelar, segmentos 35 al 40) con calibrador milimétrico (Feijoo, 2008; Feijoo y Celis, 2010, 2011, 2012). Posteriormente, se disectó el individuo sobre la región dorsal para ver estructuras internas tales como: la posición, estructura y forma de los disepimentos, espermatecas, glándulas calcíferas, genitales, próstatas, nefridios y para extraer las cerdas común y genital. Se usó el ocular milimétrico con precisión de 0.01 mm para medir la distancia entre las cerdas y establecer el tamaño de las estructuras con relación al plano de observación.
Para cada individuo se describieron las estructuras que conforman la morfología externa (color, forma, tamaño, diámetro, presencia poros, tipo de cerdas, posición del clitelo y bandas de la pubertad) e interna (número y posición de espermatecas, vesículas seminales y glándulas calcíferas, aparato reproductor masculino y femenino, tipo de nefridios, etc.). Para la ubicación de la familia, género, subgénero y especie, se utilizaron las claves comparativas de Feijoo (2008), Feijoo y Celis (2010, 2011, 2012). Los holotipos y paratipos fueron depositados en la Colección Nacional de Lombrices de Tierra de la Universidad Tecnológica de Pereira (CNLTUTP), Pereira, Colombia.
Descripción
Rhinodrilidae (Benham, 1890) (emend James, 2012) Género Rhinodrilus (Perrier, 1872)
Diagnosis. Ocho hileras de cerdas por segmento dispuestas en series longitudinales regulares, raramente irregulares en la región posterior. Un par de poros masculinos intraclitelares, poco frecuente 2 pares. Una molleja en el segmento 6. Tres pares de glándulas calcíferas en los segmentos 7-9, con estructura tubular dicotómica (paniculada) o tubular compuesta. Dos pares de testículos e infundíbulos seminales en los segmentos 10 y 11. En pocos casos se presentan glándulas prostáticas. Un par de ovarios en el segmento 13. Dos a 8 pares de espermatecas presentes entre los segmentos 6-7 y 14-15, intracelómicas, raramente intraparietales.
Especie tipo: Rhinodrilus paradoxus (Perrier, 1872). Número de especies del género: 49. Distribución: Brasil (29), Venezuela (11), Colombia (4), Guyana Francesa (3), Trinidad (2), Perú (1) y Argentina (1). En algunos casos se ha encontrado 1 especie en 2 países (Brown y Fragoso, 2007).
Rhinodrilus alecrisus sp. nov.
http://www.zoobank.org/urn:lsid:zoobank.org:act:ADE19F9C-1F9D-44A2-BCC0-F6CB9ADC32EB

(fotografía: A. Feijoo).
Figura 1 Rhinodrilus alecrisus, sp. nov. Holotipo. CNLT-UTP-0310. A. Cerda común, segmento 38; B, muestra cerda genital a, segmento 19; C, región ventral del clitelo; D, espermateca del segmento 7; E, glándulas prostática lado izquierdo, segmentos 19/20. P: Próstata.
Descripción. Especie epigea. Caracteres externos: forma del cuerpo cilíndrica. El color del ejemplar es café en el dorso, marrón en el clitelo y amarillo claro en la región ventral. La longitud del holotipo es 85.8 mm, con diámetro en la región anteroclitelar de 3.34 mm. Número de segmentos: 162. Tres paratipos: longitud 64.4 mm y ancho 3.0 mm, 89.6, 3.1; 87.4, 3.2, respectivamente; número de segmentos: 120, 128, 136, respectivamente. Los segmentos son visibles a partir del primero y el prostomio es alargado y con forma de lóbulo. Ocho hileras de cerdas estrechamente pareadas. Las cerdas ab son visibles a partir del segmento 7. La cerda común mide 0.6 mm y tiene forma de gancho (Fig. 1A). La relación entre las cerdas entre los segmentos 35-40 es aa:ab:bc:cd:dd = 5.6:1.0:8.3:0.7:25.7. Las cerdas ab de los segmentos 19-20 se transforman en genitales, las cuales miden 1,2 mm, están recubiertas por folículos largos, son rectas, con la punta en forma de broca y con la ornamentación presentando excavaciones cubiertas por gránulos con forma ovalada (Fig. 1B).
El clitelo, con forma de silla, ocupa los segmentos 16-24 (= 9) y en el campo genital masculino se localiza una fosa puberal en el intercepto de 19/20 en medio de las cerdas aa (Fig. 1C). Los poros masculinos se localizan en 19/20, mientras que los poros femeninos se visualizaron en 14/15. Los nefridioporos en línea con las cerdas c, d. Un par de poros de las espermatecas en 8/9. Un par de poros prostáticos se presentan en el intersegmento de 19/20.
