











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
México, al igual que el resto del mundo, atraviesa una crisis de la biodiversidad debido a un aumento en la tasa de extinción de especies causada por factores como la contaminación, sobreexplotación, introducción de especies exóticas y, especialmente, la pérdida de los hábitats naturales (Martínez-Meyer et al., 2013). Además, este problema se exacerba porque todavía existen muchos vacíos en nuestro conocimiento de la diversidad de especies, especialmente en el ámbito local; por ejemplo, para muchas regiones aún no existen inventarios o estudios adecuados sobre su biodiversidad, por lo que es importante continuar con los estudios a esta escala (Gallardo y Aguilar-Rodríguez, 2011; Rojas-Soto y de Ita, 2005; Villa-Bonilla et al., 2008).
En México se ha documentado la presencia de 1,150 especies de aves, alrededor de 10% del total mundial (Gill y Donsker, 2017; Navarro-Sigüenza et al., 2014; Robles et al., 1989). Esta diversidad se debe a la gran variedad de climas y ecosistemas que hay en nuestro territorio, producto de su compleja orografía, y a la confluencia de las regiones biogeográficas Neártica y Neotropical (Escalante et al., 1993; Navarro-Sigüenza et al., 2014). En el estado de Veracruz se ven reflejadas esta biodiversidad y complejidad geográfica, ya que encontramos áreas de costa, planicies y zonas montañosas surcadas por cañadas (Alcántara-Carbajal, 1993; Gallardo y Aguilar-Rodríguez, 2011; Hernández-Baz e Iglesias, 2001; Moreno-Casasola et al., 2010; Villaseñor, 2003).
Veracruz es uno de los estados con mayor número de investigaciones ornitológicas publicadas (Alcántara, 1993; Bojorges-Baños y López-Mata, 2001; Ferrari-Pérez, 1886; Loetscher, 1941, 1955; Lowery y Dalquest, 1951; Rodríguez-Yáñez et al., 1994; Ruelas-Inzunza, 2007; Stresemann, 1954), en parte debido a que desde el siglo XVI el puerto de Veracruz fue un importante punto de llegada para naturalistas e investigadores extranjeros y una transitada vía de comunicación con la capital del país (González, 1999). Asimismo, algunos de los temas recientes de especial interés para los investigadores, han sido las aves migratorias (p. ej., Blidstein, 2004; Loetscher, 1941; Ruelas-Inzunza et al., 2010) y las plantaciones de café como refugios de la biodiversidad (p. ej., Cruz-Angón y Greenberg, 2005; Gallina et al., 1996; Gordon et al., 2007; Leyequien et al., 2010; Moguel y Toledo, 1999; Pineda et al., 2005).
Algunos estudios avifaunísticos se han realizado en regiones cercanas al municipio de Misantla (Bojorges-Baños y López-Mata, 2001, 2005). Esta región, por su localización geográfica en el centro norte del estado y en el paso entre la Cd. de México y el puerto de Veracruz, ha sido moderadamente estudiada desde el punto de vista de su diversidad biológica. Destacan los estudios realizados en botánica (Lascuráin et al., 2007; Zamora y Avedaño, 1998), de los cuales el trabajo de investigación más completo sobre la caracterización de la vegetación de Misantla lo realizó Gómez-Pompa (1966), en el que describe las principales comunidades vegetales de la zona.
Rzedowski (2006) basándose en el trabajo de Gómez-Pompa, además identifica para Misantla a las selvas de Pseudolmedia oxyphyllaria y de Brosimum alicastrum como bosques tropicales perennifolios y a la selva de lauráceas como bosque mesófilo de montaña según su esquema de clasificación.
El estudio científico de las aves de Misantla se inició formalmente en el siglo XIX con el trabajo de algunos exploradores y recolectores como Ferdinand Deppe y Frederic D. Godman (Alcántara-Carbajal, 1993; Stresemann, 1954; Warner y Harrell, 1957), sin embargo, a la fecha no se ha publicado un estudio avifaunístico completo del municipio y sus áreas de influencia (Martínez-Gómez, 1996; Rodríguez-Yáñez et al., 1994). En el presente trabajo presentamos un inventario completo de las aves de la región. Con base en esta información, caracterizamos la riqueza y composición de especies por localidad y tipo de vegetación. Adicionalmente, comparamos la diversidad actual con la reportada por los naturalistas del siglo XIX para evaluar el estado de conservación actual de las comunidades de aves.
Materiales y métodos
El área de estudio comprendió al municipio de Misantla, ubicado en la región centro-norte del estado de Veracruz, aproximadamente a 40 km al norte de la ciudad de Xalapa, en la región centro-norte del estado de Veracruz (fig. 1). La cabecera municipal y principal asentamiento es la ciudad de Misantla que se encuentra en las coordenadas 19.93° N, -96.85° O, a una altitud de 300 m (EMM, 2005). El municipio pertenece a la cuenca hidrológica Nautla y Otros, de la vertiente del golfo (CNA, 1998) y es atravesado por el río Misantla, al oeste se encuentra el río Quilate y al norte el río Nautla. El resto del municipio es recorrido por una multitud de arroyos, afluentes de los ríos principales (CNA, 1998).

Figura 1 Ubicación del municipio de Misantla y localidades muestreadas. A) Ciudad de Misantla, B) Ignacio Zaragoza, C) cerro Quebrado, D) cerro Gordo, E) Los Ídolos, F) Pueblo Viejo, G) Gutiérrez Nájera, H) Díaz Mirón, I) Villa Nueva.
