











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Cryptocanthon Balthasar es un género neotropical de escarabajos perteneciente a la subfamilia Scarabaeinae. Son principalmente coprófagos de pequeño tamaño, entre 3-5.2 mm de largo y 1.7-3.2 mm de ancho, distribuidos en ambientes húmedos con abundante hojarasca en ecosistemas de bosques lluviosos, montanos y nublados desde México hasta Brasil (Cook, 2002). Aspectos básicos sobre su biología como ciclos de vida, estrategias de nidificación, actividad diaria, requerimientos alimenticios y microambientales, además de descripciones de sus estados inmaduros son desconocidos. Generalmente en estudios ecológicos, el género ha sido asignado al gremio rodador, sin embargo, no existe evidencia empírica que confirme dicho comportamiento, incluso su comportamiento rodador es dudoso debido a sus limitantes físicas de tamaño corporal, que es semejante a otros géneros neotropicales como Canthochilum Chapin, Agamopus Bates y Zonocopris Arrow, los cuales presentan tamaños menores a 7 mm y no son capaces de generar y rodar bolas de excremento o utilizan otros recursos para alimentación (Halffter y Halffter, 1989; Vaz de Mello, 2007).
La posición sistemática del género es incierta y su monofila no ha sido comprobada. Tradicionalmente Cryptocanthon ha sido ubicado en la tribu Deltochilini (= Canthonini), sin embargo, en la reciente definición de la tribu Deltochilini sensu novo el género no fue incluido (Tarasov y Dimitrov, 2016), siendo más adecuada su ubicación en la tribu Demarziellini (Vaz de Mello, 2007). Balthasar (1942) describió el género con base en la especie C. paradoxus de Ecuador y mencionó su probable relación con los géneros Ipselissus Olsoufieff (= Canthonella Chapin) y Zonocopris Arrow. Howden (1973) revisó por primera vez el género, describió 8 especies y sugirió un mayor grado de parentesco con Bdelyropsis Pereira, Vulcano y Martínez. Tres años después, Howden (1976) describió 2 especies de Colombia y México y en la década siguiente, Howden y Gill (1987) adicionaron 2 especies más de Panamá y Costa Rica. Vaz de Mello (2007) incluyó a Cryptocanthon en el análisis filogenético del género Zoonocopris Arrow, sugiriendo una relación de parentesco entre Zonocopris y el clado Cryptocanthon + (Paracryptocanthon + Bdelyrus). Tarasov y Génier (2015) establecieron como grupo monofilético al clado conformado por los géneros Apotolamprus y Arachnodes de Madagascar y su parentesco con el género neotropical Bdelyrus, que combinados (Bdelyrus + (Apotolamprus + Arachnodes)) constituyen actualmente el grupo hermano de Cryptocanthon.
El estudio sistemático más reciente del género fue realizado por Cook (2002), en el cual se establecieron relaciones de parentesco a nivel de grupos: 1) especies relictuales del norte de América del Sur (C. andersoni Cook, C. gilli Cook y C. nebulinus Howden); 2) especies de tierras bajas amazónico-guyanesas (C. campbellorum Howden, C. galboa Cook, C. peckorum Howden y C. pumilus Cook); 3) especies de América Central (C. bochilae Cook, C. brevisetus Howden, C. cristobalensis Howden, C. denticulum Cook, C. howdeni Cook, C. lindemanae Howden y Gill, C. lobatus Howden, C. montebello Cook, C. newtoni Howden, C. ocosingo Cook, C. osaensis Cook, C. rayonensis Cook y C. solisi Cook), y 4) especies andinas (C. altus Howden, C. chiriquinus Howden y Gill, C. curticrinus Cook, C. escobari Cook, C. foveatus Cook, C. Génieri Cook, C. humidus Howden, C. lobipygus Cook, C. medinae Cook, C. napoensis Cook, C. otonaga Cook, C. paradoxus Balthasar, C. parvus Howden, C. punctatus Cook y C. urguensis Cook). En total, el estudio incluyó 35 especies de las cuales 22 fueron nuevas. Posteriormente, 2 trabajos adicionaron 4 especies al género: C. andradei, C. buriticaorum y C. pulidoae de Colombia (Arias-Buriticá y Medina, 2014), y C. chimalapensis de México (Mora-Aguilar y Delgado, 2018). Así, el género actualmente comprende 39 especies.
La definición del género Cryptocanthon ha sido sustentada por caracteres homoplásicos en diferentes propuestas filogenéticas (Cook, 2002; Tarasov y Génier, 2015; Vaz de Mello, 2007). La siguiente combinación de caracteres es diagnóstica: cuerpo ovalado y convexo dorsalmente (Figs. 1,2,3); clípeo cóncavo, margen anterior con 2 a 6 dientes agudos y flexionados, separados por una emarginación en forma de U y superficie vertical del clípeo con o sin fovea media (Fig. 9); ojos en vista dorsal ligeramente visibles o no, cuando la cabeza se retrae (Figs. 1,2,3); hipómero pronotal excavado anteriormente, a menudo con carena longitudinal incompleta en la mitad basal (Fig. 8F); élitros con 7 estrías dorsales, cada estría conformada por 2 líneas onduladas paralelas (Fig. 8C, D); élitros bruscamente inflexos lateralmente en la séptima estría, formando una amplia pseudoepipleura (Figs. 5F, 6F, 8G, H); estrías 8 a 10 ubicadas en la pseudoepipleura; octava y novena estría a menudo incompletas o ausentes, décima estría indicada por una hilera de puntuaciones cerca del margen epipleural; a menudo con pequeña fovea basal cerca del margen basal; epipleura surge ligeramente por encima del nivel de la pseudoepipleura y es más estrecha (Fig. 8 G, H); pata anterior con foseta trocanto-femoral anterior situada en el borde del trocánter; espolón apical de la protibia simple; pigidio vertical, convexo, con tumefacción medial o cresta longitudinal, base carenada, con surco medio (Cook, 2002; Vaz de Mello et al., 2011).

Figura 1 Aspecto dorsal de las especies del género Cryptocanthon de Colombia. A) C. altus, B) C. andradei, C) C. buriticaorum, C. campbellorum, E) C. escobari, F) C. foveatus. Escala: 1mm.

Figura 2 Aspecto dorsal de las especies del género Cryptocanthon de Colombia. A) C. humidus, B) C. medinae, C) C. parvus, D) C. peckorum, E) C. pulidoae, F) C. pumilus. Escala: 1 mm.

Figura 3 Nuevas especies. A) C. cephalopunctatus sp. nov. ♂, B) C. cephalopunctatus sp. nov. ♀, C) C. encenillensis sp. nov. ♂, D) C. encenillensis sp. nov. ♀, E) C. mailinae sp. nov. ♂, F) C. mailinae sp. nov. ♀. Escala: 1 mm.

Figura 4 Cryptocanthon cephalopunctatus sp. nov. A) Cabeza, B) hipómero, C) tórax (ventral), D) pigidio, E) metatibia, F) inflexión lateral del élitro. Escala: 0.5 mm.

Figura 5 Cryptocanthon encenillensis sp. nov. A) Cabeza, B) hipómero, C) tórax (ventral), D) pigidio, E) metatibia, F) inflexión lateral del élitro. Escala: 0.5 mm.

Figura 6 Cryptocanthon mailinae sp. nov. A) Cabeza, B) hipómero, C) tórax (ventral), D) pigidio, E) metatibia, F) inflexión lateral del élitro. Escala: 0.5 mm.
En Colombia, el género estaba representado por 12 especies pertenecientes a 2 grupos definidos por Cook (2002): 3 especies del grupo de tierras bajas amazónico-guyanesas y 9 del grupo andino, de las cuales 7 son braquípteras. Este último grupo reúne a las especies que no presentan puntuaciones umbilicadas en la cabeza y se considera evolutivamente el más reciente, exhibiendo más caracteres derivados (Cook, 2002). Su evolución habría ocurrido en el Plioceno, durante la orogenia del norte de los Andes y con la subsecuente invasión de especies de tierras bajas transandinas hacia la región de América Central después del cierre del istmo de Panamá, favorecida por el cambio de la corriente subacuática del Atlántico que permitió las condiciones para un clima húmedo hacia el norte a lo largo de la costa atlántica de América Central, lo que habría proporcionado un corredor para la dispersión hacia el norte de la fauna andina adaptada al bosque húmedo (Cook, 2002).
En el presente trabajo se describen 3 especies nuevas de Cryptocanthon de los Andes de Colombia, se proporcionan datos de distribución geográfica, notas sobre el estado de conservación de las especies y una clave de identificación para las especies presentes en el país con base en caracteres morfológicos externos. La revisión de especímenes de Cryptocanthon de la Colección Entomológica del Instituto Humboldt (IAvH-E) permitió la verificación taxonómica de los registros, contribuyendo a la actualización de la lista de especies del país. Con este trabajo, Colombia se posiciona como el país con mayor número de especies con 15 especies, seguido por México con 10.
