











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
La introducción de una especie exótica a un ecosistema con poblaciones nativas puede generar cambios en las relaciones ecológicas a causa de la competencia por recursos (Sanders et al., 2003). El grado de competencia entre especies nativas e invasoras obedece principalmente a las condiciones ambientales específicas del sitio como la disponibilidad de alimento y agua, así como la heterogeneidad y estructura del hábitat, las cuales en ocasiones permiten a las especies dividir los recursos disponibles y evitar la competencia (Gabor et al., 2001; Ilse y Hellgren, 1995a). Se ha documentado que una mayor disponibilidad de recursos alimentarios en hábitats heterogéneos permite el uso diferencial de los recursos, y por ende, la segregación espacio-temporal (Desbiez et al., 2009; Galetti et al., 2015; Keuroghlian et al., 2009), mientras que hábitats menos heterogéneos y con limitada disponibilidad de recursos conducen a un mayor grado de competencia (Gabor et al., 2001; Ilse y Hellgren, 1995b).
Los cerdos asilvestrados (Sus scrofa) están considerados entre las especies exóticas más dañinas a nivel mundial (Lowe et al., 2000). Su alta plasticidad ecológica les ha permitido adaptarse a nuevos ecosistemas y proliferar en grandes poblaciones alrededor del mundo, originando impactos ambientales como degradación del hábitat, destrucción de cultivos y depredación (Lowe et al., 2000). Se ha especulado que características particulares como sus hábitos generalistas, su capacidad de búsqueda de recursos y su capacidad para explotar recursos alimentarios como proteína animal e incluso semillas grandes que no pueden ser aprovechadas por otras especies, les confieren ventajas sobre los pecaríes (Desbiez y Keuroghlian, 2009). Como consecuencia, se predice que la presencia de esta especie exótica en lugares con poblaciones silvestres de pecaríes puede conducir a la competencia o a la división de nicho (Sicuro y Oliveira, 2002). La división de nicho es definida como el uso diferencial de los recursos alimentarios, del espacio y del tiempo por diferentes organismos o especies, que pueden explicar cómo las especies con traslape de requerimientos ecológicos consiguen coexistir (Desbiez et al., 2009).
La coexistencia de pecaríes de collar (Pecari tajacu) y cerdos asilvestrados ha sido registrada en la región de la laguna de Términos, al suroeste del estado de Campeche, desde hace más de 20 años (Góngora et al., 2011; Hidalgo-Mihart et al., 2014, 2017). Esta región del sur de México presenta una constante presión antrópica debido a la expansión de la frontera agropecuaria y al incremento de los cultivos de palma de aceite (Elaeis guineensis), los cuales ponen en peligro los últimos remanentes de vegetación conservada (Rivera-Arriaga y Villalobos-Zapata, 2005). La presencia de los cerdos asilvestrados en la región de la laguna de Términos podría generar impactos sobre las poblaciones nativas de pecaríes con las cuales interaccionan debido a la competencia por recursos. Esta problemática puede agravarse a causa de las presiones antrópicas a las que se enfrenta esta región, favoreciendo a los cerdos asilvestrados que tienden a aprovechar zonas abiertas, como cultivos y áreas ganaderas (Keiter y Beasley, 2017), a diferencia de los pecaríes de collar que tienden a utilizar fragmentos de vegetación con mayor cobertura forestal (Beca et al., 2017). El uso y selección de hábitat son de los principales mecanismos que se ven modificados debido a la competencia por recursos. Sin embargo, existen otros mecanismos como el patrón de actividad de las especies que puede ser modificado en función de la competencia (Galetti et al., 2015; Herrera et al., 2018).
El presente trabajo tuvo como objetivo evaluar el uso y selección de hábitat de pecaríes de collar y cerdos asilvestrados, e identificar las variables antrópicas y ecológicas que determinan la presencia de ambas especies en la región de la laguna de Términos, Campeche; así como analizar el patrón de actividad de pecaríes de collar en presencia y ausencia de cerdos asilvestrados. Particularmente, se responden las siguientes preguntas: ¿existe diferencia en el uso y selección de hábitat entre pecaríes de collar y cerdos asilvestrados?, ¿cuáles son las variables antrópicas y ecológicas que influyen en la presencia de ambas especies?, ¿la presencia de los cerdos asilvestrados modifica los patrones de actividad de los pecaríes de collar?
Materiales y métodos
El área de la laguna de Términos se localiza en la región occidental de Campeche, comprende el Área de Protección de Flora y Fauna Laguna de Términos y su zona de influencia (19°03’, 17°24’ N, -92°69, -90°16’ O; Fig. 1). El relieve es variable, formado en su mayoría por planicies inundables que se ubican a nivel del mar y algunos lomeríos que alcanzan los 100 m de altura. Las planicies inundables se encuentran sujetas a regímenes de inundación anual que pueden llegar a durar hasta 8 meses. El clima de la región es cálido subhúmedo con una temperatura promedio anual de 27 °C. La precipitación puede alcanzar los 2,000 mm al año (INEGI, 2016).