Orográficamente, la zona norte del municipio es de baja altitud y está dominada por planicies por debajo de los 600 m snm como el cerro de la Espaldilla y el cerro Quebrado. La zona sur corresponde a una cañada profunda flanqueada por 2 líneas montañosas derivadas de la sierra de Chiconquiaco que alcanzan los 1,800 m snm (fig. 1). El municipio de Misantla, con una superficie de 538 km2, se encuentra además en la zona de colindancia de la provincia biogeográfica de la Planicie Costera del Golfo y la provincia de la Sierra Madre Oriental (Morrone, 2005). Existe un gradiente de menor a mayor precipitación de la zona costera hacia el interior y de norte a sur. Además, la sierra de Chiconquiaco actúa como un frente de lluvia generando una mayor precipitación en sus faldas. El gradiente de temperatura también tiene una dirección noreste-suroeste de mayor a menor y se encuentra ligada a las diferencias de altitud (Gómez-Pompa, 1966). La vegetación original fue bosque tropical perennifolio en las tierras bajas, bosque mesófilo de montaña y vegetación ribereña en el borde de los ríos (Gómez-Pompa, 1966; Rzedowski, 2006).
El muestreo de campo lo realizamos de marzo de 2010 a abril de 2011. Elegimos las localidades a muestrear de manera que abarcaran todos los tipos de vegetación, tanto primaria, como aquella con distinto grado de perturbación (tabla 1).
Tabla 1 Localidades visitadas y tipo de vegetación encontrados en cada una. BT= Bosque tropical perennifolio. BM= Bosque mesófilo de montaña. VR= Vegetación ribereña. SBT= Vegetación secundaria derivada de bosque tropical perennifolio. SBM= Vegetación secundaria derivad de bosque mesófilo de montaña. CA= Cafetal de sombra. CU= Cultivos. PG= Pastizal ganadero. PU= Pueblos
| Localidad | Coordenadas (N, O) | Altitud m snm | Tipos de vegetación |
| Cerro Gordo | 19.935°, 96.769° | 378 | BT, VR, SBT, CU, PG, PU |
| Cerro Quebrado | 20.009°, 96.916° | 434 | BT, VR, SBT CA, CU, PG, PU |
| Díaz Mirón | 19.785°, 96.872° | 865 | BM, SBM, CA, CU, PG, PU |
| Gutiérrez Nájera | 19.792°, 96.861° | 846 | VR, SBM, CA, CU, PU |
| Ignácio Zaragoza, 1.7 km O | 19.983°, 96.963° | 137 | VR, SBT, CU, PG |
| Los Ídolos | 19.894, 96.857° | 373 | VR, SBT, CA, CU, PG, PU |
| Misantla, alrededores | 19.333, 96.85° | 300 | VR, CU, PG |
| Pueblo Viejo | 19.815°, 96.872° | 730 | BM, SBM, VR, CA, CU, PG, PU |
| Villa Nueva | 19.773°, 96.867° | 970 | BM, SBM, VR, CA, CU, PG, PU |
La información sobre la composición de las aves se obtuvo a través de puntos de conteo de radio fijo de 50 m, ubicados cada 150 m con el fin de minimizar las posibilidades de reconteo (Ralph et al., 1996). A lo largo de una ruta de puntos de conteo de aproximadamente 1,500 m de longitud, registramos cada especie de ave identificada por avistamiento, cantos o llamados, anotamos el número de individuos y el tipo de vegetación en que fueron observados. En cada sitio de muestreo se realizaron 4 rutas de puntos, procurando seguir los puntos cardinales. Las observaciones fueron realizadas por una sola persona (AOV) y cada localidad fue visitada en 2 ocasiones para abarcar tanto la temporada reproductiva como la de migración. Realizamos una recolecta científica selectiva (permiso de colecta FAUT-0169), con el fin de formar una colección de referencia de las aves de la zona mediante el uso de redes de niebla de 12 m de largo por 3 m de alto. Las redes fueron ubicadas en terrenos particulares alejados de la ruta de puntos de conteo, en momentos diferentes respecto a las observaciones. Los ejemplares capturados fueron recolectados y depositados para su conservación en la colección ornitológica del Museo de Zoología “Alfonso L. Herrera” de la Facultad de Ciencias de la Universidad Nacional Autónoma de México (MZFC-AV). Además, con el fin de detectar especies no registradas por los otros métodos, realizamos observaciones libres fuera de las redes, puntos de conteo y durante la estancia en la zona de estudio. La lista de especies resultante sigue el orden taxonómico propuesto por el International Ornithological Committee (Gill y Donsker, 2017)
Para evaluar qué tan completo fue nuestro muestreo, estimamos una curva de acumulación de especies basada en los registros por salida aleatorizados, usando el programa EstimateS (Colwell, 2013) para reducir el sesgo por el orden en que se registraron las especies. A partir de la curva estimada, utilizamos el modelo de Clench (Clench, 1979; Soberón y Llorente, 1993) para estimar la riqueza esperada. Como parte de la caracterización de la diversidad de aves de la región, obtuvimos de la literatura el estatus de estacionalidad (de acuerdo con Howell y Webb, 1995), endemismo (siguiendo a González-García y Gómez-de Silva, 2003), conservación (IUCN, 2017; Semarnat, 2010) y si se trataba de especies exóticas (Conabio, 2016).