Materiales y métodos
El estudio se realizó con base en especímenes depositados en la Colección Entomológica del Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH-E). La información adicional sobre la distribución geográfica se obtuvo a partir de la revisión o consulta de registros de las siguientes colecciones (curadores o encargados entre paréntesis):
CECC-CALT: Colección de escarabajos coprófagos de Colombia, Bogotá, Cundinamarca (Alejandro Lopera Toro). CEUA: Colección Entomológica Universidad de Antioquia, Medellín, Colombia (Martha Wolff). CEUN-PSO: Colección Entomológica de la Universidad de Nariño, Pasto, Nariño. (Guillermo Castillo). MEFLG: Museo Francisco Luis Gallego, Universidad Nacional de Colombia, campus Medellín, Antioquia (Sandra Uribe Soto). MEUV: Museo de Entomología de la Universidad del Valle (Carmen Elisa Posso Gómez). UPTC: Museo de Historia Natural Luis Gonzalo Andrade, de la Universidad Pedagógica y Tecnológica de Colombia (Fredy Molano).
La terminología para los caracteres morfológicos internos y externos usados se basan en Howden (1973, 1976) y Cook (2002). La extracción de los órganos genitales se realizó siguiendo la metodología propuesta por Medina et al. (2003). Las estructuras esclerotizadas se aclararon en una solución diluida de hidróxido de potasio (10%) y se neutralizaron en una solución diluida de ácido acético (15%). Los órganos genitales se preservaron en un vial con glicerina el cual se ubicó debajo del espécimen. Los escleritos del saco interno de los parámeros se nombraron siguiendo la terminología de Medina et al. (2013) y Arias-Buriticá y Medina (2014). La descripción del esclerito placa y elongado se basó en la terminología de Gonzalez y Vaz de Mello (2014). Con el fin de observar estructuras taxonómicas importantes (foveas, suturas, surcos y microescultura), los especímenes se lavaron con un limpiador ultrasónico UD1005H-3L siguiendo las recomendaciones de Howden (1973) y fueron examinados usando un estereomicroscopio Nikon SMZ 645 a una magnificación de 6.5-40X. Las medidas de largo total y ancho humeral se realizaron con un micrómetro ocular. Las imágenes del hábito dorsal de las especies y caracteres morfológicos usados en las descripciones fueron tomadas con una cámara MC 190 HD marca Leica de 10 megapixeles adaptada a un estereomicroscopio Trinocular Leica-S8 APO, apilando una serie de fotografías con el programa Helicon focus (Helicon Soft). Los datos espaciales para elaborar los mapas de distribución fueron obtenidos en los lugares de recolección y la información consignada en las etiquetas de los especímenes se convirtió en capas vectoriales de puntos con el programa ArcGIS 10.2 (ESRI 2011, Licencia E300 04/26/2013).
En este trabajo se usó el concepto filogenético de especie de Wheeler y Platnick (2000): “Las especies son conjuntos de pequeñas agregaciones de poblaciones diagnosticables por una única combinación de estados de caracteres. Así, los estados de caracteres son usados para respaldar la hipótesis de que todos los individuos examinados corresponden a la misma especie”. Estos estados únicos de caracteres obedecen principalmente a un aislamiento genético, en el caso de aquellos organismos cuya reproducción es sexual, como el caso de las especies de Cryptocanthon.
Descripción
Cryptocanthon cephalopunctatus Martínez-Revelo, Torres y Neita sp. nov. (Figs. 3A, B, 4, 7O, 9I, 10L, 11O, 12O, 13N, 14N, 15N, 17) http://zoobank.org/urn:lsid:zoobank.org:act:BA6D4122-68E5-4235-AF51-04518E6B43E7

Figura 7 Tibia anterior. A) C. campbellorum, B) C. foveatus, C) C. pulidoae, tibia posterior, D) C. andradei, E) C. buriticaorum, F) C. campbellorum, G) C. escobari, H) C. foveatus, I) C. medinae, J) C. parvus, K) C. pulidoae, sutura meso-metaesternal, L) C. buriticaorum, M) C. pulidoae, N) C. pumilus, O) C. cephalopunctatus. Escala: 0.5 mm.

Figura 8 Pronoto. A) C. mailinae, B) C. pulidoae, élitro, C) C. altus, D) C. foveatus, hipómero, E) C. foveatus, F) C. medinae, inflexión lateral del élitro, G) C. campbellorum, H) C. parvus.
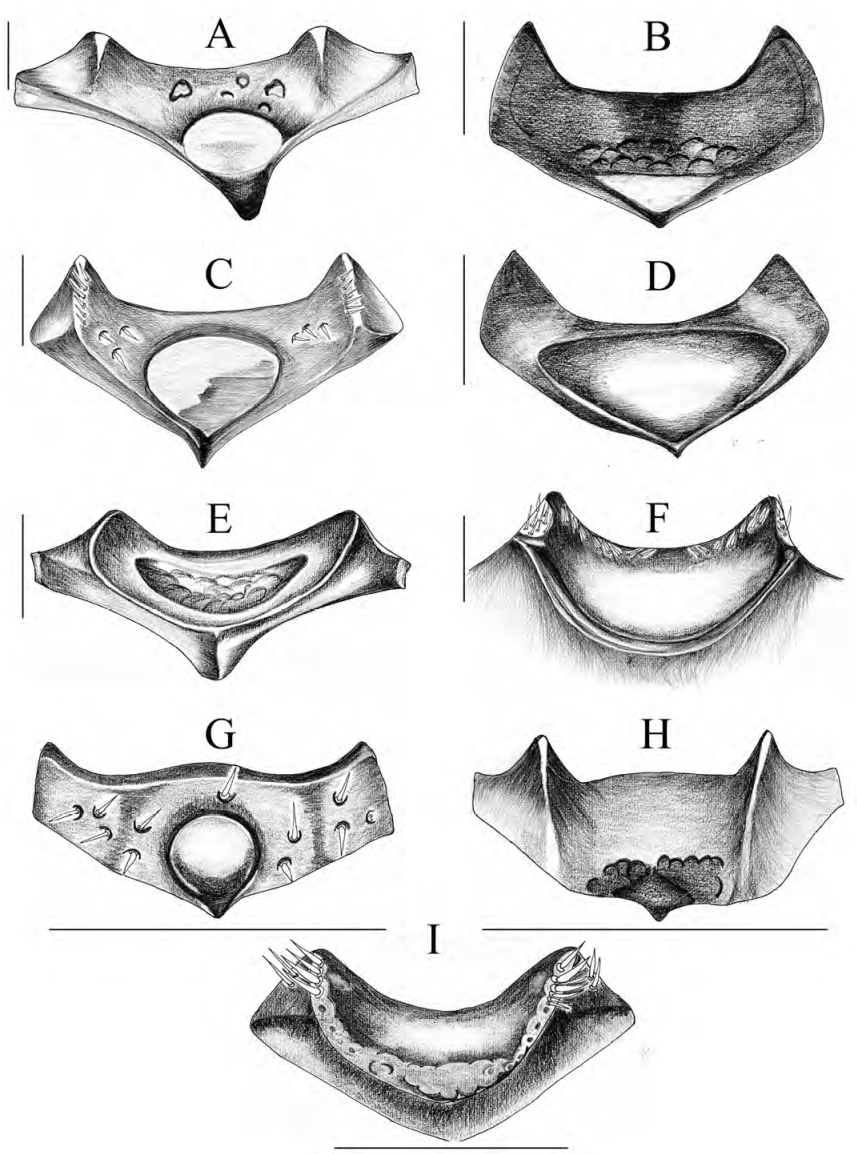
Figura 9 Superficie vertical del clípeo. A) C. altus, B) C. buriticaorum, C) C. encenillensis, D) C. escobari, E) C. foveatus, F) C. mailinae, G) C. parvus, H) C. pumilus, I) C. cephalopunctatus. Escala: 0.1 mm.

Figura 10 Pigidio. A) C. altus, B) C. andradei, C) C. buriticaorum, D) C. campbellorum, E) C. escobari, F) C. foveatus, G) C. humidus, H) C. medinae, I) C. parvus, J) C. pulidoae, K) C. pumilus, L) C. cephalopunctatus. Escala: 0.5 mm.