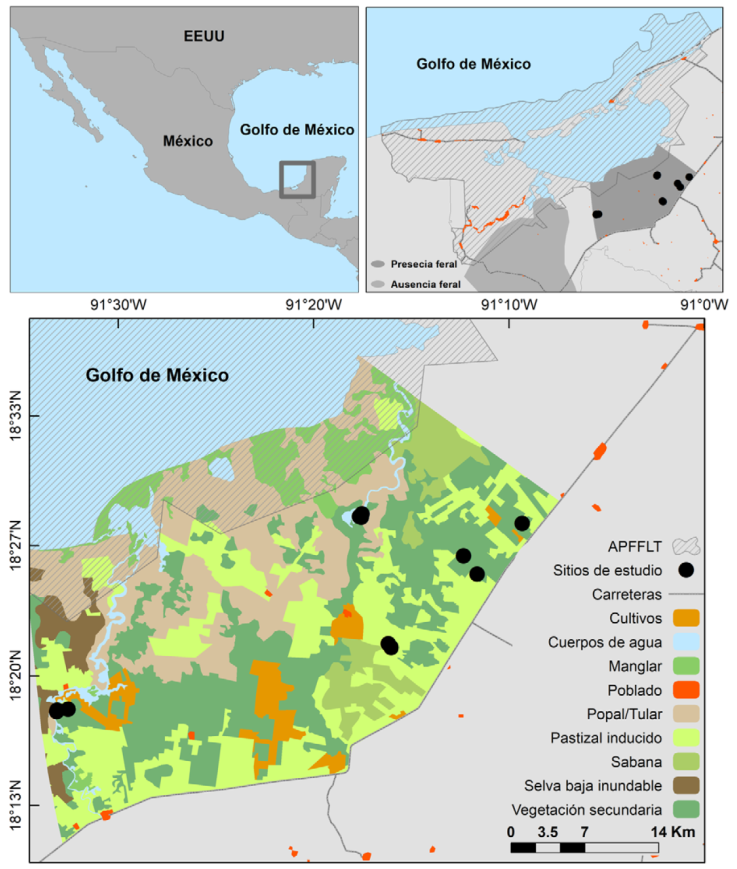
Figura 1 Localización geográfica del área de la laguna de Términos, Campeche, México. En el polígono “Presencia” se ha detectado la presencia de las 2 especies, mientras que en el polígono “Ausencia” solo se ha registrado la presencia de pecaríes de collar.
La tenencia de la tierra está bajo un régimen comunal y privado cuyas tierras son destinadas a la ganadería y al cultivo de palma de aceite (INEGI, 2016; Sánchez-Ramayo, 2003). La vegetación está compuesta por un mosaico de asociaciones vegetales entre las que se encuentran la selva mediana subcaducifolia, selva baja inundable, manglar, vegetación de pantano y vegetación secundaria (Miranda y Hernández, 1951). Una de las mayores problemáticas que enfrenta la zona es la rápida transformación de la vegetación natural a pastizales destinados a ganadería y a cultivos de palma de aceite. Se estima una pérdida del 43% de vegetación original, de la que la mayor parte corresponde a bosques tropicales (Soto-Galera et al., 2010).
Para la colecta de datos, se eligió un polígono de aproximadamente 1,265 km2 donde los cerdos asilvestrados han sido frecuentemente registrados y se establecieron 16 transectos de longitud variable (
Actualmente existe cierta controversia en el uso del término hábitat, el cual según Morrison et al. (2006) se refiere a los recursos y condiciones en un área que afectan la ocupación, supervivencia y reproducción de una especie. Sin embargo, frecuentemente se ha utilizado el término hábitat como sinónimo de tipo de vegetación. En este trabajo, selección de hábitat es definido como el uso diferencial de un tipo de asociación vegetal por las especies objetivo (Morrison et al., 2006). Para conocer la disponibilidad del hábitat, se registró el tipo de vegetación en los transectos cada 100 m y cada vez que se encontró rastro de la presencia de alguna de las especies objetivo. Los diferentes tipos de hábitats se clasificaron de acuerdo con Miranda y Hernández (1951) como: 1) vegetación secundaria (acahuales), 2) pastizal inducido (potreros), 3) popal/tular, 4) manglar, 5) sabana, 6) cultivos (palma africana) y 7) selva baja inundable (tintales). La abundancia de cerdos asilvestrados puede tener efectos negativos en la presencia de pecaríes de collar (Ilse y Hellgren, 1995b), por lo que se empleó la abundancia de cerdos asilvestrados por transecto como variable ecológica que afecta la presencia de pecaríes de collar y viceversa. Se empleó la abundancia de pecaríes de collar en cada transecto como variable ecológica que afecta la presencia de cerdos asilvestrados. Esta variable se calculó como el número de rastros de la especie entre la longitud del transecto (rastros/km).
Las variables de vegetación, antrópicas y físicas fueron obtenidas mediante cartografía digital con ayuda de ArcGIS (ESRI, 2013). Las variables relacionadas a la vegetación como la cobertura arbórea (Beca et al., 2017) y el porcentaje de popal/tular/palma de aceite afectan la presencia de ambas especies (Hernández-Pérez, 2019), por lo que fueron empleadas como variables predictoras. La información de estas variables fue generada creando un buffer con un radio de 1,800 m alrededor del punto medio de cada transecto y se obtuvo el porcentaje de cobertura arbórea y el porcentaje de popal/tular dentro de cada buffer (Hansen et al., 2013). El área del buffer (n = 10 km2) fue seleccionada debido a que abarca el ámbito hogareño de ambas especies, los cuales oscilan entre 2 y 7 km2 para pecaríes de collar, y entre 6 y 10 km2 para cerdos asilvestrados (Galetti et al., 2015; Ilse y Hellgren, 1995b). La densidad humana y distancia a asentamientos humanos son variables antrópicas que pueden determinar la presencia de las especies (García-Marmolejo et al., 2015), por lo que fueron empleadas como predictoras de la actividad humana en la región (Contreras-Moreno et al., 2016). Para la variable de densidad humana se creó un buffer con un radio de 1,800 m alrededor del punto medio de cada transecto y se obtuvo el número de habitantes dentro de cada buffer, mientras que la variable distancia a asentamientos humanos fue generada midiendo la distancia mínima entre el punto medio de cada transecto y el borde del asentamiento humano más cercano (INEGI, 2010). La distancia mínima a cuerpos de agua permanentes como los ríos, juega un papel importante en la dinámica de distribución y movimiento de ambas especies objetivo (Gabor et al., 2001; Keuroghlian et al., 2009), por lo que fue empleada como una variable física para determinar la presencia de ambas especies. Esta variable fue generada midiendo la distancia mínima entre el punto medio de cada transecto y el río más cercano (Maderey y Torres-Ruata, 1992).