Para estimar la abundancia relativa utilizamos el método de porcentaje de representatividad (Pettingill, 1970), comparando el número de días en que se registró cada especie residente con el número de días de trabajo totales (Howell y Webb, 1995), el número de días que se registró cada especie migratoria con los días de trabajo correspondientes a la época de presencia de las especies visitantes de invierno y las reproductoras de verano, según fuera el caso. Para las especies accidentales y transeúntes se utilizó como referencia el número de días en las salidas en que fueron registradas respecto al número de días de trabajo de esas salidas. Debido a que no obtuvimos valores de representatividad de 100%, ajustamos las categorías de abundancia relativa y las definimos de la siguiente forma: rara de 0 a 9%, no común de 10 a 27%, común de 28 a 81% y abundante de 81 a 90%.
Para conocer la similitud en la composición de las comunidades de aves, usamos los datos de presencia-ausencia de especies por localidad para generar una matriz de disimilitud utilizando el coeficiente de Sørensen (Koleff et al., 2003; Sørensen, 1948), y con ella generar (empleando el promedio de sus valores) un dendrograma mediante el método jerárquico de ligamiento promedio (Sokal y Michener, 1958)
Para conocer la variación en la composición de especies de aves por tipo de vegetación, conformamos un dendrograma de disimilitud siguiendo el mismo método usado en el análisis de las localidades. Los tipos de vegetación naturales, de acuerdo con Gómez-Pompa (1966) y Rzedowski (2006) y observaciones propias, fueron: 1) bosque tropical perennifolio (BTP), 2) bosque mesófilo de montaña (BMM), 3) vegetación ribereña (VR).
Los tipos de vegetación modificados fueron: 1) vegetación secundaria de bosque tropical perennifolio (SBTP), 2) vegetación secundaria de bosque mesófilo de montaña (SBMM), 3) cafetal de sombra (CA), 4) cultivos y sus bordes (CU), 5) pastizal ganadero y sus bordes (PG), 6) pueblos (PU), 7) zona urbana (ZU) que corresponde a la ciudad Misantla. Dado que en la ciudad no se realizó un muestreo sistemático, no se le incluye en los análisis de los tipos de vegetación, pero se le reporta en el listado de especies (apéndice 1).
Para explorar si existía independencia entre la abundancia relativa de las especies y la cantidad de tipos de vegetación que puede ocupar, realizamos una prueba gama de Goodman-Kruskal, que mide el grado de relación entre variables categóricas ordinales en una tabla de contingencia (Goodman y Kruskal, 1954). También realizamos curvas de rango abundancia o curvas de Whittaker (Feinsinger, 2001), para evaluar de forma gráfica el grado de dominancia y las especies dominantes por tipo de vegetación.
Para conocer la composición de las comunidades de aves anterior a este estudio, llevamos a cabo una búsqueda amplia de literatura (Loetscher, 1955; Miller et al., 1957; Warner y Harrell, 1957) y en bases de datos especializadas (GBIF, 2017; Navarro-Sigüenza et al., 2003; VertNet, 2017) de las que se recabó información de ejemplares de la región depositados en las colecciones biológicas del mundo. Para comparar los cambios en la avifauna, primero, elaboramos una base de datos que integró todos los registros de encontrados tanto en la literatura como en el presente estudio. Posteriormente, para cada especie, se homogenizaron las siguientes variables utilizando únicamente los valores reportados en la literatura especializada: tipo de vegetación: (F-bosques y selvas, E-bordes de bosques y selvas, N-hábitat abiertos, A-acuático; Parker et al., 1996); estacionalidad (R-residente permanente, W-visitante de invierno, S-residente de verano, T-transeúnte, A-accidental; Howell y Webb, 1995); categoría de protección (P-en peligro, A-amenazada, Pr-bajo protección especial; Semarnat, 2010); época (siglo XIX, para todos los registros realizados entre 1829 y 1897; GBIF, 2017; Loetscher, 1955; Miller et al., 1957; Navarro-Sigüenza et al., 2003; Warner y Harrell, 1957; VertNet, 2017) y siglo XXI, para todos los registros realizados entre 2009 y 2014 (eBird, 2017; GBIF, 2017). Del siglo XX, únicamente encontramos 3 registros de 1956 y 1965 (GBIF, 2017), por lo que los asignamos al siglo XXI por su mayor cercanía temporal. Para evaluar las diferencias en la composición de especies el siglo XIX y XXI, realizamos una prueba de t pareada para cada variable explicativa, (categoría de hábitat, estacionalidad y de protección), usando la proporción de especies como variable de dependiente. Todas estas pruebas y análisis fueron realizadas utilizando el software R (R Core Team, 2013) y los paquetes vegan (Oksanen et al., 2013) y vcdExtra (Friendly, 2016).
Resultados
Durante el presente estudio se obtuvieron 6,412 registros de 161 especies de aves pertenecientes a 40 familias y 15 órdenes (apéndice 1). Con 3,072 horas red, recolectamos 110 ejemplares de 63 especies pertenecientes a 17 familias y a 5 órdenes. Estos ejemplares se integraron a la colección de aves del Museo de Zoología “Alfonso L. Herrera” de la Facultad de Ciencias con los números de catálogo MZFC-AV 24145 a 24254.