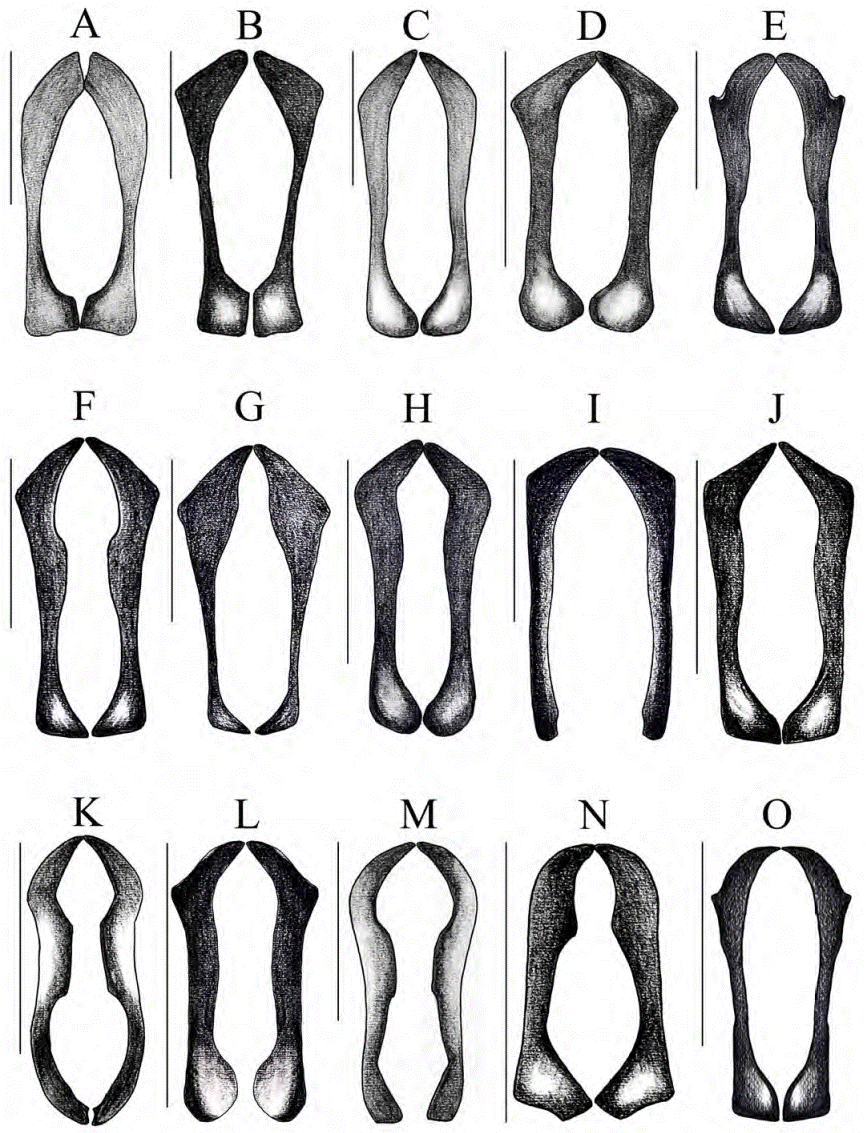
Figura 11 Aedeago, vista caudal. A) C. altus, B) C. andradei, C) C. buriticaorum, D) C. campbellorum, E) C. encenillensis, F) C. escobari, G) C. foveatus, H) C. humidus, I) C. mailinae, J) C. medinae, K) C. parvus, L) C. peckorum, M) C. pulidoae, N) C. pumilus, O) C. cephalopunctatus. Escala: 0.5 mm.

Figura 12 Aedeago, vista lateral. A) C. altus, B) C. andradei, C) C. buriticaorum, D) C. campbellorum, E) C. encenillensis, F) C. escobari, G) C. foveatus, H) C. humidus, I) C. mailinae, J) C. medinae, K) C. parvus, L) C. peckorum, M) C. pulidoae, N) C. pumilus, O) C. cephalopunctatus. Escala: 0.5 mm.
Descripción. Holotipo. Macho. Medidas. Longitud total: 2.70 mm; ancho humeral: 1.68 mm. Cuerpo color cobrizo con cedas castañas en toda la superficie, tibias rojizas (Fig. 3A). Cabeza. Clípeo no proyectado anteriormente. Puntuaciones fuertemente impresas, densas en la región frontal, oceladas con borde incompleto y sedas cortas de color pardo distribuidas en toda la superficie, separadas por una distancia menor a su diámetro (Fig. 4A). Margen anterior del clípeo en vista frontal con 2 dientes cortos ampliamente separados y flexionados; disco central del clípeo cóncavo; sutura clípeo-genal evidente; margen lateral del clípeo en vista frontal convexa entre los dientes y la gena, dando un aspecto ovalado a la cabeza; diente de la gena levemente proyectado. Superficie vertical del clípeo sin fovea (Fig. 9I). Pronoto. Convexo en la parte media y plano lateralmente, con 2 depresiones laterales en el disco. Ángulo anterior y márgenes laterales rectos con declive en el borde anterior. Ángulos posteriores obtusos. Superficie con puntuaciones densas, profundas y oceladas más evidentes en la región lateral e incompletas en la región media y el margen anterior; sedas pardas, cortas y curvas más densas hacia los bordes laterales (Fig. 3A). Élitros. Convexos transversal y longitudinalmente; con 7 estrías fuertemente impresas, formadas por 2 líneas ligeramente onduladas, más evidentes en la región posterior de los élitros. Interestrías con una hilera de puntuaciones con sedas a cada lado adyacente a la estría; sedas pardas, cortas, con ápice curvo. Quilla de la pseudoepipleura fuerte, sin sedas en el borde; superficie sin microescultura, con sedas pequeñas y pardas, distribuidas irregularmente en el margen lateral externo y de mayor longitud en la porción apical; octava estría completa, profunda y formada por puntuaciones más largas que anchas, separadas por una distancia menor a su largo; sin fovea basal. Alas. Alas metatorácicas completamente reducidas. Hipómero sin microescultura y carena posterior, puntuaciones oceladas profundas y ovaladas más densas en el margen de las coxas y separadas entre sí una vez su diámetro hacia el borde lateral. Prosterno. Superficie con puntuaciones oceladas densas en la región media. Mesosterno. Superficie con puntuaciones circulares, pequeñas y dispersas en la región central y profundas y separadas entre sí menos de una vez su diámetro en los márgenes laterales. Sutura mesometaesternal angulada. Superficie del mesepisterno como en el margen lateral del mesosterno. Metasterno. Superficie con microescultura débilmente marcada, casi imperceptible, con puntuaciones pequeñas en la parte anterior y media, separadas entre sí 2 veces su diámetro y hacia la parte posterior y lateral de mayor tamaño y más densos separados menos de una vez su diámetro (Fig. 7O). Metaepisterno con microescultura débilmente marcada y con una hilera de puntuaciones oceladas profundas. Patas. Profémur en vista ventral con puntuaciones oceladas y profundas, dispersas y finas en la mitad anterior, y más densas y profundas en la mitad posterior cerca de la unión al trocánter. Superficie de la protibia en vista ventral sin puntuaciones; con una hilera de puntos con setas al lado derecho de la carena central; con 3 dientes apicales en el borde externo, los 2 primeros más cercanos que el segundo y tercero y con dientes de menor tamaño entre ellos. Muesca preapical en el borde interno presente en machos. Mesofémur y metafémur en vista ventral con puntuaciones oceladas, más anchas que largas y dispersas. Mesotibia dilatada hacia el ápice, en vista ventral con 2 hileras laterales de sedas pardas en los márgenes, desde la región media hacia el ápice. Borde interno de la metatibia curvo en la región media; superficie dorsal plana con una hilera de sedas desde la región media hasta el margen apical interno; superficie ventral convexa; margen interno de la tibia lobulado (Fig. 4E). Abdomen. Superficie del último esternito con puntuaciones grandes y densas. Pigidio convexo; superficie con puntuaciones oceladas; sedas pardas, largas y curvas en la región media; margen de la base con depresión media sin fovea (Fig. 4D). Órgano genital (11O, 12O, 13N 14N, 15N). En vista caudal, parámeros estrechos en la región basal, margen interno recto débilmente curvado hacia el ápice, ápice subcuadrangular; margen anterior recto sin proyecciones (Fig. 11O). Saco interno con 3 escleritos apicales, esclerito basal con un anillo débilmente esclerotizado; con un mango agudo hacia la parte apical, región superior ensanchada en su parte media, ápice romo (Fig.13N). Esclerito placa elongado, con un extremo largo y agudo y el opuesto con 2 proyecciones lobuladas, con una proyección semicircular de mayor tamaño en la parte superior y una proyección subrectangular de menor tamaño en la parte lateral, ápice curvo (Fig.15N). Esclerito elongado con proyecciones setiformes cortas y base ensanchada donde se une una segunda estructura esclerotizada (Fig. 14N).

Figura 13 Esclerito basal. A) C. altus, B) C. andradei, C) C. buriticaorum, D) C. campbellorum, E) C. encenillensis, F) C. escobari, G) C. foveatus, H) C. humidus, I) C. mailinae, J) C. medinae, K) C. parvus, L) C. pulidoae, M) C. pumilus, N) C. cephalopunctatus. Escala: 0.1 mm.

