











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La depredación de semillas es un fenómeno que incide en la regeneración y la dinámica poblacional de las especies vegetales, y puede ocurrir antes o después de que las semillas sean dispersadas (Kolb et al., 2007). El consumo de semillas que se realiza antes de la dispersión se conoce como depredación predispersión y comúnmente se lleva a cabo por diversas especies de invertebrados, mientras que los mamíferos son los consumidores más importantes en la depredación post-dispersión (Crawley, 2000).
Los escarabajos de la familia Curculionidae suelen ser los depredadores predispersión más comunes de semillas en el género Quercus (Johnson et al., 2009). Sin embargo, también es común la depredación por larvas de lepidópteros del género Cydia (Tortricidae) (Branco et al., 2002; Soria, Cano et al., 1999). Diversos estudios han analizado la depredación que llevan a cabo los curculiónidos y su relación con el tamaño de las bellotas, así como sus posibles efectos en la germinación y la regeneración de las poblaciones de encinos (Espelta, Bonal et al., 2009; Fukumoto y Kajimura, 2000; Lombardo y McCarthy, 2009; Yi y Zhang, 2008). Sin embargo, la depredación por larvas de lepidópteros ha recibido mucha menos atención (Soria et al., 2002).
En general, las larvas de curculiónidos suelen ser más abundantes que las de Cydia (Bonal et al., 2007; Fukumoto y Kajimura, 2000; Soria, Cano et al., 1999; Soria, Villagrán et al., 1999), aunque no siempre es el caso (Branco et al., 2002; Díaz-Fleischer et al., 2010). La presencia de más de una larva por bellota, fenómeno conocido como superinfestación, es relativamente común en los curculiónidos, pero es un evento muy improbable en el caso de las palomillas del género Cydia (Bonal y Muñoz, 2008, 2009; Soria, Villagrán et al., 1999).
El tamaño de las bellotas influye en la depredación predispersión realizada por curculiónidos. Algunos estudios han mostrado que los niveles de depredación en especies de Quercus que coexisten son mayores en la especie con bellotas más grandes que en la que tiene bellotas más pequeñas ( Espelta, Bonal et al., 2009; Espelta, Cortés et al., 2009). Se ha mostrado también que al interior de algunas especies de encinos las bellotas más grandes son más atacadas, lo que muestra que las hembras son capaces de seleccionar a las bellotas por su tamaño (Bonal et al., 2012; Díaz-Fleisher et al., 2010). Esto permite que las larvas encuentren el alimento suficiente para su desarrollo; incluso se ha documentado una relación positiva entre el tamaño de las larvas de Curculio y el tamaño promedio de las bellotas en Q. ilex (Bonal et al., 2012). En las palomillas del género Cydia no se ha reportado selección de las bellotas por su tamaño o por algún otro factor; sin embargo, se sabe que sus larvas tienen mayor movilidad que las de los curculiónidos (Jiménez et al., 2006), por lo que podrían alimentarse de más de una bellota.
En México se ha documentado la depredación predispersión en algunas especies de encinos, pero generalmente el énfasis se ha puesto en la evaluación de su magnitud y de las pérdidas que representa en términos de la capacidad de germinar de las semillas (Díaz-Fleischer et al., 2010; González-Salvatierra et al., 2013). En contadas ocasiones se identifica el daño causado por diferentes tipos de insectos, y no se han hecho comparaciones entre distintas especies de encinos que coexisten en una misma localidad, de forma que no es posible establecer si existen preferencias de distintos tipos de insectos por algunas especies de Quercus, ni el efecto que otros factores, como el tamaño de la bellota, la densidad del arbolado o la concentración de taninos en las bellotas, puedan tener en estas interacciones. Desafortunadamente, el conocimiento actual de las distintas especies de curculiónidos y palomillas que infestan a las bellotas de encinos mexicanos es muy limitado, y la única descripción disponible de algunas especies de Curculio se publicó en los años setenta (Gibson, 1977), por lo que persisten grandes lagunas en el conocimiento de la diversidad real de estos escarabajos. Tampoco hay información disponible sobre las especies de Cydia presentes en las distintas especies de encinos, ni sobre el nivel de especificidad de esta interacción.