La curva de acumulación de especies no llega a una asíntota y con el modelo de Clench estimamos 207 especies, por lo que se infiere que durante el presente trabajo registramos el 77.7% de las especies esperadas (fig. 2). Del total de las especies, 104 son residentes permanentes y 56 migratorias y, de éstas últimas, 5 son residentes de verano, 45 son visitantes de invierno y 6 son transitorias. Registramos 4 especies cuasiendémicas (Campylopterus curvipennis, Amazilia yucatanensis, Icterus graduacauda y Arremonops rufivirgatus), 2 semiendémicas (Empidonax oberholseri e Icterus cucullatus) y 3 exóticas invasoras (Bubulcus ibis, Columba livia y Passer domesticus). Además, registramos 24 especies en categoría de protección, lo que representa un 14% del total. La mayor parte de las especies presentaron valores bajos de abundancia relativa, solo Cardellina pusilla fue abundante y 95 especies raras (apéndice 1).

Figura 2 Curva de acumulación de especies. Los círculos negros muestran el número estimado de especies acumuladas por número de salida según el resultado de la aleatorización realizada por el software EstimateS (Colwell, 2013). La línea continua corresponde al modelo de Clench (a = 85.7639, b = 0.4137) ajustado a los datos, con el cual se estima 207 especies esperadas (a/b = 207.2745).
Las localidades con un mayor número de especies fueron Los Ídolos con 87 y Villa Nueva con 82. Las localidades con menos registros fueron Gutiérrez Nájera con 38 y cerro Gordo con 41 especies. El dendrograma de disimilitud entre localidades muestra que las principales agrupaciones corresponden a la altitud y tipo de vegetación primaria en que se encuentran, de hecho, parecen corresponder a 3 pisos altitudinales, dado que la primera está formada por las localidades de la zona baja del municipio (Ignacio Zaragoza a 1.7 km O, cerro Gordo, los alrededores de la ciudad de Misantla, cerro Quebrado y Los Ídolos); todas ellas con elevaciones de 100 a 500 m snm. Otro grupo está formado por las localidades de la zona alta del municipio; ésto es, Díaz Mirón, Gutiérrez Nájera y Pueblo Viejo, que se encuentran a altitudes de 600 a 950 m snm, mientras que la localidad de Villa Nueva (970 m snm) se encuentra en un tercer piso altitudinal (tabla 1, fig. 3).
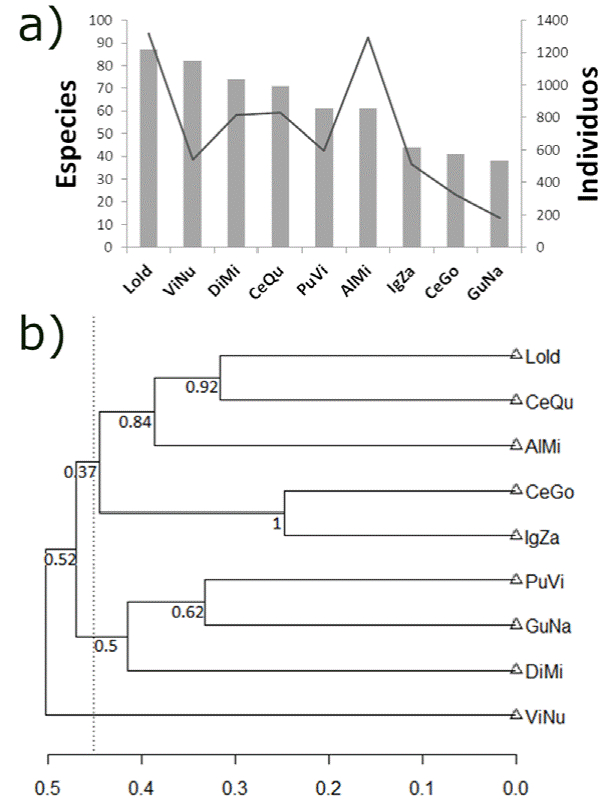
Figura 3 Riqueza y abundancia de especies por localidad. a) Las barras corresponden al número de especies en cada localidad mientras que la línea continua al número de individuos. b) Dendrograma de disimilitud (índice de Sørensen y método de ligamiento promedio) (promedio disimilitud = 0.45) entre localidades. En los nodos se muestra el valor de soporte de ramas (bootstrap, 1,000 repeticiones). LoId-Los Ídolos, ViNu-Villa Nueva, DiMi-Díaz Mirón, CeQu-cerro Quebrado, PuVi-Pueblo Viejo, AlMi-Alrededores de Misantla, IgZa-Ignacio Zaragoza, CeGo-cerro Gordo, GuNa-Gutiérrez Nájera.
El tipo de vegetación con mayor número de especies de aves fue el cafetal de sombra con 87 especies, seguido del pastizal ganadero con 84. Los tipos de vegetación con menor número de especies fueron el bosque tropical perennifolio, el bosque mesófilo de montaña y los tipos de vegetación secundaria derivada de ambos. En el dendrograma de disimilitud entre tipos de vegetación (fig. 4) se observa que la línea de referencia separa a la mayoría de los tipos de vegetación naturales y la vegetación secundaria en ramas independientes mientras que se forma un grupo que incluye al bosque tropical perennifolio como rama hermana de los tipos vegetación más perturbados (pastizal ganadero, cultivos, cafetal de sombra y pueblos).

Figura 4 Riqueza y abundancia de especies por tipo de vegetación. Las barras corresponden al número de especies en cada tipo de vegetación mientras que la línea continua al número de individuos. b) Mean = 0.68. Dendrograma de disimilitud (índice de Sørensen y método de ligamiento promedio) (promedio disimilitud = 0.68) entre hábitat. En los nodos se muestra el valor de soporte de ramas (bootstrap, 1,000 repeticiones). BT-Bosque tropical perennifolio, BM-bosque mesófilo de montaña, VR-vegetación ribereña, SBT-vegetación secundaria derivad de bosque tropical perennifolio, SBM-vegetación secundaria derivad de bosque mesófilo de montaña, CA-cafetal de sombra, CU-cultivos, PG-pastizal ganadero, PU-pueblos.