Figura 14 Esclerito elongado. A) C. altus, B) C. andradei, C) C. buriticaorum, D) C. campbellorum, E) C. encenillensis, F) C. escobari, G) C. foveatus, H) C. humidus, I) C. mailinae, J) C. medinae, K) C. parvus, L) C. pulidoae, M) C. pumilus, N) C. cephalopunctatus. Escala: 0.2 mm.
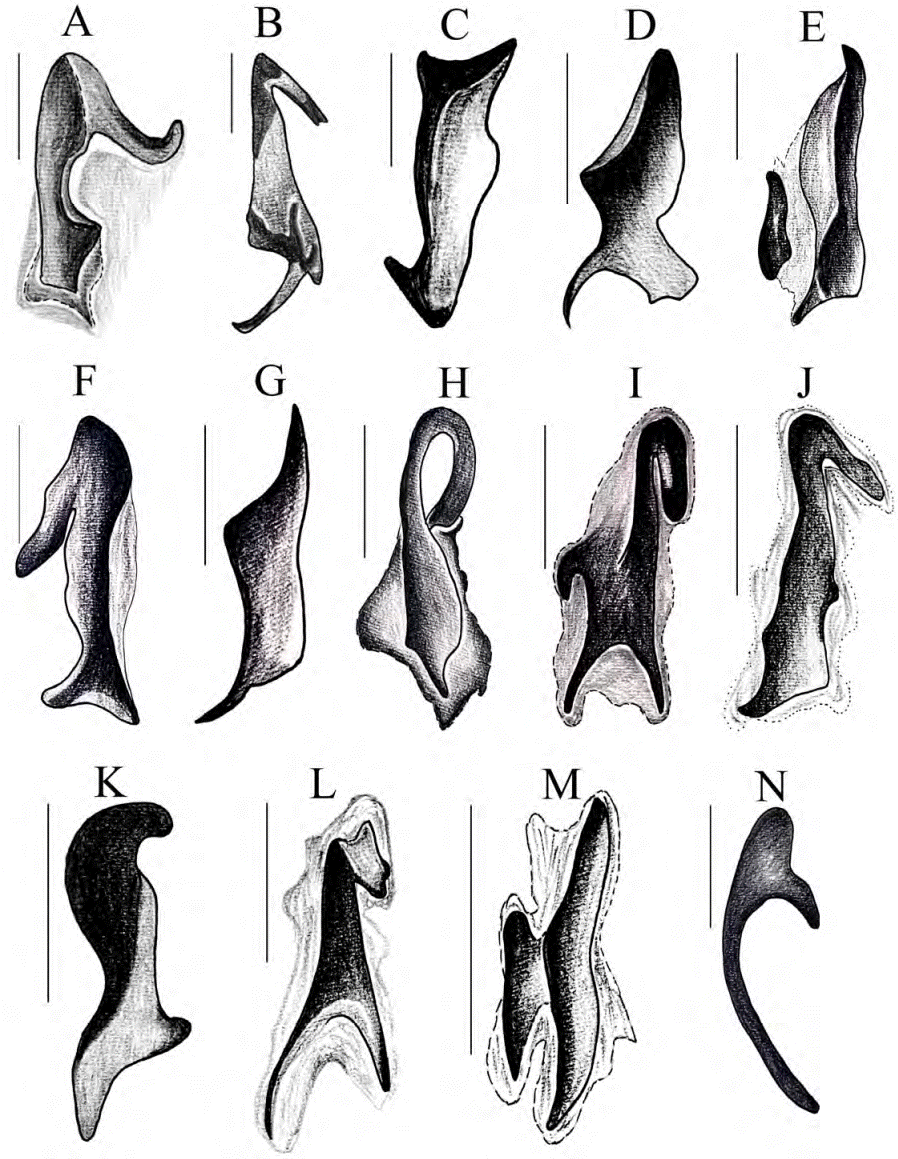
Figura 15 Esclerito placa. A) C. altus, B) C. andradei, C) C. buriticaorum, D) C. campbellorum, E) C. encenillensis, F) C. escobari, G) C. foveatus, H) C. humidus, I) C. mailinae, J) C. medinae, K) C. parvus, L) C. pulidoae, M) C. pumilus, N) C. cephalopunctatus. Escala: 0.2 mm.
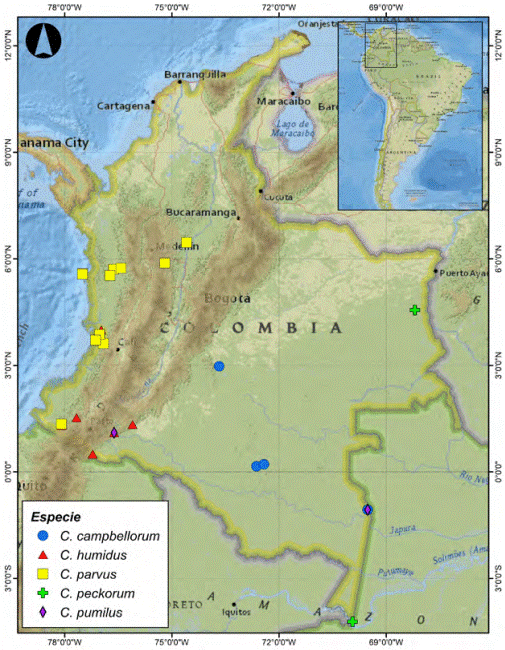
Figura 16 Distribución geográfica de Cryptocanthon en Colombia. Especies con alas posteriores desarrolladas de tierras bajas.
Variación. Machos. Longitud total: 2.63 mm-2.79 mm; ancho humeral: 1.67 mm-1.84 mm. Hembra. Longitud total: 2.45 mm-2.79 mm; ancho humeral: 1.62 mm-1.84mm. Características similares al macho, excepto que el margen interno de la metatibia es casi recto, liso y con ápice interno recto. Dientes clipeales y de la protibia más pronunciados. Protibia sin muesca preapical (Fig. 3D).
Resumen taxonómico
Localidad tipo: Colombia. Santander. Bolívar. Vereda el Espejo. Flanco nororiental de la serranía de las Quinchas. Depósito de tipos, holotipo: Colección Entomológica del Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH-E). Paratipos: Colección Entomológica del Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Villa de Leyva Boyacá (IAvH-E); Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá D.C. (ICN); Museo de Historia Natural Luis Gonzalo Andrade, de la Universidad Pedagógica y Tecnológica de Colombia (UPTC).
Material tipo examinado: holotipo [♂]. Colombia, Santander, Bolívar, vereda el Espejo, bosque contiguo a la estación Santa Rosa, bosque secundario, trampa de excremento humano #T5 (6°08’52” N, 73°48’53.7” O, 1,549 m), 29 nov - 01 dic 2016, Xiomara Villalba C. y C. D. Hernández, IAvH-E-201753. Etiqueta roja impresa: Cryptocanthon cephalopunctatus holotipo Martínez-Revelo, et al. sp. nov. Paratipos: 34 especímenes (20 ♂♂, 14 ♀♀). Colombia, Santander, Bolívar, vereda el Espejo, bosque contiguo a la estación Santa Rosa, borde de bosque, trampa de excremento humano #T3 (6°08’3.5” N, 73°49’16” O, 1,664 m), 29 nov - 01 dic 2016, Xiomara Villalba C. y C. D. Hernández, IAvH-E-201776, 1 ♂; trampa de excremento humano #T7 (6°08’05” N, 73°49’08” O, 1,718 m), 22 - 24 sep 2015, Xiomara Villalba C., J. Gil N., IAvH-E-212447, 1 ♂; bosque secundario, trampa de fruta #T3 (6°08’52” N, 73°48’53.7” O, 1,549 m), IAvH-E-201775, 1♀; trampa de excremento humano #T2, ICN, 1 ♂; trampa de excremento humano #T4, IAvH-E-201781, 1 ♀; UPTC, 1 ♀; trampa de excremento humano #T9 (6°08’48’’ N, 73°48’48’’ O, 1,549m), IAvH-E-201774, 1 ♂; trampa de excremento humano #T4, 29 nov - 01 dic 2016, Xiomara Villalba C. y C. D. Hernández, IAvH-E-201762-67, 3 ♂♂ y 3 ♀♀; ICN, 1 ♀; trampa de excremento humano #T5, IAvH-E-201752, IAvH-E-201754-61, IAvH-E-201777-80, 7 ♂♂ y 6 ♀♀; trampa de excremento humano #T9, IAvH-E-201768-73, 5 ♂♂ y 1 ♀; UPTC, 1 ♂.
Ecología y distribución: esta especie fue recolectada en trampas de caída cebadas con excremento humano y frutas en un fragmento de bosque subandino de aproximadamente 100 hectáreas, entre los 1,549 a 1,718 m altura. Sus poblaciones presentan baja abundancia, en el evento de captura se recolectaron 40 individuos de una muestra total de 3,080 individuos de Scarabaeinae. La especie es conocida únicamente de la localidad tipo (Fig. 17).