Caracteres internos: los septos de los segmentos 6/7 - 9/10 son gruesos, cónicos e interpenetrados. La molleja redonda y muy musculosa se sitúa en el segmento 6. Tres pares de glándulas calcíferas de estructura tubular dicotómica se localizan en cada uno de los segmentos 7, 8, 9. La transición esófago-intestino comienza en los segmentos 21/22 y el tiflosol inicia en el intersegmento 24/25. No hay ciego intestinal. Tres pares de corazones laterales en 7-9 y 3 pares de corazones intestinales se sitúan en los segmentos 10, 11, 12. Hay 1 par de holonefrídios por segmento. Dos pares de sacos testiculares periesofágicos se localizaron en 10 y 11. Las vesículas seminales pares en los segmentos 11 y 12. Un par de conductos deferentes ingresan a cada próstata en los segmentos 19-20, y de allí, van en dirección del poro masculino para desembocar al exterior. El par de próstatas tiene forma de racimo, color blanquecino y se encuentra en el intersegmento 19/20 (Fig. 1E). Dos ovarios con color brillante se presentan en la cavidad del segmento 13 y el poro femenino se visualizó en el intersegmento de 14/15; los embudos femeninos no se observaron. Se situó 1 par de espermatecas con forma tubular en el segmento 8, que atraviesa 2 anillos y termina en 10 (Fig. 1D).
Resumen taxonómico
Material examinado. Material tipo. Holotipo: clitelado (CNLT-UTP-0310). Colombia, departamento de Amazonas: Corregimiento de Puerto Alegría (00°59’34.3” S, 74°01’10.4” O). Hábitat: lomeríos con bosque secundario maduro, sin uso reciente, altitud 100 m snm, 10 cm profundidad. 10 de julio de 2016. Paratipos: (8) (CNLTUTP-0311). Colombia; departamento del Putumayo: puerto Leguízamo (00°08’42.1” S, 74°46’40.9” O), lomeríos con bosque secundario maduro intervenido, sin uso reciente. Altitud 130 m snm, 11 de julio de 2016. Cols.: D. Castro, O. Ramírez y C. Peña-Venegas; departamento del Caquetá, Morelia, sector de Lagunilla (01°26’28.8” S, 75°39’10.3” O), lomeríos con relictos de selva maduro moderadamente intervenido, sin uso reciente. Altitud 150 m snm, 10 cm de profundidad, 19 de marzo de 2016; departamento del Caquetá: Florencia, sector Doradas Altas (01°42’29.8” S, 75°41’32.4” O), montaña con potreros en barbecho con más de 5 años sin uso. Altitud: 1,000 m snm, 6 de abril de 2016. Cols.: Y. Virguez y C. Peña-Venegas.
Etimología. El nombre específico fue dado como tributo al corregimiento del departamento de Amazonas (latín alecris = alegría).
Comentarios taxonómicos
Rhinodrilus alecrisus sp. nov. es cercana a R. curiosus (Righi et al., 1976), R. curtus (Stephenson, 1931) y R. lourdesae (Righi, 1986) por la posición del clitelo y la presencia de 1 par de glándulas prostáticas. Sin embargo, se diferencian por las siguientes características. Rhinodrilus curiosus contiene poros de las espermatecas en los segmentos 7-8 y 8-9 vs. 1 solo en los segmentos 8-9; tiene 2 pares de espermatecas en 8-9 con un pequeño divertículo vs.1 par en 9 y 10 sin divertículo; difiere en la forma y posición de las estructuras en el campo genital masculino y presenta 1 par de glándulas prostáticas en el segmento 20 vs. 19-20; muestra 2 pares de corazones en 10 y 11 vs. 3 pares de corazones intestinales que se sitúan en los segmentos 10-12; exhibe 2 pares de vesículas seminales largas que ocupan más de 2 segmentos vs. cortas y restringidas a los segmentos 11 y 12. Además, por la forma, tamaño de la cerda común, genital y las espermatecas.
Rhinodrilus curtus muestra 2 pares de espermateca en 8 y 9, vs. 1 par en 9 y 10; tiene 2 pares de corazones intestinales en 10 y 11, vs. 10-12; exhibe 2 pares de vesículas seminales largas que ocupan más de 2 segmentos vs. cortas y restringidas a los segmentos 11 y 12. Además de la forma, tamaño de la cerda común, genital y las espermatecas.
Rhinodrilus lourdesae presenta 2 pares de poros de las espermatecas en 7-8 y 8-9 vs. 1 par en 8-9; la transición esófago-intestino se da en el segmento 15, con inicio del tiflosol en 26 vs. 21-22 y 24-25; la fosa puberal se despliega en el segmento 21 entre cerdas aa, vs. el intercepto de 1920, la molleja el larga, cilíndrica y cónica, vs. redonda y muy musculosa. Además de la forma, tamaño de la cerda común, genital y las espermatecas.
Rhinodrilus buree sp. nov.
http://www.zoobank.org/urn:lsid:zoobank.org:act:7CFCA6D4-A72F-42EB-9B45-F3B69C5E19DD

(fotografía: A. Feijoo).
Figura 2 Rhinodrilus buree sp. nov. Holotipo. CNLT-UTP-0308. A, Cerda común, segmento 35; B, cerda genital, segmento 8; C, vista ventral del clitelo entre los segmentos 16-25; D, espermatecas de los segmentos 8 y 9; E, muestra del par de próstatas y los conductos deferentes de los segmentos 19 y 20.