En este contexto, el objetivo general de este trabajo es analizar la depredación predispersión realizada por escarabajos de la familia Curculionidae y por palomillas del género Cydia en 3 especies de encinos que coexisten en el sur de la cuenca de México, de la cual es parte el volcán Ajusco. En particular, nos planteamos: a) establecer si existen preferencias de los curculiónidos o las palomillas por alguna de las especies de estudio, b) en caso de haberla, analizar si está asociada con el tamaño de las bellotas de cada especie, c) analizar la relación entre la depredación por curculiónidos y el tamaño de la semilla al interior de cada especie, y d) evaluar la superinfestación por curculiónidos en las especies de estudio.
Aunque diversos trabajos han analizado la depredación realizada por curculiónidos, éste es, hasta donde sabemos, el primer análisis que incluye tanto curculiónidos como palomillas del género Cydia. Debido a que no es posible diferenciar especies de curculiónidos, es muy probable que el análisis incluya en un mismo grupo a más de una especie de esta familia de escarabajos. En el caso de las palomillas, es muy poco probable que haya más de una especie del género Cydia, pues en los trabajos publicados hasta ahora no se ha registrado más de una especie en un hospedero. Este estudio tiene además la particularidad de ampliar el análisis de la depredación predispersión al incluir a una especie de encino con bellotas muy pequeñas (Q. castanea), que coexiste con otros con bellotas más grandes; los estudios realizados hasta ahora incluyen especies cuyas bellotas se encuentran en un intervalo de tamaños de medio a grande.
Materiales y métodos
El estudio se llevó a cabo en el Centro de Educación Ambiental Ecoguardas (CEA), que tiene una superficie de 132.6 ha y se localiza en la parte media del volcán Ajusco, al sur de la cuenca de México (19°16’40.63”19°15’45.84” N, 99°12’23.088”- 99°11’19.23” O; PAOT, 2009). La altitud promedio de la zona de estudio es de 2,507 m snm. El clima es templado subhúmedo con lluvias en verano (Cb’ (wa)(w)(ig), con una temperatura media anual entre 12 y 18 °C y una precipitación anual de alrededor de 1,000 mm (Álvarez-Cruz, 1992). En el CEA se encuentra un remanente de la vegetación nativa del Pedregal de San Ángel y forma parte de las 5 zonas protegidas que se encuentran en el derrame del Xitle (Cano-Santana et al., 2006). La vegetación dominante consiste de bosque de encino y matorral xerófilo. En el bosque de encino dominan: Q. castanea, Q. rugosa, Q. obtusata, Q. crassipes y Q. laurina, y en el matorral xerófilo: Buddleja cordata, B. parviflora, Dodonaea viscosa y Pittocaulon praecox (PAOT, 2009; H. Díaz-Guzmán, obs. pers.).
Diversas especies de escarabajos de la familia Curculionidae depredan las bellotas de los encinos; entre ellos destacan los del género Curculio, presentes en los encinares del Ajusco (C. Bonfil, obs. pers.). Estos escarabajos tienen apéndices locomotores largos y un rostrum alargado, que les permite sujetar y perforar las bellotas para luego depositar sus huevos dentro (Bonal et al., 2011; Hughes y Vogler, 2004). Su ciclo de vida se relaciona temporalmente con el de los encinos, ya que los adultos consumen las hojas y las hembras ovipositan en las bellotas, en donde se desarrollan las larvas. En algunos casos más de una hembra oviposita en la misma bellota, fenómeno que se conoce como superinfestación (Bonal et al., 2009). En este trabajo se denominará a las larvas, sin distinguir géneros o especies, como curculiónidos.
Las palomillas del género Cydia (Tortricidae), en su etapa larvaria son consumidoras de semillas y frutos; las hembras adultas depositan los huevos sobre las bellotas o en las hojas y tallos cercanos a ellas, y cuando las larvas eclosionan se introducen en los frutos (Rohlfs, 1999; Soria et al., 2002). Generalmente no se encuentra más de una larva de palomilla en la misma bellota, pero se ha observado que una larva puede desplazarse de una bellota a otra si no encuentra suficiente alimento, aunque no es un fenómeno común (Jiménez et al., 2006). En este trabajo se usarán indistintamente los términos palomillas o Cydia al referirnos a estos organismos.