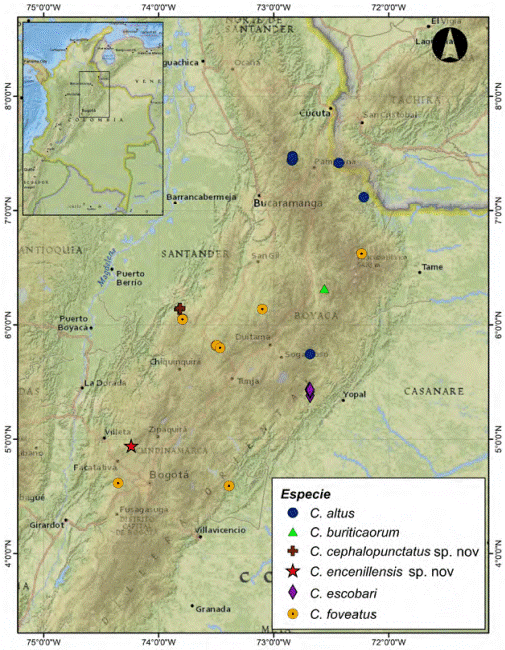
Figura 17 Distribución geográfica de Cryptocanthon en Colombia. Especies braquípteras de la cordillera oriental.
Etimología: el epíteto kephalḗ (griego) + punctatus (latín) hace referencia a las puntuaciones fuertemente impresas en la superficie de la cabeza, característico de la especie.
Comentarios taxonómicos
Cryptocanthon cephalopunctatus sp. nov. se caracteriza por la siguiente combinación de caracteres: margen lateral del clípeo convexo entre los dientes y la gena; superficie vertical del clípeo y epipleura sin fóvea (Figs. 4F, 9I); alas meta-torácicas reducidas e hipómero sin carena posterior. Está relacionada con las especies C. medinae, C. andradei y C. punctatus por presentar la superficie vertical del clípeo sin fovea, margen lateral del pronoto recto y borde interno de la metatibia curvada en la región media. Se distingue de las demás especies por que la protibia del macho presenta muesca preapical (diferente a C. punctatus); el hipómero no presenta carena basal longitudinal (diferente a C. medinae y C. andradei), la cabeza tiene puntuaciones fuertemente impresas, densas y oceladas en toda la superficie (Fig. 4A); margen interno de la tibia posterior lobulada (Fig. 4E); pigidio con estrecha depresión media en la base, sin fovea (Fig. 4D); ápice de los parámeros en vista caudal subcuadrangular (Fig. 11O); esclerito placa elongado, con un extremo largo y agudo y el opuesto con 2 proyecciones lobuladas, con una proyección semicircular de mayor tamaño en la parte superior y una proyección subrectangular de menor tamaño en la parte lateral, ápice curvo (Fig. 15N).
Cryptocanthon encenillensis Martínez-Revelo, Torres y Neita sp. nov. (Figs. 3C, D, 5, 9C, 11E, 12E, 13E, 14E, 15E, 17) http://zoobank.org/urn:lsid:zoobank.org:act:E7C18B8F-925D-4C25-83ED-6492B0B57396
Descripción. Holotipo. Macho. Medidas. Longitud total: 3.4 mm; ancho humeral: 2.5 mm. Cuerpo color cobrizo brillante, con cedas castañas en toda la superficie y márgenes menos esclerotizados de color rojizo (Fig. 3C). Cabeza. Clípeo no proyectado anteriormente. Superficie con puntación ocelada, separadas por una distancia menor a su diámetro en la región media y con mayor distancia hacia el borde anterior (Fig. 5A). Margen anterior del clípeo en vista frontal con 2 dientes cortos ampliamente separados y flexionados; disco del clípeo cóncavo; sutura clípeo-genal evidente; margen lateral del clípeo en vista frontal convexa entre los dientes y la gena, dando un aspecto ovalado a la cabeza; diente de la gena levemente proyectado. Superficie vertical del clípeo ampliamente foveada (Fig. 9C). Pronoto. Convexo en la parte media y plano lateralmente, con 2 depresiones laterales en el disco. Ángulo anterior y márgenes laterales rectos con un declive en el borde anterior. Ángulos posteriores obtusos. Superficie con puntuación densa y profunda, ocelada en los bordes posterolaterales y simple en la región media y el margen anterior; sedas pardas, cortas y curvas más densas hacia los bordes laterales (Fig. 3C, D). Élitros. Convexos transversal y longitudinalmente; superficie con 7 estrías fuertemente impresas, formadas por 2 líneas ligeramente onduladas, más evidentes hacia la región posterior de los élitros. Interestrías con una fila de puntuaciones con sedas a cada lado adyacente a la estría; sedas pardas y cortas. Quilla de la pseudoepipleura fuerte, sin sedas en el borde; superficie sin microescultura, con sedas pequeñas y pardas, distribuidas irregularmente en el margen lateral externo y de mayor longitud hacia el ápice; octava estría completa, profunda y formada por puntuaciones más largas que anchas, separadas por una distancia menor a su largo; sin fovea basal (Fig. 5F). Alas. Alas metatorácicas completamente reducidas. Hipómero sin microescultura y carena posterior (Fig. 5B), puntuaciones oceladas profundas y ovaladas más densas en el margen de las coxas y separadas entre sí una vez su diámetro hacia el borde lateral. Prosterno. Superficie con puntuaciones oceladas densas en la región media. Mesosterno. Superficie con puntuaciones circulares, pequeñas y dispersas en la región central, profundas y separadas entre sí menos de una vez su diámetro en los márgenes laterales. Sutura meso-metaesternal angulada. Mesepisterno con la misma superficie que el margen lateral del mesosterno. Metasterno. Superficie con microescultura débilmente marcada, casi imperceptible, en la parte anterior y media con puntuaciones pequeñas separadas entre sí 2 veces su diámetro y hacia la parte posterior y lateral de mayor tamaño y más densos separados menos de una vez su diámetro (Fig. 5C). Metaepisterno con microescultura débilmente marcada y con una hilera de puntuaciones oceladas profundas. Patas. Profémur en vista ventral con puntuaciones profundas, dispersas y finas en la mitad anterior y más densas y oceladas en la mitad posterior cerca de la unión al trocánter. Superficie de la protibia en vista ventral sin puntuaciones; con fila de puntuaciones con sedas al lado derecho de la carena central; con 3 dientes apicales en el borde externo, los 2 primeros más cercanos que el segundo y tercero y con dientes de menor tamaño entre ellos. Muesca preapical en el borde interno presente en machos. Mesofémur y metafémur en vista ventral con puntuaciones finas y dispersas. Mesotibia dilatada hacia el ápice, en vista ventral con 2 filas laterales de sedas pardas en los márgenes, desde la región media hacia el ápice. Borde interno de la metatibia fuertemente curvada en la región media; superficie dorsal plana con fila de sedas desde la región media que termina en el margen apical interno; superficie ventral convexa; margen interior de la tibia truncado formando un ángulo obtuso con el margen apical, donde se ubica un diente corto y romo (Fig. 5E). Abdomen. Superficie del último esternito con puntuaciones grandes y densas. Pigidio convexo; superficie con puntuaciones oceladas; sedas pardas, cortas y curvas en la región media; margen de la base con depresión media que termina en fovea (Fig. 5D). Órgano genital (Figs. 11E, 12E, 13E, 14E, 15E). Parámeros en vista caudal estrechos hacia la parte basal, margen interno con un ensanchamiento en la parte media que se curva hacia el ápice y convergen en punta aguda; ángulo del borde externo del ápice obtuso (Fig. 11E). Saco interno con 3 escleritos apicales, esclerito basal con un anillo débilmente esclerotizado; con un mango agudo hacia la parte apical, ensanchado en la parte media terminando abruptamente hacia el cuarto apical, donde forma una pequeña saliente y se estrecha hacia el ápice (Fig. 13E); esclerito placa de forma rectangular con un extremo agudo y el opuesto con una proyección lateral que termina en punta (Fig. 15E). Esclerito elongado con proyecciones setiformes largas y base ensanchada donde se une una segunda estructura esclerotizada (Fig. 14E).
Variación: hembra. Longitud total: 3.73 mm; ancho humeral: 2.58 mm. Características similares al macho, a excepción del borde interno de la metatibia menos curvo que en el macho y sin diente preapical. Dientes clipeales y de la protibia más pronunciados. Protibia sin muesca preapical (Fig. 3D).
Resumen taxonómico
Localidad tipo: Colombia. Cundinamarca. San Francisco. RN la Nube. Corredor de bosques andinos de los páramos Sumapaz-Chingaza-Iguaque. Depósito de tipos: Colección Entomológica del Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH-E).