Descripción. Especie epigea. Caracteres externos. Forma del cuerpo cilíndrica. Color del ejemplar es marrón en el dorso, gris en el clitelo y amarillo claro en la región ventral. La longitud del holotipo es 205.2 mm, con diámetro en la región anteroclitelar de 4.7 mm. Número segmentos 181. Cinco paratipos: longitud y diámetro 171.0 mm, 4.18 mm; 168.4, 4.92; 143.5, 4.3; 122.4, 4.0 y 119.4, 3.96, respectivamente. Número de segmentos: 180, 169, 149, 144, 141, respectivamente. En el holotipo, los 2 primeros segmentos se encuentran invaginados y el prostomio es lobulado y presenta 5 divisiones con forma de anillos. Se presentan 8 hileras de cerdas estrechamente pareadas. Las cerdas ab son visibles a partir del segmento 6. La cerda común mide 0.9 mm y tiene forma de gancho con una fuerte curvatura en el extremo superior y en 2/3 tiene un ensanchamiento que luego se reduce y deja ver algunas líneas incompletas que se alternan en la ornamentación de la cerda (Fig. 2A). La relación en las cerdas entre los segmentos 35-40 es aa:ab:bc:cd:dd = 6.6:1.0:8.3:0.7:26.4. Las cerdas ab de los segmentos 8 y 20-22, se transforman en genitales, la cerda mide 2.9 mm, está recubierta por folículos largos, es casi recta, pues al inicio tiene una ligera curvatura y luego finaliza con forma de punta de aguja, además, en la ornamentación presenta profundas excavaciones con forma “u” (Fig. 2B).
El clitelo en forma de silla ocupa los segmentos 1624, 25 (= 9, 10), mientras que el campo genital masculino se localiza entre 18-22 (= 5), en el segmento 21 se forma una depresión con forma de rectángulo y bordes circulares. Pequeñas papilas circulares redondean las cerdas ab de los segmentos 7 hasta 25 (Fig. 2C). Los poros masculinos se localizan en 19 y 21, mientras que los poros femeninos se visualizaron en el intersegmento 16-17. Los nefridioporos en línea con las cerdas d. Dos pares de poros de las espermatecas en 7-8, 8-9. Dos pares de poros prostáticos se presentan en el intersegmento de 20-21 y 21-22.
Caracteres internos. Los septos de los segmentos 6-7 y 12-13 son delgados y fuertes. La molleja redonda musculosa se sitúa en la cavidad del segmento 6. Tres pares de glándulas calcíferas de estructura tubular dicotómica se localizan en cada uno de los segmentos 7-9. La transición esófago-intestino comienza en los segmentos 22-23 y el tiflosol inicia en el intersegmento 25-26. No hay ciego intestinal. Tres pares de corazones laterales en 7-9 y 4 pares de corazones intestinales se sitúan en los segmentos 10-13. Hay 1 par de holonefridios por segmento. Dos pares de sacos testiculares periesofágicos se localizaron en 10 y 11. En los segmentos 20 y 22 hay 2 pares de glándulas prostáticas con forma alargada y de color blanquecino. Las vesículas seminales pares en los segmentos 11 y 12. Dos pares de conductos deferentes atraviesan cada próstata y a cada lado donde se sitúan las 4 glándulas prostáticas en los segmentos 20-21 y 21-22 (Fig. 2E), para luego desembocar al exterior por el poro masculino en los mismos segmentos. Un par de ovarios se presentan en la cavidad del segmento 13 y el poro femenino se visualizó en 16-17; los embudos femeninos no se visualizaron. Dos pares de espermatecas con forma de lengüeta están en los segmentos 8 y 9 (Fig. 2D).
Resumen taxonómico
Material examinado. Material tipo. Holotipo: espécimen clitelado (CNLT-UTP-0308). Colombia. Departamento de Amazonas: Leticia, comunidad de San Antonio de los Lagos (04°11’35.8” S, 69°57’9.5” O). Tipo vegetación: bosque secundario inundable. Altitud: 95 m snm, colectada en el suelo. Fecha colecta: 14 de septiembre de 2016. Col.: J. Guerra. Paratipos: Colombia. Departamento de Amazonas: 5 adultos (CNLT-UTP-0309), Leticia, comunidad San Antonio de los Lagos (04°11’35.8” S, 69°57’9.5” O), selva secundaria inundable. Altitud: 95 m snm. Fecha colecta: 14 de septiembre de 2016. Col.: J. Guerra.
Etimología. La nueva especie fue nombrada con la palabra buree, que significa lombriz de tierra en lengua Ticuna, en homenaje a la comunidad indígena donde se encontró por primera vez. La población local en español reconoce la especie con el nombre de “zapana”, por su gran tamaño.
Comentarios taxonómicos
Rhinodrilus buree sp. nov., está cercana a R. senex (Righi, 1984a) por la presencia de 2 pares de glándulas prostáticas. Sin embargo, se distancian porque en R. senex, el campo genital masculino tiene otras características, además, difieren en la forma, tamaño de la cerda común, genital y las espermatecas, no tiene vesículas seminales y presenta 5 pares de corazones.
Rhinodrilus muruiborane sp. nov.
http://www.zoobank.org/urn:lsid:zoobank.org:act:B5084416-4DCE-4ADF-9761-6D17BE3BEF4B

(fotografía: A. Feijoo).
Figura 3 Rhinodrilus muruiborane sp. nov. Holotipo. CNLT-UTP-0306. A, Cerda común, segmento 35; B, cerda genital, segmento 8; C, vista ventral del clitelo, enseña la fosa genital; D, espermatecas de los segmentos 7, 8 y 9.