Las 3 especies de encinos estudiadas [Q. obtusata Humb. et Bonpl., Q. rugosa Née y Q. castanea Née (Fagaceae)] son comunes en los encinares de la cuenca de México y tienen una amplia distribución en el país (Romero-Rangel et al., 2015). Presentan una fructificación anual y los frutos maduran entre septiembre y noviembre (Romero-Rangel et al., 2015). Quercus obtusata tiene las bellotas más grandes (17-20 mm de largo, 15-20 mm ancho), seguidas por las de Q. rugosa (15-25 mm largo, 8-12 mm ancho) y, finalmente, las de Q. castanea son las más pequeñas (10-12 mm largo, 8-10 mm ancho; González-Villarreal, 1986).
Entre octubre y noviembre de 2015 se realizaron 8 recorridos en el CEA, durante los cuales se identificaron y marcaron 15 árboles (5 por especie), y se recolectaron 50 o más bellotas por árbol, las cuales se numeraron individualmente según el árbol del que procedían. Se midió el largo (D 1) y ancho (D 2) de cada bellota con ayuda de un vernier digital, para estimar su volumen con la fórmula:
donde r1 = D 1/2 y r2 = D 2/2.
Se colocó individualmente una bellota, con un número asignado, en una cavidad de charola para hielo, realizando revisiones diarias durante un mes, para registrar el número y tipo de larvas presentes en cada una. Transcurrido un mes se abrieron todas las bellotas, para registrar si quedaba alguna larva dentro; cuando tenían excremento se determinó de qué tipo de larva procedía, ya que es posible distinguir fácilmente entre el excremento de larvas de curculiónidos y de palomillas. Cuando se encontró más de una larva en una bellota, se registraron todas las larvas que procedían de una misma bellota.
Para evaluar si existen diferencias significativas en el tamaño (volumen) de las bellotas entre las 3 especies de encino, se realizó un Andeva de una vía y se obtuvieron intervalos de confianza de 95%. Por otra parte, para analizar el efecto del tamaño de las bellotas y la especie de Quercus en la depredación predispersión total (i.e., la realizada por larvas de curculiónidos y de palomillas), se usaron modelos lineales generalizados de efectos mixtos (GLMM) con distribución binomial, usando el software R (R Core Team, 2018). Se probaron 2 modelos: uno con interacción entre las variables y uno aditivo (i.e., sin interacción). Éstos se compararon utilizando pruebas de razón de verosimilitud, observando si al eliminar la interacción se generaban diferencias significativas entre ellos. En ambos se usaron como variables explicativas la especie de encino (Q. rugosa, Q. obtusata y Q. castanea) y el volumen de cada bellota, y se incluyó como factor aleatorio el árbol (individuo). La variable de respuesta fue la condición de la bellota, con 2 categorías (sana o depredada). En el análisis se usaron los datos de todas las bellotas, excluyendo solamente a las depredadas por ambos tipos de organismos (curculiónidos y palomillas en la misma bellota). Estas bellotas se excluyeron de éste y todos los análisis subsecuentes debido a que representaban un porcentaje muy bajo (3%) de la muestra total (e incluso < 1% en Q. castanea), y su inclusión dificultaba el análisis.
Para evaluar si existen diferencias en la proporción de bellotas depredadas, ya sea por larvas de curculiónidos o de palomillas, se elaboraron 4 modelos GLMM con distribución binomial: uno aditivo, uno con interacción entre variables y 2 con una sola variable explicativa, la especie de Quercus o el volumen de las bellotas. La variable de respuesta fue la proporción de bellotas depredadas, con 2 categorías (depredadas por curculiónidos o por palomillas). Al igual que en los modelos anteriores, se usaron como variables explicativas la especie de encino y el volumen de la bellota, y como factor aleatorio el árbol. En este análisis se usó la muestra de bellotas depredadas (i.e., se excluyó a las sanas). Una vez seleccionado el modelo, se hizo la trasformación logarítmica de los estimadores, para obtener valores entre 0 y 1, con la fórmula:
donde Y es la probabilidad de que una bellota sea depredada por un curculiónido y z es el modelo lineal seleccionado para este tipo de depredación (z = ßx1 + ßx2). Por lo tanto, la probabilidad de depredación por palomillas corresponde a 1 - Y. De los estimadores del modelo seleccionado, se obtuvieron los intervalos de confianza, que también se transformaron con las fórmulas antes descritas. En todos los estadísticos se estableció como valor crítico p = 0.05.
La superinfestación se analizó comparando las proporciones de bellotas superinfestadas en cada especie (usando como muestra el número total de bellotas en que se pudo registrar el número de larvas de curculiónidos presentes) y mostrando la proporción de bellotas con diferente número de larvas (i.e., cuántas tenían 2, 3 o más larvas por bellota) en cada especie de Quercus.