Material tipo examinado: holotipo [♂]. Colombia, Cundinamarca, San Francisco, Reserva la Nube, bosque de encenillares (4°56’36.9” N, 74°14’21.0” O, 2,707 m), trampa de excremento humano #T1 (24), 12 - 16 dic 2016, J. C. Neita, IAvH-E-184501; etiqueta roja impresa: Cryptocanthon encenillensis holotipo Martínez-Revelo, et al. sp. nov. Paratipo: igual información que el holotipo, IAvH-E-184500, 1 ♀.
Ecología y distribución: esta especie presenta baja abundancia, en 37 eventos de captura con trampas de caída cebadas con excremento humano, sólo se recolectaron 2 individuos de un total de 1,190 ejemplares. C. encenillensis coexiste con 6 especies de Scarabaeinae comunes en bosques andinos: un representante del género Canthidium, Dichotomius inachoides (Felsche, 1901), Homocopris achamas (Harold, 1867), Ontherus brevicollis Kirsch, 1871, Onthophagus curvicornis (Latreille, 1811) y Uroxys coarctatus Harold, 1867, siendo la especie con menor tamaño y abundancia del ensamble. Su distribución se restringe a la localidad tipo (Fig. 17), ubicada en un paisaje altamente fragmentado perteneciente al corrector de conectividad de los Parques Naturales Nacionales Chingaza, Sumapaz y el Santuario de Flora y Fauna Iguaque, entre los departamentos de Boyacá y Cundinamarca. El fragmento de bosque donde se capturó la especie tiene un área cercana a los 5 km2, caracterizado por la dominancia de encenillos (Weinmannia spp.) y una alta riqueza de plantas de las familias Asteraceae, Ericaceae y Melastomataceae.
Etimología: el epíteto “encenillensis” hace referencia a los bosques de encenillo, donde la especie fue descubierta.
Comentarios taxonómicos
Cryptocanthon encenillensis se caracteriza por la siguiente combinación de caracteres: margen lateral del clípeo en vista frontal convexo entre los dientes y la gena; superficie vertical del clípeo ampliamente foveada (Fig. 9C); epipleura sin fovea basal (Fig. 5F); alas metatorácicas reducidas; hipómero sin carena posterior (Fig. 5B) y margen basal del pigidio con depresión media que termina en fóvea (Fig. 5D). La especie se relaciona con C. foveatus por presentar el margen lateral del pronoto recto; fóvea profunda en la superficie vertical del clípeo y en el margen basal del pigidio; fémures y tibias posteriores curvados. Se distingue por la fovea de la superficie vertical del clípeo amplia entre el ápice de los dientes clipeales; la curvatura media de la tibia posterior más acentuada y el margen interno con un diente preapical romo (Fig. 5E), el margen interno de los parámeros en vista caudal se ensanchan en la parte media y se curvan hacia el ápice que converge en punta aguda mientras el ángulo del borde externo es obtuso (Fig. 11E); esclerito basal ensanchado en la parte media terminando abruptamente hacia el cuarto apical, donde forma una pequeña saliente y se estrecha hacia el ápice (Fig.13E), el esclerito elongado tiene proyecciones setiformes muy largas (Fig. 15E) y esclerito placa de forma rectangular con un extremo agudo y el opuesto con una proyección lateral que termina en punta (Fig. 14E).
Cryptocanthon mailinae Martínez-Revelo, Torres y Neita sp. nov. (Figs. 3E, F, 6, 8A, 9F, 11I, 12I, 13I, 14I, 15I, 18) http://zoobank.org/urn:lsid:zoobank.org:act:2BC34AB1-BEC5-4023-A691-750A9CA4A639

Figura 18 Distribución geográfica de Cryptocanthon en Colombia. Especies braquípteras de la cordillera central y occidental.
Descripción. Holotipo. Macho. Longitud total: 3.7 mm; ancho humeral: 2.5 mm. Cuerpo color rojizo brillante, con cedas castañas en toda la superficie (Fig. 3D). Cabeza. Clípeo no proyectado anteriormente. Superficie con puntuaciones oceladas, separadas por una distancia menor a su diámetro en la región central y con mayor distancia hacia el borde anterior (Fig. 6A). Margen anterior del clípeo en vista frontal con 2 dientes cortos ampliamente separados y flexionados; disco del clípeo cóncavo; sutura clípeo-genal evidente; margen lateral del clípeo convexa entre los dientes y la gena, dando un aspecto ovalado a la cabeza; diente de la gena levemente proyectado. Superficie vertical del clípeo sin fovea (Fig. 9F). Pronoto. Convexo en la parte media y plano lateralmente; depresiones laterales superficiales en el cuarto posterior. Ángulo anterior del pronoto recto; márgenes laterales curvos en la base. Ángulos posteriores obtusos (Fig. 3E, F). Superficie con puntuaciones densas y profundas, oceladas en los bordes posterolaterales y simples en la región central y el margen anterior. Con sedas pardas, cortas y curvas más densas hacia los bordes laterales. Élitros. Convexos transversal y longitudinalmente; con 7 estrías profundamente marcadas, formadas por 2 líneas con ondulaciones evidentes, disco y la región apical con círculos. Interestrías con una fila de puntuaciones con sedas a cada lado adyacente a la estría; sedas pardas, cortas y rectas. Quilla de la pseudoepipleura fuerte, sin sedas evidentes en el borde. En vista anterolateral, borde externo de la última interestría con una hilera de sedas cortas y curvas. Superficie de la pseudoepipleura con microescultura fuertemente impresa; octava estría completa y fuertemente impresa más evidente en la región anterior, compuesta por 2 líneas que rodean puntuaciones semicirculares grandes, distanciadas por una distancia menor a su diámetro; sin fovea basal (Fig. 6F). Alas. Alas metatorácicas completamente reducidas. Hipómero con microescultura punticulada evidente, sin carena posterior, puntuaciones oceladas profundas y ovaladas más densas en el margen de las coxas y separadas entre sí una vez su diámetro hacia el borde lateral (Fig. 6B). Prosterno. Superficie con puntuaciones oceladas en el borde lateral junto al hipómero. Mesosterno. Superficie con microescultura evidente, en la parte media con puntuaciones dispersas y hacia los lados con puntuaciones grandes, profundas y separadas entre sí menos de una vez su diámetro. Sutura meso-metaesternal bien angulada. Mesepisterno con la misma superficie que los bordes laterales del mesosterno. Metasterno. Superficie con microescultura foveolada fuertemente impresa; región anterior y media lisa, región posterior y lateral con puntuaciones grandes y densas, separadas menos de una vez su diámetro (Fig. 6C). Metaepisterno con microescultura evidente y con una hilera de punturas similares a los de la zona lateral del metaesterno. Patas. Profémur en vista ventral con puntuaciones profundas, dispersas y finas en la mitad anterior y más densas y oceladas en la mitad posterior cerca de la unión al trocánter. Superficie de la protibia en vista ventral lisa; con una carena central marcada, glabra; con 3 dientes apicales en el borde externo, los 2 primeros más cercanos que el segundo y tercero y con dientes de menor tamaño entre ellos. Muesca preapical en el borde interno presente en machos. Mesofémur y metafémur en vista ventral con puntuaciones finas y dispersas. Mesotibia dilatada hacia el ápice, en vista ventral con 2 hileras laterales de sedas pardas en los márgenes laterales, desde la región media hacia el ápice. Metatibia débilmente curvada; superficie dorsal plana con una hilera de sedas desde la región media hasta el margen interno; margen lateral interno fuertemente crenulado evidente desde la región media hacia el ápice, margen lateral apical externo en vista dorsal agudo (Fig. 6E). Abdomen. Superficie del último esternito con puntuaciones grandes y densas. Pigidio convexo con puntuaciones oceladas grandes, con sedas conspicuas; margen de la base con una estrecha depresión media (Fig. 6D). Órgano genital (Figs. 11I, 12I, 13I, 14I, 15I). Parameros subrectangulares con falobase subcuadrangular (Fig. 12I). En vista caudal, estrechos en la parte media y ensanchados hacia el ápice, ápice truncado (Fig. 11I). Saco interno con 3 escleritos apicales, esclerito con un anillo débilmente esclerotizado; con un mango agudo hacia la parte apical, curvo y recto en la parte media, terminando hacia la zona basal con una proyección que se ensancha hacia el ápice (Fig. 13I). Esclerito placa con un extremo en forma de gancho con ápice romo, en el extremo opuesto el ápice con forma triangular, región media escasamente esclerotizada (Fig. 15I). Esclerito elongado con proyecciones setiformes y base poco ensanchada (Fig. 14I).
Variación: macho. Longitud total: 3.25 mm - 4.05 mm; ancho humeral 2.54 mm - 3.04 mm. Hembra. Longitud total: 3.19 mm - 3.96 mm; ancho humeral: 2.53 mm - 2.86mm. Características similares al macho, excepto que el borde interno de la metatibia es liso. Dientes clipeales y de la protibia más pronunciados. Protibia sin modificación.