Descripción. Especie epigea. Caracteres externos: Forma del cuerpo cilíndrica con reducción del diámetro después del segmento 35. Color del ejemplar es azul en el dorso, gris en el clitelo y amarillo claro en la región ventral. La longitud del holotipo es 74.6 mm, con diámetro en la región anteroclitelar de 4.18 mm. Número segmentos 63. Un paratipo: longitud 119.4 mm y ancho 4.4 mm. Número de segmentos: 184. En el holotipo se observan 8 hileras de cerdas estrechamente pareadas y visibles a partir del segmento 8. La cerda común mide 0.62 mm y tiene forma de gancho con una fuerte curvatura en el extremo superior (Fig. 3A). La relación entre las cerdas entre los segmentos 40 y 45 es aa:ab:bc:cd:dd = 6.5:1.0:9.0:7.5:27.5. Las cerdas ab de los segmentos 7-9 y 18-23 se transforman en genitales, ella mide 1.1 mm, está recubierta por folículos, presenta ligera curvatura al inicio, después es recta y termina con punta de lanza, tiene algunas excavaciones de forma ovalada que se alternan de manera desigual en los contornos (Fig. 3B).
El clitelo ocupa los segmentos ½ 16-25 (= 9.5) y los tubérculos de la pubertad se localizan entre ½19 - ½23 (= 4) (Fig. 3C). Los poros masculinos se localizan en el intersegmento 16-17, mientras que los poros femeninos se visualizaron en 14-15. Los nefridioporos en línea con las cerdas c. Tres pares de poros de las espermatecas en 6-7, 7-8, 8-9.
Caracteres internos. Los septos de los segmentos 6-7 y 12-13 son delgados con forma de membranas. La molleja con forma de disco se encuentra en la cavidad del segmento 6. Tres pares de glándulas calcíferas de estructura tubular dicotómica se localizan en cada uno de los segmentos 7-9. La transición esófago-intestino se da en el segmento 23 y el tiflosol comienza en el segmento 24. No hay ciego intestinal. Se localizaron 3 pares de corazones laterales en 7-9 y 2 pares de corazones intestinales en los segmentos 10 y 11. Hay 1 par de holonefrídios por segmento. Dos pares de sacos testiculares periesofágicos se ubicaron en 10 y 11. Dos pares de vesículas seminales se sitúan en los segmentos 11 y 12. Los infundíbulos seminales partes desde los segmentos 10 y 11 para luego desembocar en el intersegmento 16/17. Un par de ovarios se presentan en la cavidad del segmento 13 y el poro femenino se visualizó en 14-15; los embudos femeninos no se visualizaron. Tres pares de espermatecas con forma de raqueta, están en los segmentos 7-9 (Fig. 3D), que se encuentran próximas a 3 pares de masas glandulares. La especie no presenta próstatas.
Resumen taxonómico
Material examinado. Material tipo. Holotipo: espécimen clitelado (CNLT-UTP-0306). Colombia. Departamento del Amazonas: corregimiento de La Chorrera, comunidad de Lago Grande (2°04’14.4” S, 72°10’14.2” O). Tipo vegetación: planicies no inundables con bosque secundario maduro donde predominan las palmas, sin uso reciente. Altitud: 100 m snm, colectada en la hojarasca. Fecha colecta: 28 de junio de 2016. Cols.: D. Castro, O. Ramírez y C. Peña-Venegas. Paratipos: 1 adulto, igual información que para el holotipo. CNLT-UTP-0307.
Etimología. El nombre específico fue propuesto como homenaje a la comunidad de Lago Grande la cual está compuesta por una población Huitoto (murui = dialecto) en territorio Bora, que, al unir los 2 nombres, conjuga palabras que identifican étnicamente a la comunidad.
Comentarios taxonómicos
Rhinodrilus muruiborane sp. nov., está estrechamente relacionada con R. appuni appuni (Michaelsen, 1892), R. romani (Michaelsen, 1928) y R. senex (Righi, 1984a), por la ubicación del clitelo y se diferencia de éstas por la posición de los tubérculos de la pubertad, el número, forma y tamaño de las espermatecas (3 pares vs. 2), posición de los poros masculinos, la transición esófago intestino y el inicio del tiflosol.
Rhinodrilus sinchi sp. nov.
http://www.zoobank.org/urn:lsid:zoobank.org:act:ABE4BC05-9623-4636-B462-AE39CCD05F61

(fotografía: A. Feijoo).
Figura 4 Rhinodrilus sinchi sp. nov. Holotipo. CNLT-UTP-0314. A, Cerda común, segmento 35; B, cerda genital, segmento 19; C, vista ventral del clitelo entre los segmentos 16-24; D, vista interna de la molleja. M: Molleja, E: espermateca, GC: glándulas calcíferas, P: próstata.