Para evaluar los patrones de actividad de los pecaríes de collar se obtuvieron registros de la especie en sitios con presencia y ausencia de cerdos asilvestrados. Los sitios con presencia de cerdos asilvestrados fueron zonas donde se obtuvieron registros de pecaríes de collar y cerdos asilvestrados en los últimos 3 años (Hidalgo-Mihart et al., 2017). En estos sitios se empleó una red de 11 estaciones de fototrampeo y 1,428 días trampa entre enero y junio de 2018. La distancia mínima entre 2 cámaras fue de 0.5 km y la máxima de 8 km. Los sitios con ausencia de cerdos asilvestrados fueron sitios donde no se ha registrado la presencia de la especie exótica o se ha registrado únicamente de manera esporádica, es decir 1 o 2 registros en los últimos 3 años. En estos sitios se empleó una red de 230 estaciones de fototrampeo con una distancia mínima de 2 km y un resultado de 54,473 días trampa efectuado por personal de la Universidad Juárez Autónoma de Tabasco (Hidalgo-Mihart et al., 2017) entre enero de 2015 y diciembre de 2017 (Fig. 1). La distancia mínima entre los sitios con presencia y ausencia de cerdos asilvestrados fue de aproximadamente 15 km. Los registros únicos o independientes fueron fotos de una especie separados por al menos 60 minutos.
Para evaluar el uso de hábitat se aplicó el índice de Levin estandarizado con ayuda del software Havistat 2.0 (Montenegro et al., 2014) independientemente para las 2 especies bajo estudio. Este índice toma valores entre 0 y 1 indicado que una especie usa una amplia variedad de hábitats cuando se obtiene un valor mayor a 0.6 (Krebs, 1999). Para estimar el grado de traslape en el uso de hábitat, se empleó el índice de Pianka mediante el paquete EcoSimR en R (Gotelli et al., 2015). El índice de Pianka varía entre 0 y 1, un valor igual a 1 indica traslape completo (Putman, 1994).
La disponibilidad de hábitats registrada en los transectos se contrastó con la proporción de tipos de vegetación donde se obtuvieron rastros de cada especie a través de un análisis de x 2 . Posteriormente, se emplearon intervalos de Bonferroni para evaluar la selección o evasión de hábitats por especie (Byers et al., 1984). Estos análisis se realizaron empleando el software Havistat 2.0 (Montenegro et al., 2014). Se verificó la concordancia entre la disponibilidad de los tipos de hábitats observados en campo con los plasmados en el mapa de vegetación y uso de suelo más actualizado del Instituto Nacional de Estadística y Geografía (INEGI, 2014) mediante el coeficiente de Kappa.
Para determinar las variables antrópicas y ecológicas que influyen en la presencia de cada especie, se emplearon modelos lineales generalizados con distribución binomial. Se realizó una regresión de pasos hacia atrás para aprovechar la mayor variabilidad del proceso de análisis a partir de un modelo completo (Crawley, 2007). Las 6 variables predictoras empleadas en los modelos fueron seleccionadas de acuerdo con criterios ecológicos relevantes para cada especie; posteriormente se realizaron pruebas de correlación de Pearson (r < 0.7) para identificar variables correlacionadas. Estos análisis fueron realizados empleando el paquete stats en R.
Los mejores modelos se seleccionaron con base en el criterio de información de Akaike corregido para tamaños de muestra pequeños (AICc), el valor de R2 y el valor de p obtenidos mediante la función aictab del paquete AICcmodavg en R (Mazerolle, 2017). Las diferencias de Akaike (∆AIC) y el peso de Akaike (AICw) se utilizaron para clasificar los modelos de mayor a menor con el respaldo de los datos. Los valores de ∆AIC > 2 se usaron para discriminar el ajuste de cada modelo (Burnham y Anderson, 1998).
El patrón de actividad fue evaluado para ambas especies mediante estadística circular con el software Oriana 4.0 (Kovach Computing Service, 2011). Para conocer el grado de traslape en los patrones de actividad de los pecaríes de collar en presencia y ausencia de cerdos asilvestrados, se aplicó un análisis de densidad de Kernel y un análisis de Wald para comparar las diferencias estadísticas entre las horas de mayor actividad, así como un análisis de Mann-Whitney para comparar diferencias entre el patrón de actividad de pecaríes en ausencia y presencia de cerdos asilvestrados. Los análisis se realizaron con los paquetes estadísticos Overlap versión 3.5.1 y Activity versión 3.5.1 en R (Ridout y Linkie, 2009; Rowcliffe, 2016).
Resultados
Se registraron 7 tipos de hábitats en 239 puntos a lo largo de los transectos recorridos, de los cuales en 50, se registraron rastros de pecaríes de collar y en 177, rastros de cerdos asilvestrados. La vegetación secundaria fue el tipo de hábitat más común en los transectos con 58.1%, seguido por pastizales inducidos con 29%, selva baja inundable con 3.8%, sabanas con 3.8%, popal/tular con 3.5%, cultivos de palma de aceite con 1.5% y manglar con 0.3% (Tabla 1). El coeficiente de Kappa mostró un 60% de concordancia entre la disponibilidad de hábitats observada en los transectos y la plasmada en el mapa más actual para el polígono de estudio (INEGI, 2014).