Resumen taxonómico
Localidad tipo: Colombia. Antioquia. Jardín. RN Mesenia. Cordillera Occidental de los Andes. Depósito de tipos: holotipo. Colección Entomológica del Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH-E). Paratipos. Museo Francisco Luis Gallego, Universidad Nacional de Colombia, campus Medellín, Antioquia (MEFLG), Colección Entomológica Universidad de Antioquia, Medellín, Colombia (CEUA).
Material tipo examinado: holotipo [♂]. Colombia, Antioquia, Jardín, Reserva Natural Mesenia, Forest, trampa de excremento humano #P5 (5°29’34” N, 75°53’53” O, 2,506 m), jul 2012, F. Edwards, IAvH-E-201733; etiqueta roja impresa: Cryptocanthon mailinae holotipo Martínez-Revelo, et al. sp. nov. Paratipos: 28 especímenes (17♂♂, 11♀♀): 2♀♀ con datos igual al holotipo IAvH-E-184488-89; #P2 (5°29’30” N, 75°43’8” O, 2,609 m), E. Felicity, IAvH-E-201720, IAvH-E-201722-25, IAvH-E-184480, 4♂♂ y 2♀♀; CEUA, 1♂ y 1♀; #P3 (5°29’30” N, 75°54’8” O, 2,609m), IAvH-E-201726-32, IAvH-E-184481-86, 8♂♂ y 5♀♀; #P5 (05°29’34” N, 75°53’53” O, 2,609 m), E. Felicity (MFLG), 1 ♂ y 1 ♀. Chocó, El Cármen de Atrato, Reserva Natural Las Tangaras, Forest, #P2 (5°50’2” N, 76°10’9” O, 1,951m), E. Felicity, IAvH-E-201721, 1♂. Risaralda, Pueblo Rico PNN Tatamá, Cuchilla La línea, #28 (05°8’54” N, 76°02’00” O, 2,620-2,680 m), 21 - 23 ago 2004, A. Pulido y E. González, IAvH-E-212450, 1 ♂; #29, IAvH-E-212449, 1♂.
Ecología y distribución: Esta especie fue recolectada en trampas de caída cebadas con excremento humano. Se distribuye en bosques andinos de la cordillera occidental en los departamentos de Antioquia, Chocó y Risaralda (Fig. 18).
Etimología: esta especie es dedicada a la investigadora Mailyn Adriana González, responsable de las campañas de caracterización biológica en zonas afectadas por el conflicto armado, promotora y defensora de las campañas de exploración y recolección de especímenes en el país.
Comentarios taxonómicos
Se caracteriza por la siguiente combinación de caracteres: margen lateral del clípeo en vista frontal convexo entre los dientes y la gena; superficie vertical del clípeo y epipleura sin fovea (Fig. 9F); alas metatorácicas reducidas; pigidio con una estrecha depresión media en la base (Fig. 6D). Esta especie está relacionada con C. pulidoae por presentar los márgenes laterales del pronoto curvos (Figs. 2E, 8B); sedas largas en la superficie dorsal de pronoto y élitros; metatibias con el borde interno débilmente curvas. Se distingue por la superficie de la epipleura y metaesterno con microescultura foveolada fuertemente impresa (Fig. 6C, F); margen lateral interno de la tibia posterior fuertemente crenulado, más evidente desde la región media hacia el ápice (Fig.6E); ápice de los parámeros en vista caudal de forma subrectangular (Fig. 11I); esclerito basal con proyección que se ensancha hacia el ápice (Fig. 12I) y esclerito placa con un extremo en forma de gancho con ápice romo, en el extremo opuesto el ápice tiene forma de triángulo, la región media escasamente esclerotizada (Fig. 15I).
Discusión
El género Cryptocanthon se encuentra distribuido ampliamente en el país, cubriendo un rango altitudinal entre los 25 a 3,000 m en las regiones naturales de la Orinoquia, Amazonía, Pacífico y Andes, sin representantes en las regiones insular y del Caribe. De las 15 especies registradas, 3 se comparten con otros países: C. campbellorum con Brasil y Perú, C. humidus con Panamá y C. peckorum con Brasil. La distribución conocida de las restantes especies se encuentra restringida a Colombia.
La cordillera de los Andes ha determinado un patrón de distribución altitudinal claramente definido en el género, observándose 3 grupos de especies: el primero está conformado por especies que se caracterizan morfológicamente por presentar las alas posteriores desarrolladas y se distribuyen en tierras bajas por debajo de los 1,540 m de altura (Fig. 16). Las especies C. humidus y C. parvus cubren una franja altitudinal superior a los 1,000 m, mientras que C. campbellorum, C. peckorum y C. pumilus tienen una mayor restricción altitudinal. Sin embargo, C. pumilus está registrada en la serranía de los Churumbelos (Putumayo), ampliando su rango altitudinal hasta los 860 m (tabla 1). El segundo grupo está conformado por especies braquípteras que se distribuyen en la cordillera oriental por encima de los 1,549 m de altura. Se observa una segregación de especies distribuidas hacia las vertientes de la cordillera: C. altus y C. escobari presentes únicamente en la vertiente oriental, C. buriticaorum, C. encenillensis y C. cephalopunctatus en la vertiente occidental y poblaciones de C. foveatus en las 2 vertientes (Fig. 17). El tercer grupo está conformado por especies braquípteras distribuidas en la cordillera occidental por encima de los 1,898 m de altura, el grupo incluye a C. andradei que también se registra en la vertiente occidental de la cordillera central y es la única especie presente en los Andes centrales. Las especies C. medinae y C. pulidoae solo se encuentran registradas en la vertiente oriental y C. mailinae presenta poblaciones en las 2 vertientes (Fig. 18). Las especies braquípteras en los diferentes ramales de la cordillera Andina se encuentran restringidas a estrechas franjas altitudinales que no sobrepasan los 600 m, excepto C. altus con un rango mayor a 700 m (tabla 1).
Tabla 1 Lista de especies de Cryptocanthon Balthasar presentes en Colombia.
| Especies | Departamento | Rango altitudinal | Colección |
| C. altus Howden | Boyacá, Norte de Santander, Santander | 2,240-3,000 | CEUA, IAvH-E, UPTC |
| C. andradei Arias y Medina | Antioquia, Risaralda | 1,898-2,556 | CEUA, MEFLG, MEUV, IAvH-E |
| C. buriticaorum Arias y Medina | Boyacá | 2,378 | IAvH-E |
| C. campbellorum Howden | Caquetá, Meta, Vaupés | 100-250 | CECC-CALT, IAvH-E |
| C. cephalopunctatus Martínez- | Santander | 1,549-1,718 | IAvH-E, ICN, UPTC |
| Revelo, Torres y Neita sp. nov. | |||
| C. encenillensis Martínez-Revelo, | Cundinamarca | 2,707 | IAvH-E |
| Torres y Neita sp. nov. | |||
| C. escobari Cook | Boyacá | 2,000-2,200 | IAvH-E, UPTC |
| C. foveatus Cook | Boyacá, Cundinamarca, Santander | 2,250-2,850 | CECC-CALT, IAvH-E, UPTC |
| C. humidus Howden | Caquetá, Nariño, Putumayo, Valle del | 500-1,540 | CECC-CALT, CEUN-PSO, IAvH-E |
| Cauca | |||
| C. mailinae Martínez-Revelo, | Antioquia, Chocó, Risaralda | 1,951-2,620 | CEUA, IAvH-E, MEFLG |
| Torres y Neita sp. nov. | |||
| C. medinae Cook | Valle del Cauca | 2,030-2,200 | IAvH-E |
| C. parvus Howden | Antioquia, Chocó, Nariño, valle del | 25-1,214 | IAvH-E, UPTC |
| Cauca | |||
| C. peckorum Howden | Amazonas, Vichada | 230-270 | IAvH-E |
| C. pulidoae Arias y Medina | Risaralda | 2,200-2,680 | IAvH-E, UPTC |
| C. pumilus Cook | Putumayo, Vaupés | 100-860 | IAvH-E |
Teniendo en cuenta la regionalización biogeográfica del Neotrópico (Morrone, 2014), el 73% de las especies colombianas del género presenta distribución restringida a una provincia biogeográfica. Aunque la delimitación de niveles de endemismo teniendo en cuenta la extensión de ocurrencia y área de ocupación es limitada, se puede considerar que las especies braquípteras son exclusivas para el país, probablemente como resultado el aislamiento geográfico entre y dentro de las cadenas montañosas. Dos especies tienen distribuciones amplias que cubren como máximo 3 provincias: C. humidus se distribuye en mayor proporción en la provincia del Chocó-Darién, con algunos registros en las provincias de Cauca y Napo y C. parvus se distribuye principalmente en la provincia de Chocó-Darién, con algunos registros en la provincia del Cauca y Magdalena. Dos especies se encuentran distribuidas como máximo en 2 provincias: C. pumilus en las provincias de Imerí y Napo y C. foveatus en la provincia de Páramo y Magdalena. El Cauca es la provincia biogeográfica con mayor riqueza de especies: C. andradei, C. mailinae, C. medinae y C. pulidoae se distribuyen únicamente en esta región. C. altus, C. buriticaorum y C. escobari se encuentran restringidas a la provincia del Páramo. C. encenillensis y C. cephalopunctatus se distribuyen solo en la provincia de Magdalena y C. campbellorum y C. peckorum se registran exclusivamente en la provincia Imerí.