Descripción. Especie epigea. Caracteres externos: forma del cuerpo cilíndrica. Color del ejemplar es marrón en la región dorsal, amarillo pálido en la región ventral y blanco lechoso en el clitelo. La longitud del holotipo es 50.4 mm, con diámetro en la región anteroclitelar de 2.28 mm. Número de segmentos 121. Longitud del paratipo 25.2 mm, con diámetro en la región anteroclitelar de 1.65 mm; número de segmentos 86. En el holotipo los segmentos son visibles a partir del segundo y el prostomio es pequeño y de forma lobular segmentado. Ocho hileras de cerdas estrechamente pareadas. Las cerdas ab son visibles a partir del segmento 4. La cerda común mide 0.25 mm, es recta y con mayor grosor en el centro (Fig. 5a). La relación entre las cerdas entre los segmentos 35-40 es aa:ab:bc:cd:dd = 5.0:1.0:6.0:1.0:16.0. Las cerdas ab de los segmentos 19- 21 se transforman en genitales, la cerda genital mide 1.1 mm, está recubierta por folículos largos, es semirrecta y a partir de 2/3 de su longitud comienza la ornamentación con algunas excavaciones en forma cónica y finaliza en punta (Fig. 5b).
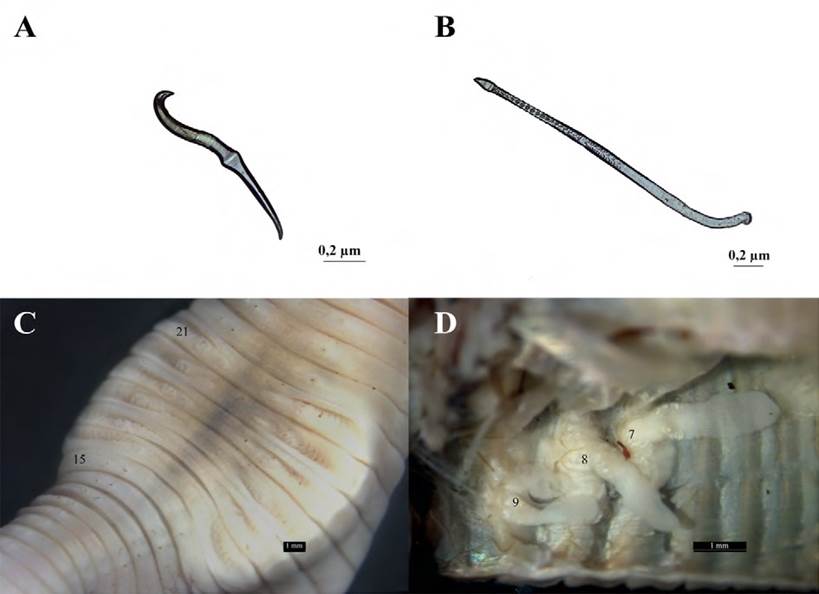
(fotografía: A. Feijoo).
Figura 5 Diachaeta (Amazo) sabalomurui sp. nov. Holotipo. CNLT-UTP-0312. A, Cerda común, segmento 35; B, cerda genital, segmento 19; C, vista ventral del clitelo; D, espermatecas de los segmentos 7, 8 y 9.
El clitelo en forma de silla ocupa los segmentos 1624 (= 9), y en el campo genital masculino se localiza una fosa puberal en el intercepto del intersegmento 19/20 en medio de las cerdas ab (Fig. 5c). Los poros masculinos microscópicos se localizan en el intersegmento de 1920, mientras que los poros femeninos microscópicos se visualizaron en 16-17; los embudos femeninos no se visualizaron. Los nefridioporos en el centro de las cerdas cd. Un par de poros espermatecales se localizan en 6-7.
Caracteres internos. Los septos de los segmentos 6-7 y 13-14 tienen forma de finas membranas, cónicos e interpenetrados. La molleja es redonda y musculosa y se sitúa en la cavidad del segmento 6. Tres pares de glándulas calcíferas en los segmentos 7-9 con estructura trabecular. La transición esófago-intestino comienza en el intersegmento 26-27 y el tiflosol inicia en el intersegmento 27-28. No hay ciego intestinal. Tres pares de corazones laterales en 7-9 y 2 pares de corazones intestinales se sitúan en los segmentos 10, 11. Hay 1 par de holonefrídios por segmento. Dos pares de test í culos debajo de los corazones de 10 y 11; los conductos deferentes van de cada lado y se desplazan hasta la próstata e ingresan en ella, para luego desembocar en el poro masculino. La especie presenta 1 par de próstatas con forma de racimo de uvas (racimosa) con un conducto visible, de color blanco en el segmento 19-20. En el segmento 11 no hay vesículas seminales, mientras que las de 12 están presentes. En el segmento 13 se localizaron los ovarios; los embudos femeninos no se visualizaron. Un par de espermatecas con forma de lengüeta ocupan 4 segmentos y se sitúan en el segmento 7 (Fig. 5d).
Resumen taxonómico
Material examinado. Material tipo. Holotipo: espécimen clitelado (CNLT-UTP-0314). Colombia. Departamento del Amazonas: corregimiento de Puerto Arica, comunidad Sábalo (02°21’11.7” S, 72°35’53.4” O). Tipo vegetación: bosque secundario moderadamente intervenido. Altitud: 95 m snm, colectada en el suelo. Fecha colecta: 3 de julio de 2016. Colectores: D. Castro, O. Ramírez y C. Peña-Venegas. Paratipo: 1 subadulto, igual información que para el holotipo. CNLT-UTP-0315.
Etimología. La nueva especie fue nombrada con la palabra sinchi que viene del vocablo indígena “sinchi” que significa conocedor, en homenaje a todos los conocedores indígenas e investigadores de la Amazonia colombiana.