Tabla 1 Selección de hábitat de pecaríes de collar (Pecari tajacu) y cerdos asilvestrados (Sus scrofa) en la región de la laguna de Términos, Campeche, México de acuerdo con el método de Byers et al. (1984).
| Uso de suelo y vegetación | Área (km2)* | Pecari tajacu | Sus scrofa | ||
|---|---|---|---|---|---|
| Frecuencia observada | Frecuencia esperada | Frecuencia observada | Frecuencia esperada | ||
| Vegetación secundaria (acahuales) | 384.40 | 21 (=) | 21.45 | 39 (-) | 74.04 |
| Pastizales inducidos (potreros) | 330.07 | 3 (-) | 17.98 | 48 (=) | 62.06 |
| Tular/Popal | 275.99 | 0 (-) | 2.17 | 26 (+) | 7.49 |
| Manglar | 92.60 | 0 (-) | 0.19 | 1 (=) | 0.64 |
| Sabana | 84.81 | 7 (=) | 2.36 | 11 (=) | 8.13 |
| Cultivos (palma africana) | 23.61 | 0 (-) | 0.93 | 14 (+) | 3.21 |
| Selva baja inundable (tintales) | 18.58 | 31 (+) | 16.93 | 75 (=) | 58.42 |
+, -, = indican si el tipo de vegetación fue seleccionado más, menos o igual de acuerdo con la disponibilidad registrada en los transectos.
* Con base en datos de INEGI (2014).
Los pecaríes de collar usaron 4 de los 7 tipos de hábitat, a diferencia de los cerdos asilvestrados, quienes usaron todos los tipos (Fig. 2). La amplitud de uso de hábitat para pecaríes fue menor (B = 0.30, n = 4, p < 0.01) con respecto a los cerdos asilvestrados (B = 0.60, n = 7, p < 0.01). De acuerdo con el índice de amplitud de Levin, los pecaríes de collar presentaron una mayor especificidad en el uso de hábitats respecto a los cerdos asilvestrados.
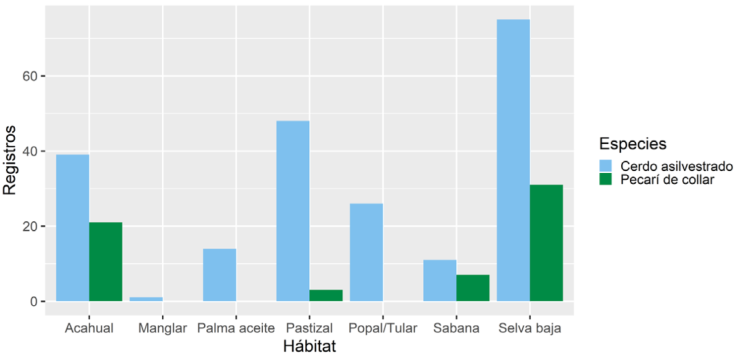
Figura 2 Tipos de vegetación y frecuencia de registros de pecaríes de collar (Pecari tajacu) y cerdos asilvestrados (Sus scrofa) en la región de la laguna de Términos, Campeche, México.
Se obtuvo un alto grado de traslape en el uso de hábitat entre ambas especies (IP = 0.86, p = 0.034). Los cerdos asilvestrados mostraron un mayor uso de áreas abiertas como potreros y cultivos, mientras que los pecaríes de collar utilizaron mayormente las áreas forestadas como la selva baja inundable. Ambas especies usaron la vegetación secundaria, pastizales inducidos, selva baja inundable y sabanas.
La selva baja inundable fue la vegetación en la que se registró la mayor cantidad de rastros y avistamientos de ambas especies. Para los pecaríes de collar se obtuvieron más registros de lo esperado en la selva baja inundable, mostrando una selección por este tipo de hábitat (n = 62, x 2 = 41.1, p < 0.01), y se obtuvieron menos registros de lo esperado en los pastizales inducidos, popales/ tulares, cultivos de palma de aceite y manglar. Los cerdos asilvestrados fueron registrados más de lo esperado en los cultivos de palma de aceite y en los popales/tulares, (n = 214, x 2 = 76.2, p < 0.01) y menos de lo esperado en la vegetación secundaria (Tabla 1).
Los modelos con el mejor ajuste para los pecaríes de collar fueron el modelo mínimo que incluyó la variable: distancia a asentamientos humanos; y el modelo B que incluyó las variables: abundancia relativa de cerdos asilvestrados, distancia a asentamientos humanos y cobertura arbórea. Para cerdos asilvestrados los modelos con el mejor ajuste fueron el modelo mínimo que incluyó las variables: abundancia de pecaríes de collar, distancia a asentamientos humanos, distancia a ríos y cobertura arbórea; y el modelo A que incluyó las variables: abundancia de pecaríes de collar, distancia a asentamientos humanos, distancia a ríos, cobertura arbórea y densidad humana (Tabla 2).