Colombia es el país con mayor riqueza de especies de Cryptocanthon, en total 15 se reconocen en el país, 5 son macrópteras con distribuciones geográficas amplias y en contraste, 10 especies presentan reducción alar y distribución geográfica restringida. Estas características tienen importantes implicaciones en términos de conservación debido a que confieren a las poblaciones un alto grado de vulnerabilidad y riesgo de extinción, teniendo en cuenta la acelerada transformación a las que están sometidos los ecosistemas andinos colombianos. La braquiptería se presenta en distintos géneros de Scarabaeinae, probablemente como resultado de convergencias morfológicas que pueden relacionarse con ambientes montanos ubicados en antiguas áreas de emergencia geológica, donde confluyen especies braquípteras, por ejemplo, la cordillera de Talamanca (Costa Rica) y las sierras Madre (México) con los géneros Onthophagus, Canthidium, Cryptocanthon y Ateuchus (Gasca et al., 2018; Kohlmann et al., 2019).
Las hipótesis que se han planteado para explicar la braquiptería en insectos de hábitats montanos incluyen factores climáticos como la disminución de la temperatura y los cambios en las propiedades físicas del aire, que hacen que el vuelo sea energéticamente más costoso (Dillon et al., 2006). Puede ser una respuesta a la estabilidad y persistencia histórica del hábitat, donde presumiblemente no es necesario un alto grado de dispersión, favoreciendo la pérdida del vuelo (Darlington, 1943; Roff, 1990). Otra explicación hace referencia al costo energético de mantener la capacidad de vuelo que excede al costo de reproducción y supervivencia, por la limitación energética del hábitat en altas elevaciones (Ikeda et al., 2012). Kohlmann et al. (2019) mencionan 2 hipótesis más citando a Lawrence et al. (1991) y Kavanaugh (1985), que consideran a la braquíptera como un valor selectivo en insectos que adoptaron una forma de vida sedentaria, críptica o parasítica o que viven en una montaña, isla o altas elevaciones, o también puede ser que la condición de desarrollo completo de las alas (condición macróptera) sea considerada una característica ancestral. Dada la variedad de argumentos para explicar la braquiptería, se hace necesario evaluar las anteriores hipótesis o proponer nuevas para entender cómo esta adaptación surgió en las especies de Cryptocanthon en los Andes colombianos.
El hábitat de Cryptocanthon en Colombia está estrictamente relacionado con ecosistemas que se caracterizan por coberturas boscosas en buen estado de conservación, alta concentración de hojarasca y humedad. Las poblaciones de las especies braquípteras principalmente de la cordillera oriental se encuentran restringidas a reductos de bosques de roble (Quercus ssp.) y encenillos (Weinmannia spp.) y las demás braquípteras a estrechas franjas de bosques andinos, áreas que se encuentran en alto grado de amenaza debido a la alta tasa de deforestación en los Andes. Aunque algunas especies se encuentran representadas en áreas protegidas como C. altus en el Parque Natural Nacional Tamá y Parque Natural Regional Sisavita (Norte de Santander), C. foveatus en el Parque Natural Nacional Nevado del Cocuy (Boyacá), C. andradei, C. mailinae sp. nov. y C. pulidoae en el Parque Natural Nacional Tatamá (Antioquia - Risaralda), C. medinae en el Parque Natural Nacional Farallones de Cali (valle del Cauca) y C. campbellorum en el Parque Natural Nacional Serranía de Chiribiquete (Caquetá), la mayor parte de las especies colombianas no cuentan con acciones de conservación concretas.
De las 15 especies de Cryptocanthon registradas en Colombia, 3 especies han sido evaluadas en la lista roja de la Unión Internacional para la Conservación de la Naturaleza (UICN): C. campbellorum se encuentra categorizada en preocupación menor (LC), C. foveatus casi amenazada (NT) y C. altus se encuentra en peligro (EN) (Vaz de Mello et al., 2013a, b, 2014). Debido al escaso conocimiento sobre el estado actual de las poblaciones y el riesgo de extinción de las especies, principalmente las que presentan condición braquíptera y que las evaluaciones previas fueron realizadas por expertos internacionales excluyendo información producida en Colombia, se hace necesario realizar nuevas evaluaciones incluyendo información proveniente de colecciones entomológicas colombianas y expertos nacionales.
Clave para la identificación de las especies de Cryptocanthon de Colombia, basada en caracteres morfológicos externos de machos
Alas posteriores desarrolladas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
1’. Alas posteriores atrofiadas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
Pigidio sin tumescencia media o cresta longitudinal (4D, 5D, 6D, 10A, B, E, F, G, H, J, L); fovea de la superficie vertical del clípeo ausente (Fig. 9F, H, I) . . . . . . . . . . . . . . . . . . . . . . . . . . C. humidus Howden
2’. Pigidio con tumescencia media o cresta longitudinal ((Fig. 10C, D, I, K); fovea de la superficie vertical del clípeo presente . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
Hipómero con carena basal (Fig. 8F); metatibia arqueada con margen crenulado (Figs. 6E, 7D, J) . C. parvus Howden
3’...............................................
Hipómero sin carena basal (Fig. 8E); metatibia casi recta con margen liso (7E, F, H, K) . . . . . . . . . . . . 4 Fovea de la superficie vertical del clípeo ausente (Fig. 9F, H, I) . . . . . . . . . . . . . . . C. pumilus Cook
4’. Fovea de la superficie vertical del clípeo circular (Fig. 9G) . . . . . . . . . . . . . . . . . . . . . . 5 Porción inflexa del élitro con fovea basal (Fig. 8G) . . . . . . . . . . . . . . . . .C. campbellorum Howden
5’. Porción inflexa del élitro sin fovea basal (Figs. 4F, 5F, 6F, 8H) . . . . . . . . . . . . C. peckorum Howden Cabeza y pronoto con puntuaciones densas y profundas (Fig. 4A); sutura meso-metaesternal cónica (Fig. 7N, O) . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. cephalopunctatus sp. nov.
6’. Puntuaciones de cabeza y pronoto menos densa y superficial (Figs. 5A, 6A); sutura meso-metaesternal angulada (Figs. 5C,6C,7L,M).........................................7 Superficie vertical del clípeo sin fovea (Fig. 9F, H, I) . . . . . . . . . . . . . . . . . . . . . . . . . . 8
7’. Superficie vertical del clípeo con fovea (fig. 9A-E, G) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
Hipómero con carena basal (fig. 8F) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
8’. Hipómero sin carena basal (figs. 4B, 5B, 6B, 8E) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
Estría de la epipleura ausente (fig. 5F); ápice interno de la metatibia proyectada lateralmente (fig.7D) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. andradei Arias y Medina
9’. Estría de la epipleura, presente e incompleta en la base (figs. 6F, 8G, H); ápice interno de la metatibia lobulado (fig. 7I) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. medinae Cook
Ángulos posteriores del pronoto curvos (fig. 8B); margen interno de la metatibia liso (fig. 7E-I, K) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. pulidoae Arias y Medina
10’ Ángulos posteriores del pronoto rectos (fig. 8A); margen interno de la metatibia crenulado (figs. 6E, 7D, J) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. mailinae sp. nov.
Fovea de la superficie vertical del clípeo estrecha, ubicada hacia el margen inferior (fig. 9A, B, E) . . . . . . . . . . . . . 12
11’ Fovea de la superficie vertical del clípeo amplia, ubicada en la región central (fig. 9C, D, G) . . . . . . . . . . . . . . 14
Estrías elitrales casi rectas, más evidente en el tercio basal (fig. 8C) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. altus Cook
12’. Estrías elitrales onduladas desde la base (fig. 8D) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
Metatibia con margen interno recto y liso (Fig. 7E, F, H), ápice externo proyectado sobrepasando los 2 primeros tarsómeros (Fig. 8E) . . . . . . . . . . . . . . . . . . . . . . . . . . .C. buriticaorum Arias y Medina
13’. Metatibia con margen interno angulado y crenulado (Figs. 6E, 7D, J), ápice externo trunco . . C. foveatus Cook
Metatibia con margen interno recto (Fig. 7E, F, H), ápice interno agudo . . . . . . . . . . . .C. escobari Cook
14’. Metatibia con margen fuertemente angulado (Figs. 5E, 7D, G, I, J), ápice interno con diente preapical (Fig. 5E) .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. encenillensis sp. nov.