Comentarios taxonómicos
Rhinodrilus sinchi sp. nov., es una especie cercana a R. curiosus (Righi et al., 1976), R. curtus (Stephenson, 1931) y R. pashanasii (Righi, 1992), por la posición del clitelo en lugares próximos a los segmentos 16-24 y la presencia de fosa puberal cercano al intersegmento 19/20. R. sinchi se distancia de las otras especies por las siguientes características:
Rhinodrilus curiosus presenta prostomio tentaculiforme (R. sinchi lobular), 2 pares de espermatecas en 8 y 9 (R sinchi, 1 par en 7), 2 pares de vesículas seminales en 11 y 12 (R sinchi, 1 par en 12), 1 par de próstatas sésiles y voluminosas en segmento 20 (R sinchi, 1 par racimosas pequeñas en 19/20). R. curtus, 2 pares de espermatecas sin divertículo en 7 y 8 (vs. 1 par en 7), 2 pares de vesículas seminales en 11 y 12 (vs. 1 par en 12) y 1 par de próstatas en segmento 21 (vs. 1 par en 19/20). R. pashanasii, 2 pares de espermatecas en 8 y 9 (vs. 1 par en 7), tubérculos de la pubertad en 19-21 (vs. fosa puberal en el intercepto del intersegmento 19-20), 4 pares de corazones intestinales entre los segmentos 10-13 (vs. 2 pares en 10, 11), 2 pares de vesículas seminales en 11 y 12 (vs. 1 par en 12) y ausencia de próstatas (vs. 1 par en 19-20).
Género: Diachaeta (Benham, 1887)
Diagnosis: 8 hileras de cerdas por segmento dispuestas en series longitudinales regulares, raramente irregulares. Un par de poros masculinos intraclitelares. Una molleja en el segmento 6. Tres pares de glándulas calcíferas en los segmentos 7-9 con estructura ventral trabecular. Un par de testículos e infundíbulos seminales en el segmento 10. En un caso se presentan glándulas prostáticas. Un par de ovarios en el segmento 13. De 1 a 3 pares de espermatecas presentes entre los segmentos 6-7 y 8-9.
Especie tipo: Diachaeta thomasi (Benham, 1887)
Subgénero Amazo (Righi, 1984a)
Diachaeta (Opisthocordis) (Righi, 1984b) Siete pares de corazones entre los segmentos 7-13. Las espermatecas presentes o ausentes.
Especie tipo: Diachaeta nia (Righi et al., 1976)
Número de especies del género 21. Distribución. Brasil (11), Colombia (3), Venezuela (2), Jamaica (2), Barbados (1), Bonaire (1) y Argentina (1) (Brown y Fragoso, 2007).
Diachaeta (Amazo) sabalomurui sp. nov.
http://www.zoobank.org/urn:lsid:zoobank.org:act:641FA819- D48F-4A7E-8F45-34B47C8CE037
Descripción. Especie epigea. Caracteres externos: forma del cuerpo cilíndrica. Color del ejemplar es amarillo pálido en el dorso, blanquecino en el clitelo y gris en la región ventral. La longitud del holotipo es 59.9 mm, con diámetro en la región anteroclitelar de 2.5 mm. Número segmentos 151. Dos paratipos: longitud 42.4 mm y ancho 2.0 mm; 32.9, 1.6, respectivamente. Número de segmentos: 157 y 159, respectivamente. En el holotipo los segmentos son visibles a partir del segundo y el prostomio es pequeño y de forma triangular. Ocho hileras de cerdas estrechamente pareadas. Las cerdas ab son visibles a partir del segmento 4. La cerda común mide 0.35 mm y tiene forma de gancho en el ápice, con una fuerte curvatura en el extremo (Fig. 5A). La relación entre las cerdas entre los segmentos 35-40 es aa:ab:bc:cd:dd = 2.6:1.0:4.3:0.7:9.9. Las cerdas ab de los segmentos 19 - 20 se transforman en genitales, la cerda genital mide 0.55 mm, está recubierta por folículos largos, es curva en la región proximal y recta en el ápice y finaliza en punta con forma de lanza y presenta algunas excavaciones con formas semilunares (Fig. 5B).
El clitelo en forma de silla ocupa los segmentos 15, 1625 (= 10, 11), los tubérculos de la pubertad se encuentran entre los segmentos 19-24 (Fig. 5C). Los poros masculinos se localizan en el intersegmento 17/18, mientras que los poros femeninos se visualizaron en 16. Los nefridioporos en línea con las cerdas cd. Tres pares de poros de las espermas se localizan en 6-7, 7-8 y 8-9.
Caracteres internos. Los septos de los segmentos 6-7 y 14-15 son medianamente gruesos, cónicos e interpenetrados. La molleja inicia con forma de punta de zapato y luego es ovalada y se sitúa en la cavidad del segmento 6. Tres pares de glándulas calcíferas en los segmentos 7-9 con estructura ventral trabecular. La transición esófago-intestino comienza en el intersegmento 17-18 y el tiflosol inicia en el segmento 18-19. No hay ciego intestinal. Tres pares de corazones laterales en 7-9 y 4 pares de corazones intestinales se sitúan en los segmentos 10-13. Hay 1 par de holonefrídios por segmento. Un par de testículos e infundíbulos seminales en el segmento 11. Un par de vesículas seminales con forma de lóbulos, se encuentran restringidas al segmento 12. Un par de ovarios en el segmento 13; los embudos femeninos no se visualizaron. Entre los segmentos 17-21 se identifican 5 pares de masas glandulares. Tres pares de espermatecas con forma de lengüeta en los segmentos 7-9 (Fig. 5D).