Tabla 2 Modelos lineales generalizados para evaluar el efecto de variables ambientales en la presencia de pecaríes de collar (Pecari tajacu) y cerdos asilvestrados (Sus scrofa) en la región de la laguna de Términos, Campeche, México. Los modelos con ∆AIC < 2 se consideraron los de mejor ajuste.
| Especie | Modelo | Variables | K | R2 | ∆AIC | AICc | AICw |
|---|---|---|---|---|---|---|---|
| Pecari tajacu | Mínimo | Dpo | 2 | 0.21 | 0 | 63.84 | 0.56 |
| Modelo B | As, Dpo, Ca | 4 | 0.26 | 1.03 | 64.86 | 0.33 | |
| Modelo A | As, Dpo, Dr, Ca | 5 | 0.26 | 3.41 | 67.25 | 0.10 | |
| Completo | As, Dpo, Dr, Ca, Dh, Pop | 7 | 0.27 | 8.36 | 72.19 | 0.01 | |
| Nulo | SV | 1 | 14.98 | 77.91 | 0 | ||
| Sus scrofa | Mínimo | Ap, Dpo, Dr, Ca | 5 | 0.51 | 0 | 48.91 | 0.56 |
| Modelo A | Ap, Dpo, Dr, Ca, Dh | 6 | 0.53 | 1.02 | 49.92 | 0.34 | |
| Completo | Ap, Dpo, Dr, Ca, Dh, Pop | 7 | 0.54 | 3.29 | 52.20 | 0.11 | |
| Nulo | SV | 1 | 30.8 | 79.71 | 0 |
Variables: Ap = abundancia Pecari tajacu, As = abundancia Sus scrofa, Ca = % de cobertura arbórea dentro de un buffer con un radio de 1,800 m alrededor de cada transecto, Dh = densidad humana dentro de un buffer con un radio de 1,800 m alrededor de cada transecto, Dpo = distancia mínima a asentamientos humanos, Dr = distancia mínima a ríos, Pop = % de cultivos de palma de aceite y popales/tulares dentro de un buffer con un radio de 1,800 m alrededor de cada transecto, SV = sin variables.
Las estimaciones de los parámetros sugieren que la probabilidad de encontrar pecaríes de collar aumentó conforme se incrementó la distancia a los asentamientos humanos (Fig. 3). Por su parte, la probabilidad de encontrar cerdos asilvestrados aumentó ligeramente con la disminución de la distancia a asentamientos humanos, la disminución del porcentaje de cobertura arbórea y al incrementar la distancia a los ríos (Tabla 3, Fig. 4).
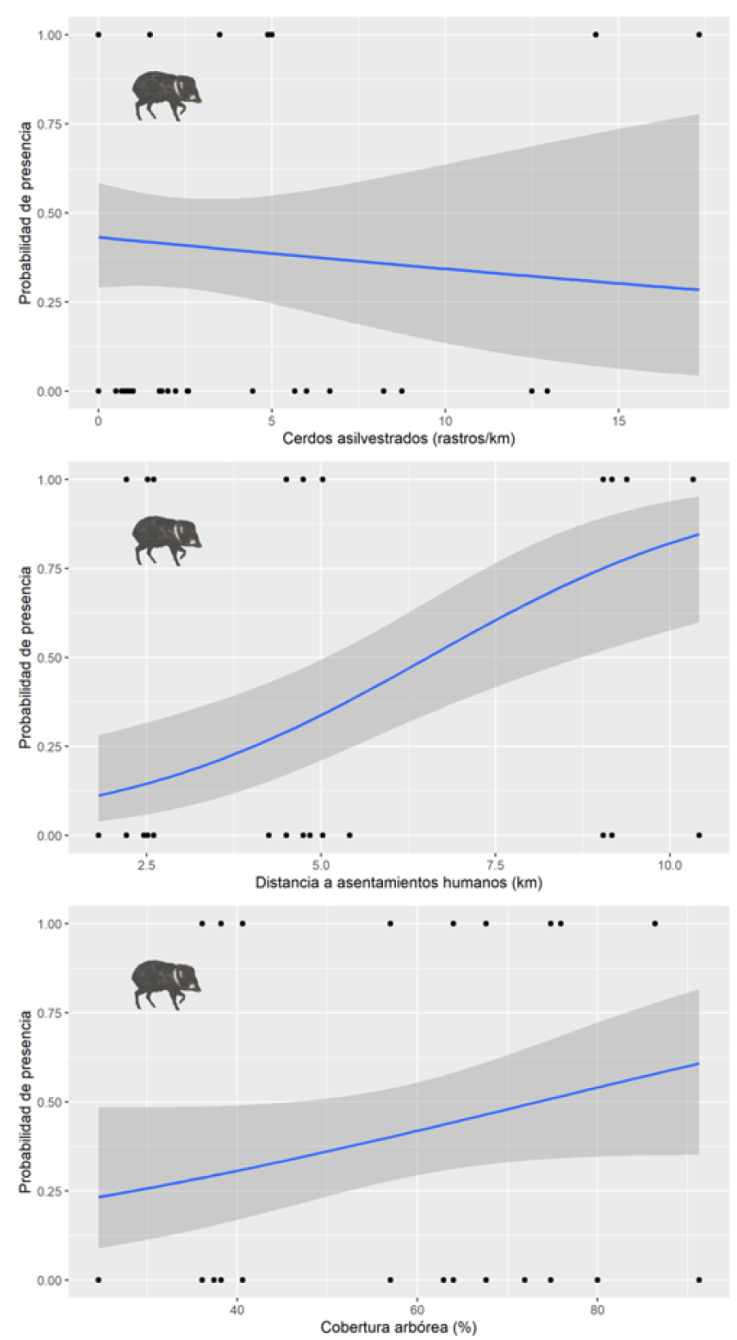
Figura 3 Variables antrópicas y ecológicas que influyen en la presencia de los pecaríes de collar (Pecari tajacu) en la región de la laguna de Términos, Campeche, México.