Un 77% de las especies fue registrado en menos de la mitad de los tipos de vegetación, mientras que unas pocas fueron comunes a todos. Se observa también que las especies abundantes ocuparon el mayor número de tipos de vegetación, mientras que las raras lo hicieron en un menor número de ellos (fig. 5). Fue más frecuente registrar a las especies raras en un solo hábitat, mientras que las especies no comunes lo fueron en 5 tipos de vegetación, las comunes en 6 y la única abundante en 9 hábitats diferentes. Por ende, el número de tipos hábitat que ocupa una especie y su abundancia relativa son variables dependientes (gama = 0.83, p < 0.05). Una notable excepción fue Glaucidum brasilianum, que siendo una especie categorizada como rara, se le detectó en 6 tipos de vegetación.
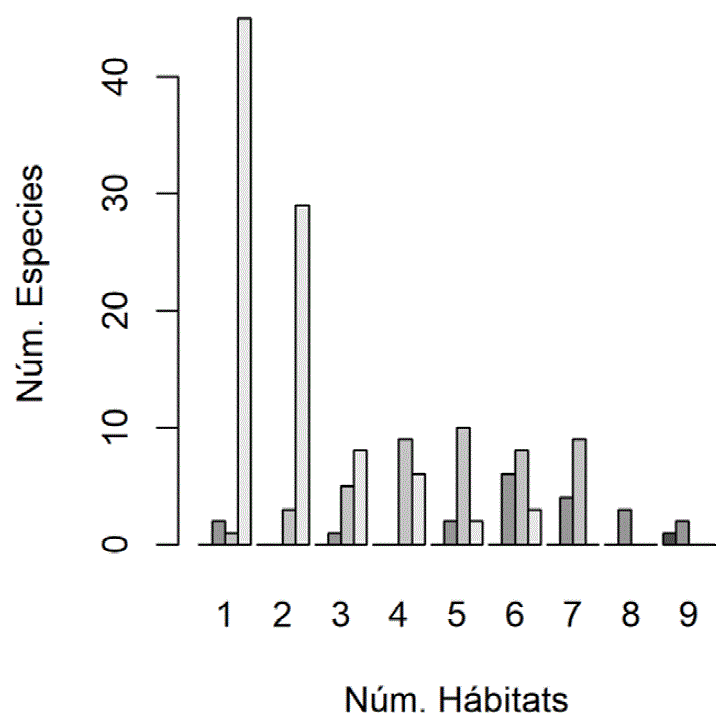
Figura 5 Número de especies en 1 a 9 tipos de vegetación. La abundancia relativa de las especies está representada por una escala de grises en las barras, siendo las barras más claras las que corresponden a la las especies raras y las más oscuras a la abundantes. Se observa que la mayoría de las especies raras son observadas en un menor número de tipos de vegetación, mientras que las más comunes se ocupan más tipos de vegetación.
Se observa que, en general, las comunidades de aves de Misantla se encuentran dominadas (fig. 6), pero hay algunos tipos de vegetación en los que el efecto es más notorio, en especial en los pueblos y en la vegetación ribereña. En los primeros destacan las especies generalistas asociadas a ambientes perturbados como Quiscalus mexicanus y Bubulcus ibis, mientras que en la vegetación ribereña son dominantes especies como Stelgidopteryx serripennis y Chaetura pelagica.

Figura 6 Curvas de dominancia de especies por tipo de vegetación. En el eje horizontal se tiene el número de especies por hábitat (rango) y en el eje vertical el logaritmo de la probabilidad individual de observar cada especie (abundancia). Las comunidades más dominadas son las que presentan una mayor pendiente en el gráfico (e.g., Pueblos), mientras que las más equitativas tienen una pendiente menor (e.g., bosque tropical perennifolio).
No obtuvimos diferencias entre la proporción de especies en el siglo XIX y XXI según los tipos de vegetación (t = -2.75, df =3 , p = 0.07), estacionalidad (t = -1.91, df = 4, p = 0.12) y protección (t = -0.79, df = 2, p = 0.51). No obstante, al comparar cualitativamente la proporción de las especies en diferentes categorías por época (fig. 7), conseguimos detectar algunos posibles patrones ecológicos. Por ejemplo, se observa un número mayor de especies acuáticas y de hábitats abiertos en la actualidad, mientras que en el siglo XIX se registraron más especies de bosque y de borde de bosque. Además, no se observaron cambios importantes en la estacionalidad, salvo por una proporción ligeramente mayor de especies transeúntes. Finalmente, se observa un porcentaje mayor de especies actualmente consideradas en peligro o amenazadas, en los estudios históricos en contraste con una proporción mayor de especies bajo protección especial en el siglo XXI.