Resultados
Se recolectaron 961 bellotas, de las cuales 42.6% (409) fueron depredadas: 34.3% (330) por larvas de curculiónidos, 5.2% (50) por larvas de palomillas y 3.0% (29) con depredación por ambos tipos de larvas. La mayor depredación de bellotas se registró en Q. obtusata, especie en la que 69.4% de las bellotas recolectadas fueron depredadas (60.0% por curculiónidos, 3.6% por palomillas y 5.8% por ambos tipos de organismos). Las bellotas de Q. rugosa tuvieron un nivel de depredación intermedio, de 37.3% (31.9% por curculiónidos, 2.6% por palomillas y 2.8% por ambos tipos de insectos), y en las de Q. castanea se registró el nivel más bajo de depredación, 24.6% (13.8% por larvas de curculiónidos, 10.1% por larvas de Cydia y 0.7% por ambos tipos de larvas).
Los niveles de depredación registrados guardan relación con el volumen promedio de las bellotas, que en el caso de Q. obtusata fue de 2.6 cm3 (± e.e. 0.9, n = 275); en Q. rugosa fue de 1.15 cm3 (± 0.7, n = 389) y el más pequeño se registró en Q. castanea (0.43 cm3 ± 0.3, n = 297). Las diferencias en el volumen de las bellotas de las 3 especies fueron significativas (F2, 958 = 747.2, p < 0.001).
Los GLMM para analizar el efecto de la especie de encino y el volumen de la bellota en la depredación total (i.e., la realizada por ambos tipos de organismos) se muestran en la tabla 1. Se compararon 2 modelos, uno aditivo y uno con interacción entre ambas variables, y se observó que al eliminar la interacción se generaron diferencias significativas entre ambos, por lo que se seleccionó el modelo que la incluye. Esta interacción se debe a que en una especie (Q. rugosa) hay una tendencia a que la probabilidad de depredación aumente con el volumen de las semillas (i.e., la pendiente fue significativamente distinta de cero, p < 0.0001; fig. 1), que no se presentó en las otras 2 especies de Quercus (Q. obtusata, p = 0.17 y Q. castanea, p = 0.7). En la figura 1 se observa que en Q. obtusata la probabilidad de depredación fue alta en todo el intervalo de volumen, de alrededor de 0.6 en las bellotas más pequeñas hasta cerca de 0.8 en las más grandes, mientras que en Q. rugosa, la variación en la probabilidad de depredación fue de 0.2 en las bellotas más pequeñas hasta > 0.8 en las más grandes. En Q. castanea, la probabilidad fue baja (< 0.3) en todo el intervalo de volumen de sus semillas, que no fue muy amplio. En este modelo, el efecto del volumen de las bellotas explica una mayor proporción de la devianza (χ2 = 12.4, g. l. 1, p = 0.0004), que la especie de encino (χ2 = 10.6, g. l. 2, p = 0.005) y que la interacción χ2 = 6.8, g. l. 2, p = 0.033).