Resumen taxonómico
Material examinado. Material tipo. Holotipo: espécimen clitelado (CNLT-UTP-0312). Colombia. Departamento del Amazonas: Corregimiento de Puerto Arica, comunidad Sábalo (02°21’11.7” S, 72°35’53.4” O). Tipo vegetación: bosque secundario moderadamente intervenido. Altitud: 95 m snm, colectada en el suelo en estrato 0-10 cm. Fecha colecta: 3 de julio de 2016; colls.: D. Castro, O. Ramírez y C. Peña-Venegas. Paratipos: igual información que para el holotipo. CNLT-UTP-0313.
Etimología. La nueva especie fue nombrada con la palabra “sabalomurui” en homenaje a la comunidad indígena donde se encontró por primera vez (Sábalo) y a la etnia que mora en la localidad (murui).
Comentarios taxonómicos
El subgénero Diachaeta (Amazo) contiene 12 especies, de las cuales 6 presentan clitelo anular: D. (A.) aceoca (Righi,1982), D. (A.) arawak (Righi, 1986), D. (A.) exul (Stephenson,1931), D. (A.) juli (Righi et al., 1978), D. (A.) mingua (Righi y Németh, 1983) y D. (A.) uaimiri (Righi y Németh,1983); 4 lo tienen en forma de silla (D. (A.) carsevenica (Černosvitov, 1934), D. (A.) kannerae ( Righi, 1984b), D. (A.) nia ( Righi et al., 1976), Diachaeta (Amazo) sabalomurui sp. nov.) y 2 combinan la presencia de clitelo anular y forma de silla (D. (A.) atroaris ( Righi et al., 1978), D. (A.) xecatu (Righi et al., 1978). Las especies que conservan clitelo anular y que combinan lo anular con silla se distancian de D. (A.) sabalomurui porque solo presenta clitelo en forma de silla, mientras que las otras especies son próximas por la forma del clitelo, sin embargo, se diferencian por las siguientes características. En Diachaeta (A.) carsevenica el clitelo se ubica en los segmentos 13-21 (vs. 15, 16-25), los tubérculos de la pubertad son estrechos y se localizan en 1/214-1/2 21 (vs. 19-24), carece de espermatecas (vs. 3 pares en 7, 8, 9) y presenta bolsas espermáticas en el intersegmento 13/14. En D. (Am.) kannerae el clitelo se sitúa en 14-1/221 (vs. 15, 16-25), par de papilas crateriformes y alargadas en 17, al lado de b, que contiene los poros masculinos (vs. papilas ausentes), ausencia de espermatecas (vs. 3 pares en 7, 8, 9) y también por la forma, tamaño de la cerda genital y las espermatecas. En D. (A.) nia el clitelo se presenta en 14-20 (vs. 15, 16-25), tubérculos de la pubertad 1 par voluminoso y ubicado en la media ventral en bc en 15-16, 2 pares de espermatecas en 13-14 (vs. 3 pares en 7, 8, 9) y también por la forma, tamaño de la cerda genital y las espermatecas.
Discusión
Las especies de Rhinodrilus presentan fuertes variaciones en el tamaño, desde individuos muy pequeños que miden desde 19 mm (R. proboscideus) o 92 a 82 mm (R. adelae) hasta los de mayor tamaño, que fluctúan entre los 565-625 mm (R. alatus Righi, 1971) o entre 620 a 860 mm (R. priolli Righi, 1967). El clitelo por lo general se visualiza entre los segmentos 15-27 y solo en R. alatus se observa entre 1/216, 17-30, 31. Los poros masculinos generalmente son microscópicos, por lo tanto, la localización, se debe realizar a partir de la disección de la lombriz y hacer el seguimiento e inspección de los conductos deferentes, para conocer el lugar de la desembocadura del poro hacia el exterior. Los poros se localizan en el intersegmento 18/19 (R. alatus), 19/20 (R. panxin Righi, 1971), 20/21 (R. ayacu Righi, 1996), 22/23 (R. pitun Righi y Ferrer, 1990), 23/24 (R. bursiferus Righi, 1971), o en el segmento 20 (R. priolli Righi, 1967) o 21 (R. mamita Cordero, 1943).
Las espermatecas en Rhinodrilus difieren en el número de pares y la posición, los que pueden ser 1 par en 7 (R. sinchi), 2 pares entre los segmentos 8-9 (R. appuni appuni Michaelsen, 1892), 3 pares en 6-8 (R. fransisci Cordero, 1944) o 7-9 (R. adelae Cordero, 1943), mientras que en 3 especies se han encontrado 5 pares en 7-11 (R. alatus), 7 pares en 7-13 (R. parvus Rosa, 1895) y 9 pares en 7-15 (R. fafner Michaelsen, 1918). Las vesículas seminales en solo un caso están ausentes (R. timote Righi, 1989) y en la mayoría de las especies son cortas y restringidas a los segmentos 11 y 12, excepto en 3 especies: en la primera, 1 par se ubica sobre la región dorsal del esófago en el segmento 11, mientras que el segundo par, se localiza en los segmentos 13-14 (R. pitun); en la segunda y tercera especies, son largas y van más allá del segmento 20 (R. curiosus Righi et al., 1976 y R. elisianeae) (Righi et al., 1976).