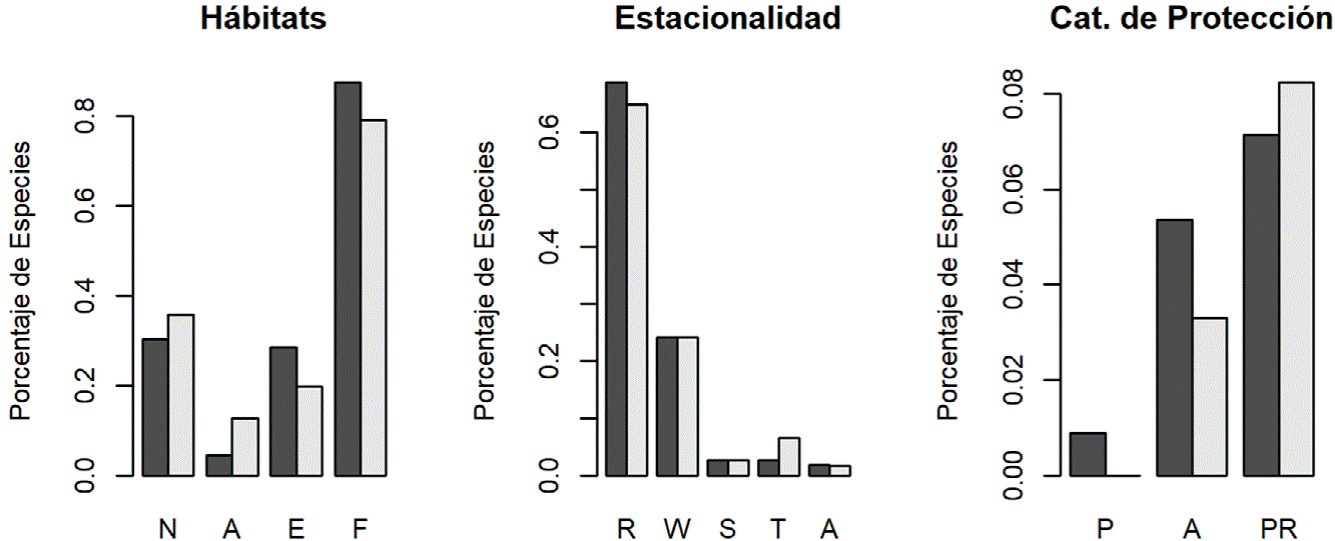
Figura 7 Porcentaje de especies registradas para cada categoría en cada época. Las barras oscuras son las especies registradas en el siglo XIX y las barras claras las registradas en el siglo XXI. No se observaron diferencias estadísticas significativas en ninguna variable: (t = -2.75, df = 3, p = 0.07), estacionalidad (t = -1.91, df = 4, p = 0.12) y protección (t = -0.79, df = 2, p = 0.51).
Discusión
El número de especies registradas durante el presente trabajo, así como la proporción de éstas con respecto a las reportadas para México, es similar al encontrado en estudios realizados en otras regiones del país, con una complejidad ambiental y a una escala semejante (538 km2) (Almazán-Núñez et al., 2009; Bojorges-Baños y López-Mata, 2001, 2005; García-Hernández, 2010; Martínez-Morales, 2007; Villa-Bonilla et al., 2008), en los que se informan entre 117 y 194 especies. Por ejemplo, muchas de las especies reportadas en el presente estudio son similares a las de la zona de protección forestal y fáunica de Santa Gertrudis, Veracruz (Bojorges-Baños y López-Mata, 2001, 2005). Además de la composición de especies, estos 2 trabajos son similares en otros aspectos como la abundancia total, la composición temporal y la abundancia relativa, siendo notoria la dominancia de las especies residentes permanentes y la existencia de un porcentaje importante de especies raras. Esta coincidencia se debe a la cercanía geográfica de ambas áreas, que es de aproximadamente 30 km, y a que comparten muchas características físicas.
Respecto a la riqueza total registrada en el presente estudio, la causa más probable de que no se alcanzaran valores cercanos a100% en el modelo de Clench fue que la mayoría de las especies son raras o poco abundantes y cabe suponer que las que teóricamente existen y no han sido registradas todavía, también lo son y, por tanto, resultan difíciles de detectar. Sin embargo, registramos todas las familias y géneros considerados omnipresentes para las aves terrestres de México (Gómez-de Silva y Medellín, 2001), con la única excepción del género Accipiter.
Detectamos que las aves que sería necesario reforzar el método para su detección serían las especies acuáticas y las nocturnas. La escasa presencia de las primeras y que requieren mayor atención se debe, en mucho, a que el municipio no posee costas, lagos ni lagunas, sin embargo, la gran cantidad de arroyos y los ríos principales, permiten la presencia de algunas de ellas como Phalacrocorax brasilianus, Cinclus mexicanus, y especies de las familias Ardeidae y Alcedinidae. En cuanto a las nocturnas, solo se detectaron por algunos llamados (Glaucidium brasilianum, Ciccaba virgata y Nyctidromus albicollis), sugerimos un método más sistemático para su registro.
Al igual que en otras zonas de Veracruz, en Misantla destaca la migración anual de aves rapaces que ocurre principalmente en abril y octubre (Ruelas-Inzunza et al., 2005). Esta migración se caracteriza por el gran número de individuos que viajan juntos y que vuelan de una manera muy visible durante el día, siguiendo las corrientes termales de aire ascendente. Esta migración está dominada por las especies Cathartes aura, Buteo platypterus y Buteo swainsoni, que son las mismas especies mencionadas como las más abundantes en durante la migración de las aves rapaces, según Bildstein (2004). Aparentemente, los individuos migratorios de ambas especies son transeúntes en la zona de estudio, por lo que no permanecen en el área.
Un registro importante fue el de un individuo de Psarocolius wagleri, especie que debe ser considerada accidental pues su área de distribución reportada en la literatura (Howell y Webb, 1995; Navarro-Sigüenza y Peterson, 2007; Peterson y Chalif, 1989) corresponde a la zona sureste del país y al extremo sur del estado de Veracruz. Sin embargo, este registro no es único ya que la especie ha sido reportada también en la región de Xalapa (eBird, 2017), lo que implicaría la ampliación del área de distribución publicada en la literatura.