Tabla 1 Modelos lineales generalizados de efectos mixtos para analizar el efecto de la especie y el volumen de la bellota en la depredación predispersión de bellotas de 3 especies de encino. Valor crítico p = 0.05
| Variable de respuesta | Modelo GLMM | p | Tamaño de muestra |
|---|---|---|---|
| Depredación total | depredada = volumen × especie + árbol vs. depredada = volumen + especie + árbol |
0.04 | N = 961 |
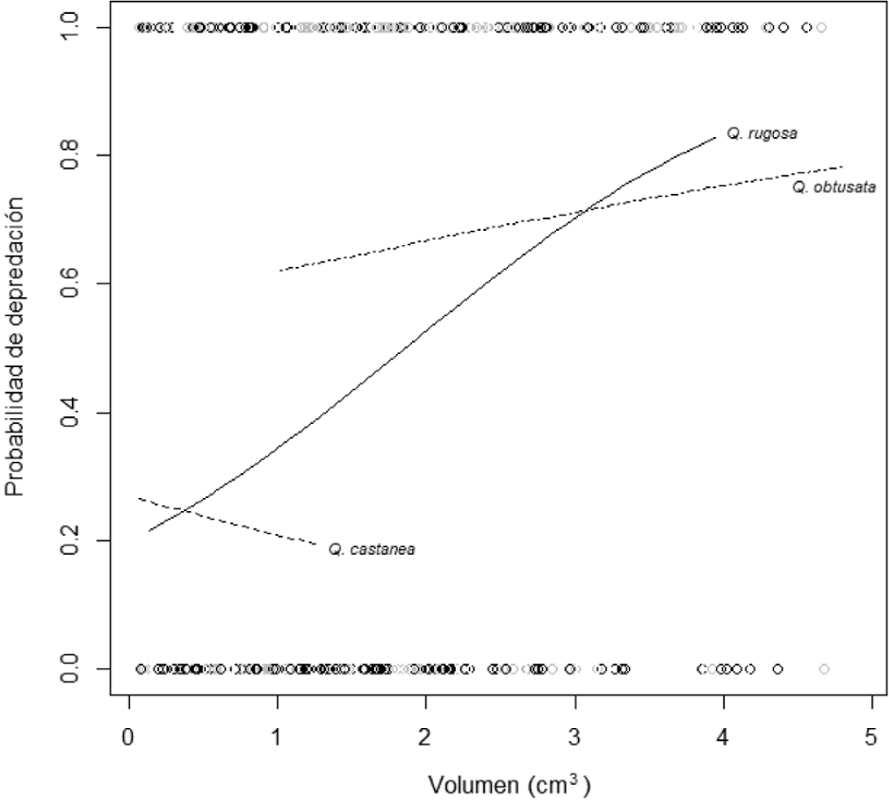
Figura 1 Efecto del volumen de la bellota en la probabilidad de depredación predispersión de bellotas de Q. rugosa, Q. obtusata y Q. castanea.
Para analizar el efecto de la especie de encino y el volumen de las semillas en la proporción de bellotas depredadas por larvas de curculiónidos o por larvas de palomillas, también se elaboraron y compararon 2 GLMM, uno aditivo y otro con interacción entre las variables. Debido a que las diferencias entre ambos no fueron significativas (p = 0.06, tabla 2), se usó el modelo aditivo, por ser más sencillo. Posteriormente, se comparó un modelo aditivo que incluye ambas variables con otro en el que se eliminó el volumen de las bellotas y no se observaron diferencias significativas. Sin embargo, al comparar el modelo con ambas variables con uno en que se eliminó la especie de encino, sí las hubo (tabla 2). Ésto indica que el mejor modelo tiene como única variable explicativa la especie de Quercus (χ2 = 47.6, g. l. 2, p = 4.6 e-11).
La proporción de bellotas depredadas solo por curculiónidos fue similar en Q. rugosa (promedio 0.94, 0.96 - 0.87, máximo y mínimo, respectivamente) y Q. obtusata (promedio 0.92, máximo 0.97, mínimo 0.90) y significativamente mayor que en Q. castanea (promedio 0.57, máximo 0.72, mínimo 0.31; fig. 2). Por otro lado, la proporción de bellotas depredadas por larvas de palomillas fue mayor en Q. castanea (promedio 0.42, máximo 0.68, mínimo 0.28), que en Q. obtusata y Q. rugosa, que registraron valores menores y similares (promedios 0.07 y 0.05, máximos 0.1, y 0.13, y mínimos 0.02 y 0.03, respectivamente).
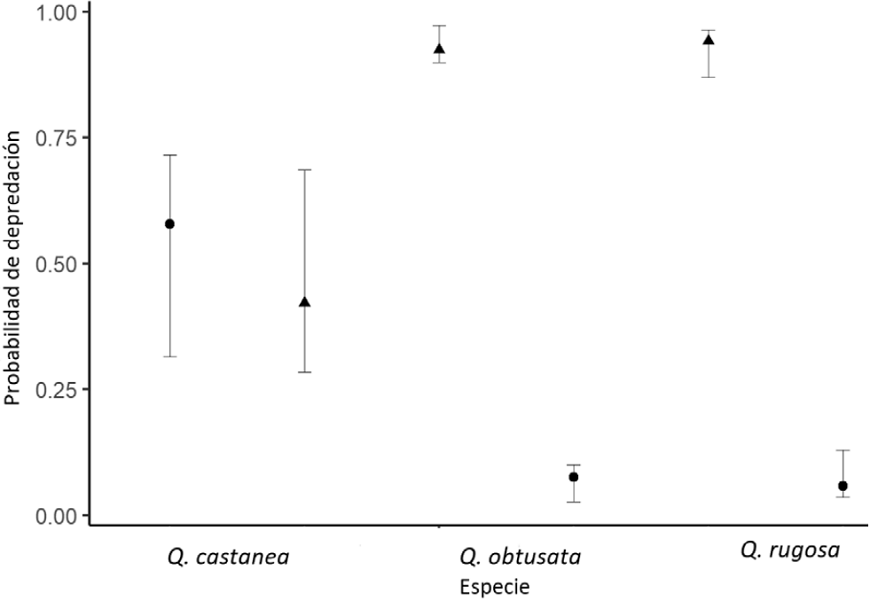
Figura 2 Probabilidad de depredación predispersión por larvas de curculiónidos (tríangulos) y de Cydia (círculos) en 3 especies de Quercus. Las barras denotan intervalos de confianza de 95%.