Tabla 3 Estimaciones, error estándar y valor de p de parámetros de los mejores modelos para pecaríes de collar (Pecari tajacu) y cerdos asilvestrados (Sus scrofa) en la región de la laguna de Términos, Campeche, México.
| Modelo | Variables | Pecari tajacu | p | Sus scrofa | p |
|---|---|---|---|---|---|
| Mínimo | Intercepto | -2.8698 (± 0.77) | 0.01 | 8.25697 (±3 .26) | 0.01 |
| Distancia mínima a asentamientos humanos | 0.4395 (± 0.12) | 0.01 | -1.25018 (± 0.64) | 0.05 | |
| Abundancia Pecari tajacu | -0.45493 (± 0.46) | 0.32 | |||
| Distancia mínima a ríos | 0.90526 (± 0.42) | 0.03 | |||
| Porcentaje de cobertura arbórea | -0.08912 (± 0.03) | 0.01 | |||
| B y A | Intercepto | -4.6603 (± 1.37) | 0.01 | 7.36751 (± 3.07) | 0.01 |
| Abundancia Sus scrofa | 0.13900 (± 0.08) | 0.11 | |||
| Distancia mínima a asentamientos humanos | 0.52775 (± 0.15) | 0.01 | -1.16660 (± 0.59) | 0.04 | |
| Porcentaje de cobertura arbórea | 0.01620 (± 0.02) | 0.30 | -0.11025 (± 0.04) | 0.01 | |
| Abundancia Pecari tajacu | -0.44733 (± 0.46) | 0.33 | |||
| Distancia mínima a ríos | 1.20586 (± 0.54) | 0.02 | |||
| Densidad humana | 0.19036 (± 0.16) | 0.24 |
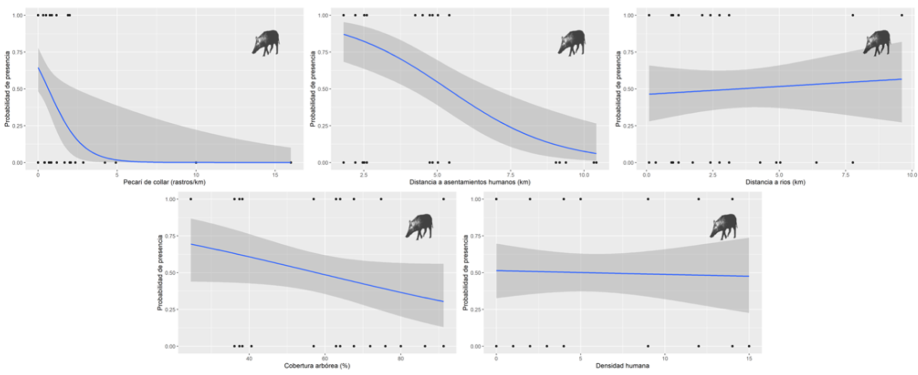
Figura 4 Variables antrópicas y ecológicas que influyen en la presencia de los cerdos asilvestrados (Sus scrofa) en la región de la laguna de Términos, Campeche, México.
Se detectaron diferencias estadísticamente significativas en los periodos de actividad de los pecaríes de collar en presencia y ausencia de los cerdos asilvestrados (U = 13.632, p < 0.05). En ausencia de cerdos asilvestrados, los pecaríes mostraron un patrón de actividad catemeral (Fig. 5a). Se observó una mayor actividad en la fase diurna con un mayor pico de actividad alrededor de las 10:03 h (n = 867, Z = 60.43, p < 0.01), en comparación con la fase crepuscular-nocturna (W = 32.39, p < 0.01; Fig. 5a). En presencia de cerdos asilvestrados, la actividad de los pecaríes se volvió principalmente diurna con un pico de actividad alrededor de las 08:45 h y con pocos registros nocturnos (n = 88, Z = 14.19, p < 0.01; Fig. 5b). Los cerdos asilvestrados mostraron un patrón de actividad nocturno, siendo más activos durante el periodo de las 00:00 a 06:00 h (n = 202, W = 7.56, p = 0.005) con un pico de mayor actividad alrededor de las 00:10 h (Z = 102.19, p < 0.01; Fig. 5c). Se estimó un mayor coeficiente de traslape de los patrones de actividad de pecaríes de collar en sitios con ausencia de cerdos asilvestrados (∆ = 0.37; Fig. 6a), en comparación con sitios donde se registraron ambas especies (∆ = 0.28; Fig. 6b).
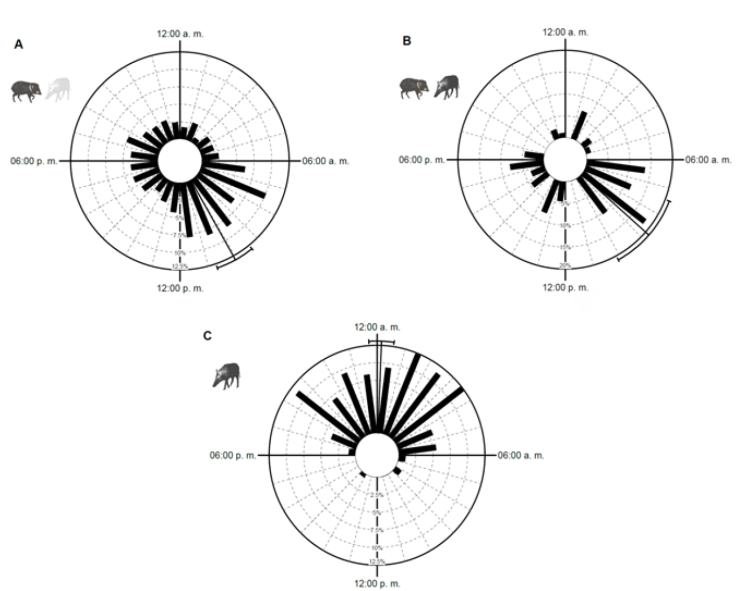
Figura 5 Patrones de actividad de: A) pecaríes de collar (Pecari tajacu) en ausencia de cerdos asilvestrados (Sus scrofa); B) pecaríes de collar en presencia de cerdos asilvestrados; C) cerdos asilvestrados en la región de la laguna de Términos, Campeche, México. Las barras muestran el porcentaje de registros independientes. El vector representa el intervalo de confianza de 95%.