En las 48 especies descritas en Rhinodrilus se destaca el carácter holoándrico del género, por la presencia de 2 pares de testículos, 1 en el segmento 10 y otro en 11, provisto de 3 pares de glándulas calcíferas y con tubos en panoja en la estructura de la glándula calcífera. Además, Rhinodrilus contiene testículos encerradas en vesículas, por lo tanto, evolucionaron en vías diferentes de géneros como Diachaeta, pues la gimnorquidia se supone fue anterior a la cleistorquidia (Feijoo, 1993).
En Diachaeta (Amazo), el tamaño de las especies es pequeño y fluctúa entre 20 a 28 mm (D. (A.) aceoca Righi, 1982), hasta 80 mm (D. (A.) arawak Righi, 1986). La posición de los poros masculinos en las especies en el subgénero es muy variable y se pueden localizar en el segmento 15 (D. (A.) atroaris Righi et al., 1978), 16 (D. (A.) nia Righi et al., 1976), 17 (D. (A.) kannerae Righi, 1984b), 18 (D. (A.) exul Stephenson, 1931), o en los intersegmentos 15-16 (D. (A.) aceoca Righi, 1982), 17-18 (D. (A.) sabalomurui, este estudio), 18-19 (D. (A.) arawak Righi, 1986), 25/26 (Diachaeta (Amazo) uaimiri Righi y Németh, 1983).
En Diachaeta (Amazo), el número y la posición de las espermatecas es inconstante y fluctúa desde 1 par en el segmento 10 (D. (A.) arawak), 2 pares en 9, 10 (D. (A.) mingua Righi y Németh, 1983) o, 13, 14 (D. (Am.) aceoca) hasta 3 pares en 7-9 (D. (A.) sabalomurui, este estudio). Las vesículas seminales pueden ser cortas y restringidas al segmento 12 (D. (A.) atroaris) o largas con punto de partida en el segmento 12 y proyectadas hacia atrás hasta el segmento 48.
La presencia de glándulas prostáticas en las especies R. alecrisus sp. nov. (1 par en los segmentos 19-20), R. buree sp. nov. (2 pares en 19-20) y R. sinchi sp. nov. (1 par de glándulas prostáticas en 19-20) son estructuras poco identificadas en las especies de Rhinodrilidae, pero que ya se habían descrito por Righi (1985) con presencia de 1 par en R. curiosus (en segmento 19/20) (Righi et al., 1976), R. curtus (21) (Stephenson, 1931), R. lourdesae (21) (Righi, 1986).
Las próstatas son órganos reproductores asociados a las genitalia masculina que consisten en una glándula y un conducto generalmente muscular para expusar la secreción. En la familia Megascolecidae están presentes en todas sus especies, con 3 tipos principales: tubular, túbulo racimosa y racimosa (forma de racimo de uvas) (Jamieson, 2006). Poseen la función de proporcionar exudados que ayudan en la entrega de esperma a la pareja durante la copulación, y probablemente cargan el ámpula de la espermateca de la pareja con soluciones nutritivas (Blackmore, 2006).
La presencia de glándulas prostáticas verdaderas (metapróstatas), es decir, glándulas con un conducto situadas atrás y asociados con el sistema reproductor masculino, es una característica exclusiva de Megascolecidae y Ocnerodrilidae. Se distinguen de las llamadas eupróstatas, de Eudrilidae, que en realidad son una dilatación del conducto masculino (Jamieson, 2006); que en esta familia se distingue por la presencia de espermatecas postesticulares (Rodríguez, 1987); característica también presente en algunas especies de Rhinodrilidae, como en D. (A.) aceoca. Asimismo, en Glossoscolecidae existen estructuras parecidas que se han descrito como cámaras copulatorias con formas tubulares y con conductos muy cortos y próximos a la región parietal (Feijoo y Brown, 2018).
En las glándulas descritas para R. alecrisus sp. nov. y R. sinchi sp. nov., se observaron las formas tubulares o racimosas con presencia de conducto, mientras que en R. buree sp. nov. no se visualizaron estas formas, sino que ellas están presentes con forma de lengua. En las 2 primeras, el conducto es perceptible, y con la disección, se logró observar que el conducto central interno recorre la glándula y se une directamente en la región intraparietal, para después desembocar al exterior en el campo genital. Posiblemente, lo visto para R. buree sp. nov. corresponde más con eupróstatas; sin embargo, se requieren de nuevos hallazgos e investigación para conocer las variaciones y el papel funcional de éstas estructuras. Asimismo, los conductos deferentes no parecen ingresar a las glándulas en R. buree sp. nov., ya que toman camino independiente de la próstata. Al respecto, Gates (1972) considera que la presencia de eupróstatas no conectadas con ningún conducto de esperma demuestra que las glándulas no pueden ser solo engrosamientos del conducto deferente, sino que ellas se desarrollan a partir invaginaciones parietales, hecho observado en Rosadrilus (Cognetti-De Martiis, 1906).