Tabla 2 Modelos lineales generalizados de efectos mixtos elaborados para analizar el efecto del volumen de la bellota y la especie de encino en la proporción de bellotas depredadas por larvas de curculiónidos o de Cydia. Valor crítico p = 0.05
| Variable de respuesta | Modelo GLMM | p | Tamaño de muestra |
|---|---|---|---|
| Proporción de bellotas depredadas por curculiónidos (Curc) o palomillas (Cydia) |
(Curc, Cydia) = volumen × especie + árbol vs. (Curc, Cydia) = volumen + especie + árbol |
0.06 | N = 375 |
| (Curc, Cydia) = volumen + especie + árbol vs. (Curc, Cydia) = especie + árbol |
0.67 | ||
| (Curc, Cydia) = volumen + especie + árbol vs. (Curc, Cydia) = volumen + árbol |
< 0.001 |
En cuanto a la superinfestación, los niveles más altos se registraron en Q. obtusata, en la que 56.6% de las bellotas depredadas por curculiónidos (considerando solo aquéllas en que se pudo observar y contabilizar directamente a las larvas) tuvo más de 1 larva. En Q. rugosa, la superinfestación se presentó en 33% de las bellotas depredadas por curculiónidos, y en Q. castanea en 29.4%. En la primera especie, además de registrarse la menor proporción de bellotas con 1 sola larva, se encontraron hasta 10 larvas en una misma bellota (fig. 3), mientras que en Q. rugosa se registró un máximo de 7, con una alta proporción de bellotas con 1 o 2 larvas. Por su parte, en Q. castanea, el máximo fue de 3 larvas por bellota, y registró la proporción más alta de bellotas con 1 sola larva (fig. 3). Ésto muestra que hay mayor superinfestación y un mayor número de larvas por bellota en la especie con las bellotas más grandes, y que este fenómeno es más raro y limitado en la especie con bellotas más pequeñas. Al interior de cada especie de encino no hubo diferencias significativas en el volumen de las bellotas con 1 sola larva y las que presentaban 2 o más (Q. rugosa t = 1.24, g. l. = 81, p = 0.21; Q. castanea U12,5 = 27, p = 0.79; Q. obtusata t = -1.77, g. l. = 136, p = 0.07).

Discusión
En este trabajo se mostró que los niveles de depredación predispersión se encuentran directamente relacionados con el volumen promedio de las bellotas de 3 especies de encinos que coexisten. Este patrón se debe a las preferencias de los curculiónidos -el principal grupo implicado en esta interacción- por ovipositar en las especies con bellotas más grandes, lo que permite que sus larvas puedan tener acceso a una mayor cantidad de sustancias de reserva de los cotiledones. Se mostró también que en las palomillas del género Cydia no se presenta una selección de las bellotas más grandes, ni al comparar entre especies, ni al interior de la especie que registró una mayor abundancia de sus larvas, Q. castanea. Por el contrario, las palomillas prefieren a la especie con bellotas más pequeñas. Aunque esta relación es muy significativa, es difícil saber si se debe a que prefieren bellotas más pequeñas per se, o a que en esta especie encuentran menos competencia con los curculiónidos, o a que seleccionan otras características de las mismas, ya sean físicas -como el grosor del pericarpio- o químicas, como el tipo y concentración de taninos, que afectan las preferencias alimenticias de los herbívoros (Robbins et al., 1987a, b; Steele et al., 1993), incluyendo larvas de curculiónidos (Weckerly et al., 1989).