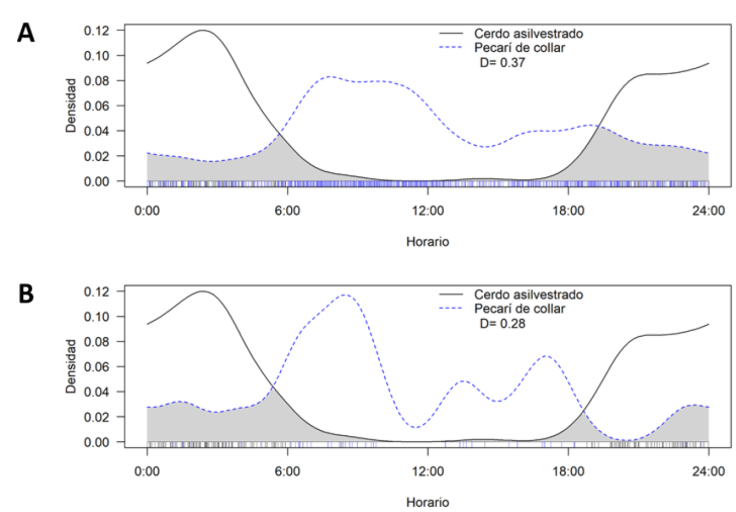
Figura 6 Traslape del patrón de actividad diaria del pecarí de collar (Pecari tajacu): A) en sitios con ausencia y B) en sitios con presencia de cerdos asilvestrados (Sus scrofa) en la región de la laguna de Términos, Campeche, México. El coeficiente de superposición a 95% es el área bajo la curva de ambas estimaciones de densidad (indicadas en color gris).
Discusión
La introducción de una especie exótica en sitios con especies ecológicamente similares puede conducir al traslape de nicho y competencia; o en dado caso a la división de recursos que permita su coexistencia (Sanders et al., 2003). Nuestros resultados reportan la división de recursos espacio-temporales entre pecaríes de collar y cerdos asilvestrados en la región de la laguna de Términos, lo cual puede estar explicando parcialmente la coexistencia entre ambas especies en el sur de México.
El grado de traslape en el uso de hábitat encontrado en el presente trabajo podría deberse a la capacidad y flexibilidad de los cerdos asilvestrados para explotar los recursos disponibles (Ilse y Hellgren, 1995a), ya sea hábitats con actividades antrópicas como pastizales inducidos o cultivos de palma de aceite. Los pecaríes de collar, por su parte hacen un mayor uso de áreas más boscosas (Beca et al., 2017; Keuroghlian et al., 2009); como se documentó en este trabajo donde la especie únicamente usó áreas con mayor cobertura arbórea y alejadas de asentamientos humanos como la selva baja inundable y vegetación secundaria.
Los cerdos asilvestrados, especie generalista, podrían tener ventajas y competir con los pecaríes de collar que son más selectivos en el uso de hábitat (Beca et al., 2017; Sicuro y Oliveira, 2002). El comportamiento selectivo en los pecaríes de collar, en específico para la región de la laguna de Términos, podría ser respuesta a la protección contra depredadores, incluido el ser humano, además podría ser una cualidad para adaptarse a presiones como la presencia de los cerdos asilvestrados (Gabor et al., 2001; Reyna-Hurtado y Tanner, 2005). Se ha documentado que un depredador tope como el jaguar (Panthera onca) tiene mayor probabilidad de cazar un pecarí de collar, debido a que los pecaríes son más pequeños y menos agresivos a diferencia de los cerdos asilvestrados (Da Rosa et al., 2016). Por otra parte, como sugieren nuestros datos, la presencia de pecaríes de collar es mayor en sitios alejados de asentamientos humanos, posiblemente por 2 razones: 1) estos sitios son los últimos remanentes de vegetación conservada, 2) son sitios menos concurridos por ganaderos y otras personas.
Los análisis sobre selección de hábitat muestran una segregación espacial entre las 2 especies. Estos datos son consistentes con la hipótesis presentada por Ilse y Hellgren (1995a) para zonas áridas, donde los cerdos asilvestrados y pecaríes de collar seleccionan diferentes tipos de hábitats basados en la cobertura arbórea y disponibilidad de recursos alimenticios. Así también coincide con lo encontrado para regiones tropicales, donde los pecaríes son más selectivos a escala de microhábitat, seleccionando áreas con mayor cobertura arbórea y con vegetación de sotobosque (Beca et al., 2017), a diferencia de los cerdos asilvestrados que son más aptos para usar áreas abiertas e inundables que no son explotadas por los pecaríes (Keuroghlian et al., 2009).