Respecto a la abundancia relativa, la mayoría de las especies fueron categorizadas como raras o no comunes. Por consiguiente, cabe suponer que las especies no registradas y que fueron predichas por el modelo de acumulación, también son raras y, por tanto, requieren mayor esfuerzo y tiempo para su detección.
De acuerdo con la similitud entre comunidades de aves, observada en el dendrograma de las localidades, sugerimos que existen 2 zonas que pueden ser diferenciadas por su avifauna, además de otras características físicas y de vegetación. Esta división corresponde con las provincias biogeográficas (Morrone, 2005, 2014) presentes en el área de estudio. Con ésto afirmamos que tenemos una “zona baja” perteneciente a la provincia de la Planicie Costera del Golfo, con un clima cálido húmedo y una vegetación originaria de bosque tropical perennifolio, aunque en la actualidad se encuentra fuertemente perturbada; también tenemos una “zona alta” perteneciente a la provincia de la Sierra Madre Oriental, con un clima templado húmedo y la vegetación dominada por bosques de galería, bosques mesófilos y cultivos de café de sombra.
La localidad de Villa Nueva se muestra en una rama del dendrograma separada del resto de las localidades. Esta separación probablemente se deba a que fue la localidad de menor tamaño y la más aislada, ya que se encuentra ubicada en la base de un acantilado, y cuenta con algunas especies que solo fueron observadas ahí (e.g., Cinclus mexicanus, Aulacorhynchus prasinus). También debe considerarse la presencia de especies de aves de ambientes templados (e.g., Piranga bidentata, Passerina caerulea, Melospiza lincolni, Buteo jamaicensis), debido a que en la cima de los acantilados, en el municipio vecino de Landero y Coss, se encuentra este tipo de clima. No debe olvidarse que la Sierra Madre Oriental ha sido considerada como parte de una zona de transición entre la región Neotropical y Neártica, particularmente el intervalo de altitudes de 1,500 a 2,000 m snm en su porción sur, donde se ubica el área de estudio, ha sido reconocido como un lugar con una elevada mezcla entre ambas biotas (Ferro et al., 2017).
Para explicar por qué los hábitats modificados por actividades humanas son los que presentaron mayor riqueza de especies de aves, hay que analizar los tipos de vegetación tomando en cuenta que están distribuidos en un mosaico. Este mosaico implica que los tipos de vegetación se encuentran en pequeñas áreas colindando unos con otros, siendo así, que los más extensos, como los campos de cultivo, pastizales ganaderos y los ríos principales, pueden recibir un importante aporte de especies que se desplazan entre los diferentes tipos de vegetación sin pertenecer exclusivamente a alguno.
También es de notar que los tipos de vegetación perturbados forman un grupo en el dendrograma, a pesar de encontrarse distribuidos por toda el área de estudio y de que cada parcela puede colindar con diferentes ambientes naturales. Esto se debe a que los tipos de vegetación perturbados, tienen características similares, producto de la acción del ser humano, y por tanto, comparten una composición de especies similar (Ortega-Álvarez y MacGregor-Fors, 2009; McKinney, 2006; Le Viol et al., 2012; Vázquez-Reyes et al., 2017). El hecho de que también se agrupen con el bosque tropical perennifolio puede ser efecto de que la mayoría de las áreas perturbadas se ubiquen en la zona baja del municipio, donde predominaba este tipo de vegetación original, y puede ser evidencia de homogenización biótica en esta área.
A su vez, uno de los efectos esperados por la homogenización biótica, es un aumento en la proporción de las especies de hábitos generalistas con respecto a las especializadas en determinados tipos de vegetación, alimentación o anidación (Vázquez-Reyes et al., 2017). En este estudio se encontró una relación de dependencia entre la abundancia de una especie y la capacidad de ocupar un mayor número de ambientes, así las especies más abundantes son generalistas, dominantes en la mayor parte de los tipos de vegetación o relacionadas con tipos de vegetación modificados por la acción humana. Por ejemplo, la especie con una mayor abundancia relativa fue C. pusilla, mientras que las especies raras son, en general, las propias de los tipos de vegetación originales de la región, el bosque tropical perennifolio y el bosque mesófilo (Howell y Webb, 1995; Parker et al., 1996).
Además, la distribución de las frecuencias de cada especie por categoría de abundancia relativa según el número de tipos de vegetación en que se les registró (fig. 5) implica que las comunidades de aves, lejos de ser equitativas como podría esperarse de un hábitat conservado, se encuentran fuertemente dominadas por unas pocas especies comunes (i.e., Coragyps atratus, Cathartes aura, Psilorhinus morio, Quiscalus mexicanus y Psarocolius montezuma), especialmente en los ambientes perturbados, y la causa más probable es la acción humana (fig. 6).