La mayor abundancia de larvas de curculiónidos que de palomillas en las bellotas es un fenómeno común, que se ha reportado en diversas especies: Q. garryana en Canadá (Rolphs, 1999), Q. rotundifolia en España (Soria, Villagrán et al. 1999), y Q. magnoliifolia y Q. glaucoides en México (Taracena, 2015), pero no siempre es así (Branco et al., 2002; Díaz-Fleisher et al., 2010).
Espelta, Bonal et al. (2009) y Espelta, Cortés et al. (2009) reportaron la preferencia de los curculiónidos por especies de Quercus con bellotas grandes y al comparar 2 especies, encontraron que en Q. humilis, la especie con bellotas grandes, pueden ovipositar 2 especies de curculiónidos con diferentes tamaños corporales, mientras que en Q. ilex, con bellotas más pequeñas, solo se encuentran larvas de la especie de gorgojo de menor tamaño. En nuestro caso, no se conoce la identidad de las especies de curculiónidos asociados a las especies de Quercus estudiadas, pues en las últimas décadas no se han realizado estudios taxonómicos sobre estos escarabajos en México. Sin embargo, es probable que haya más de 1 especie presente, en cuyo caso, los curculiónidos con menor tamaño corporal podrían ovipositar en especies con bellotas más pequeñas. Existe evidencia -en un estudio no publicado- de que a través de métodos moleculares, se detectó la presencia de larvas de varias especies de curculiónidos en 2 especies de encinos de la sierra del Chichinautzin (Taracena, 2015), lo que nos permite suponer que puede haber más de 1 especie de curculiónido en los encinares de esta zona del Ajusco, que forma parte de la misma cadena montañosa. De cualquier forma, los datos indican, que hasta cierto punto, los curculiónidos evitan ovipositar en Q. castanea, que solo presentó larvas de escarabajos en alrededor de 14% de sus bellotas, mientras que este porcentaje fue cercano a 32% y 60% en Q. rugosa y Q. obtusata, respectivamente.
La presencia de larvas de curculiónidos no parece inhibir la depredación por palomillas en Q. rugosa y Q. obtusata, pues los porcentajes de larvas o heces de ambos tipos de insectos presentes en una misma bellota fueron similares a los registrados sólo para palomillas. En contraste, en Q. castanea la presencia de larvas o heces de ambas especies se registró en menos de 1% de las bellotas, lo que sugiere que su tamaño pequeño restringe la presencia simultánea de ambos tipos de depredadores.
La probabilidad de infestación aumentó con el volumen de la bellota de forma significativa sólo en una especie, Q. rugosa. Un estudio previo realizado con esta especie mostró que la pérdida parcial de los cotiledones afecta mucho más a la germinación y el crecimiento de plántulas provenientes de bellotas pequeñas que a las que provienen de bellotas grandes (Bonfil, 1998), por lo que la depredación de las bellotas grandes no necesariamente implica que pierdan su capacidad de germinar (Branco et al., 2002; Fukumoto y Kajimura, 2000). Es muy probable que lo mismo suceda con Q. obtusata, cuyas bellotas son incluso mayores. Dado que esta última especie presentó una alta proporción de bellotas depredadas en todo el intervalo de volumen (fig. 1), es probable las hembras de curculiónidos no seleccionen a las bellotas una vez superado cierto umbral de tamaño, como se ha reportado en 2 especies de Curculio de España (Espelta, Cortés et al., 2009).
Con respecto a la superinfestación, destaca que la mayor proporción de bellotas con más de 1 larva, así como el mayor número de larvas por bellota, se presentaron en la especie con bellotas más grandes, mientras que la especie de encino con las bellotas más pequeñas presentó un nivel de superinfestación mucho menor y el número máximo de larvas por bellota más bajo. Hasta donde sabemos, es la primera vez que se hacen comparaciones de este fenómeno entre más de 2 especies de Quercus que coexisten y que presentan diferencias tan marcadas en el tamaño de las bellotas, y las tendencias observadas son muy claras a que la superinfestación se concentre en la especie con las bellotas más grandes. Al interior de las especies de estudio no hubo evidencia de que las hembras de curculiónidos seleccionaran a las bellotas más grandes para una segunda o tercera oviposición en una misma bellota.
Este estudio representa un primer acercamiento para conocer la magnitud de la depredación predispersión y la superinfestación en 3 especies de encinos que coexisten, así como las preferencias de distintos tipos de depredadores y algunos factores que contribuyen a explicarlas.