Los análisis aquí presentados indican una mayor selección de popales/tulares y cultivos de palma de aceite por parte de los cerdos asilvestrados. Las áreas abiertas e inundables como los popales/tulares podrían estar siendo seleccionadas como áreas de forrajeo debido a la presencia de herbáceas como el platanillo de agua (Thalia geniculata). A su vez, son áreas que se inundan temporalmente y que permiten la termorregulación de la especie (Gabor et al., 2001). En Sudamérica, la palma Attalea phalerata, especie similar a la palma de aceite, pero originaria de la Amazonía, es común en la dieta de los cerdos asilvestrados (Desbiez et al., 2009). Durante el transcurso de este estudio se colectaron contenidos estomacales de cerdos asilvestrados (datos no publicados) en los que se documentó la presencia de una gran cantidad de frutos de palma de aceite. Aparentemente, las plantaciones de palma podrían estar jugando un papel importante en la presencia y abundancia de los cerdos asilvestrados en la región, dotando de una abundante fuente de recursos disponibles a la especie exótica.
El impacto de las actividades antrópicas es a menudo muy complejo, ocasionando modificaciones en el paisaje, así como en las relaciones ecológicas y comportamiento de muchas especies (Ohashi et al., 2013). La heterogeneidad de hábitat en la región de la laguna de Términos, a causa de actividades antrópicas, está permitiendo que la competencia entre pecaríes de collar y los cerdos asilvestrados sea mínima, debido a la presencia de zonas de cultivo y ganaderas explotadas por los cerdos asilvestrados, así como áreas conservadas en las que la presencia de pecaríes de collar es mayor.
Se ha considerado al pecarí de collar como una especie flexible que puede habitar tanto áreas impactadas por actividades humanas, así como áreas en buen estado de conservación (Altrichter y Boaglio, 2004; Reyna-Hurtado y Tanner, 2005). No obstante, los datos presentados en este estudio indican que la presencia de los pecaríes de collar está correlacionada positivamente con la distancia a asentamientos humanos, puesto que las mayores frecuencias de rastros y avistamientos de esta especie se obtuvieron en fragmentos de selva alejados de los asentamientos humanos. Estos datos son consistentes con lo documentado por otros autores, quienes indican que la presencia y abundancia de pecaríes de collar se ven negativamente afectadas cuando enfrentan presiones antrópicas como cacería, disminución de cobertura arbórea, densidad y proximidad a asentamientos humanos (Beca et al., 2017; Briceño-Méndez et al., 2016).
La probabilidad de presencia de los cerdos asilvestrados está correlacionada negativamente con la distancia a los asentamientos humanos y con el porcentaje de cobertura arbórea. Esto podría deberse a que muchos de los cultivos de palma de aceite y pastizales cultivados se encuentran cercanos a las comunidades humanas, siendo los cultivos de palma de aceite uno de los hábitats seleccionados por la especie, y los pastizales cultivados sitios con baja cobertura arbórea. Estas variables antrópicas afectan de manera positiva la existencia de la especie como lo demuestran otros estudios en áreas impactadas por actividades humanas en Norteamérica (Keiter y Beasley, 2017; McClure et al., 2015). A pesar de no ser estadísticamente significativa, existe una correlación positiva entre la presencia de los cerdos asilvestrados y la distancia a los ríos. Dicha correlación podría deberse a que los ríos en la región son los principales medios de comunicación entre comunidades humanas y entre sitios de caza. Posiblemente los cerdos asilvestrados evitan la cercanía a ríos debido a que son más propensos a ser cazados si son observados desde botes o canoas a diferencia de sitios alejados, que representa más esfuerzo para el cazador, puesto que necesita invertir más tiempo en la búsqueda de su presa. En cuanto a disponibilidad de agua, los ríos no son la única fuente. La región de la laguna de Términos es un humedal que se encuentra sujeto a regímenes de inundación anual que pueden llegar a durar hasta 8 meses, además de esto la gran cantidad de bebederos artificiales (jagüeyes) creados por los ganaderos para suministrar de agua al ganado doméstico en época de estiaje. Estas características podrían beneficiar a los cerdos asilvestrados, los cuales hacen uso de áreas impactadas como potreros donde se ubican dichos jagüeyes, así como otras zonas inundables que les proveen de refugio y una gran variedad de recursos alimentarios (Rosell et al., 2004).
Muchos organismos tienden a modificar sus patrones de actividad para reducir o evitar la competencia por recursos con otras especies. Tal es el caso de los pecaríes de collar, los cuales se ha comprobado que modifican sus patrones de actividad con relación a la presencia de los cerdos asilvestrados (Galetti et al., 2015). Los análisis en este estudio reafirman este comportamiento. Incluso al comparar el patrón de actividad de pecaríes de collar en sitios con ausencia y presencia de la especie exótica, se observó un mayor traslape en sitios con ausencia indicando una posible modificación de los patrones de actividad de los pecaríes de collar en áreas donde coexisten con los cerdos asilvestrados. Existen otras variables no tomadas en cuenta en este estudio, las cuales podrían estar afectando la dinámica de los patrones de actividad de ambas especies. Tal es el caso de la cacería y persecución con perros, actividad que es muy común en el sur de México.
Es posible que la heterogeneidad del paisaje en la región de la laguna de Términos, donde la composición de áreas altamente impactadas por el ser humano ligadas a fragmentos aún conservados está permitiendo la división de nicho y por consiguiente la segregación espacio-temporal entre ambas especies. Sin embargo, si las influencias antrópicas como el establecimiento de cultivos de palma de aceite se incrementa, podrían permitir la expansión de la distribución de la especie exótica hacia otros sitios. Por su parte, la fragmentación del hábitat y disminución en la cobertura arbórea puede desequilibrar aún más el proceso de coexistencia, y al no haber hábitats idóneos para los pecaríes, la disputa por los recursos podría derivar en una mayor competencia y favorecer el incremento de la población de cerdos asilvestrados en la región.