Es importante mencionar que carecemos de un muestreo sistemático para los registros históricos, por lo que cualquier comparación con nuestros datos será de forma parcial. Sin embargo, podemos inferir de estos datos que los ambientes naturales de la región de Misantla se han visto afectados por actividades humanas desde mucho tiempo atrás. En tiempos prehispánicos perteneció al extremo sur del Totonacapan y sus habitantes comerciaban principalmente con productos de los bosques (González, 1999; González-de la Lama, 1990). Sin embargo, con la llegada de los españoles a la región en 1521, la población se concentró en la zona baja y se estableció una economía en la región principalmente basada en la ganadería extensiva (González, 1999; González-de la Lama, 1990). Es difícil determinar el grado de perturbación a la vegetación en esa época puesto que no existe una descripción detallada de la distribución de los parches de vegetación hasta 1966. Sin embargo, es de esperarse que en el mismo siglo XVI ya existieran amplios pastizales ganaderos en toda la zona norte del actual municipio, y desde entonces hasta la actualidad la presión sobre los ecosistemas originales no ha disminuido. Esto queda evidenciado al comparar los primeros registros científicos de aves en la región (1824 y 1888) con los actuales (1960 y 2011), ya que el número de especies por tipo de vegetación no está relacionado con la fecha de su registro. Los cambios menores que se aprecian son los que podrían esperarse de un aumento en el grado de perturbación de la zona, esto es, una disminución de las especies asociadas a los bosques y un aumento en las asociadas a zonas abiertas, producto a su vez de una probable disminución menor en la cobertura de vegetación original sustituida por áreas de ganadería y cultivo. Si bien en el siglo XIX no existía ninguna clasificación de especies en riesgo, resulta interesante hacer una aproximación a través de las categorías actuales de la NOM-059 (Semarnat, 2010). Con ello observamos la pérdida de unas pocas especies por su mayor sensibilidad a la perturbación antrópica. Debido a la falta de datos del siglo XIX, no tenemos la forma de realizar una comparación sobre las modificaciones en la abundancia de las especies de aves, sin embargo, este aspecto puede impactar severamente en la viabilidad de las poblaciones, y se ha reportado recientemente que puede ocurrir una importante caída en la abundancia de un gran número de especies de aves, aun cuando la riqueza sea similar (Pennisi, 2019).
Una revisión de la información presentada hasta ahora muestra que el municipio ha sufrido un alto grado de perturbación desde tiempos históricos y que esto se ve reflejado sobre todo en la mayor abundancia de especies generalistas y una disminución de las especies amenazadas. Sin embargo, aun permanecen zonas de bosque mesófilo de montaña, donde se registró un mayor número de especies de aves y donde se encuentran las áreas con un menor grado de perturbación. También en la sierra de Misantla existen grandes extensiones de bosque que son utilizadas cada año por las aves migratorias (Ruelas-Inzunza et al., 2005), por lo que sería importante establecer un plan de manejo para conservar la gran diversidad que existe en el área. Por otro lado, en la zona norte del municipio, las localidades de cerro Quebrado y Los Ídolos fueron las que presentaron una mayor riqueza de especies y que poseen zonas de vegetación original, por lo que también deben considerarse como prioritarias para la conservación. Estas áreas de vegetación remanente se encuentran en las zonas más escarpadas (Gómez-Pompa, 1966) y probablemente son las mismas desde tiempos históricos, lo que las vuelve factibles de conservarlas a largo plazo, especialmente si reciben un manejo de protección.
Debido a la naturaleza fragmentaria de los parches remanentes de vegetación original, el método de protección más viable sería el basado en pequeñas reservas de propiedad privada o por unidades de manejo ambiental (UMA). También se puede aprovechar de forma sustentable el gran número de recursos naturales que las comunidades de la zona alta pueden obtener de sus bosques y la importancia de éstos en la captura de agua de lluvia, afianzamiento del suelo para prevenir deslaves y reducción en la crecida de los ríos en tierras bajas, mediante un esquema de conservación por pago de servicios ecosistémicos (Almeida-Leñero et al., 2007; Camargo et al., 2012). Finalmente, no se puede dejar de lado la posibilidad de utilizar el ecoturismo de observación de aves como una forma de educación ambiental y también para generar un ingreso económico para los pobladores, favoreciendo la conservación de las aves y sus hábitats (Cantú et al., 2011). El área de estudio presenta potencial para dicha actividad ya que en un breve recorrido se puede observar una avifauna, y en general una biodiversidad, asociada a dos zonas biogeográficas, a varios tipos de vegetación y un importante aporte de aves migratorias, incluidas las del río de rapaces. El éxito de cualquier medida tomada en favor de la conservación de las aves de la región dependerá de la concientización y participación de sus habitantes, pues son ellos quienes viven diariamente en contacto con el medio y quienes tienen una mayor influencia en su conservación o perturbación. Así, es de suma importancia que los estudios de biodiversidad, como el presente, se realicen con el acuerdo y la contribución de los pobladores y que la información generada pueda volver a ellos de diversas formas además de los productos científicos.
Los estudios faunísticos a escala local son importantes como un método para detectar áreas de gran diversidad o de importancia para la conservación, que podrían pasar desapercibidas cuando se realizan a escalas mayores, y para reunir información sobre los procesos que ocurren a nivel de comunidades (Rojas-Soto y de Ita, 2005; Villa-Bonilla et al., 2008). El presente estudio nos permitió ejemplificar cómo se organiza la diversidad de aves una zona que presenta mucha heterogeneidad de ambientes, incluyendo zonas conservadas y otras muy perturbadas, tanto de afinidad tropical como templada. Como era de esperarse encontramos una mayor diversidad de aves en las zonas con menor perturbación y destacamos que dichas áreas también son una fuente desde donde las aves pueden desplazarse a áreas con mayor perturbación. La comparación con los registros históricos, nos permitió concluir que cuando hay una lenta modificación en el ambiente, la composición de especies de aves puede mantenerse aproximadamente estable, sin embargo, esto no descarta que pueda ocurrir un deterioro constante, en particular en la abundancia de las especies.

