











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Collecting and preserving specimens has been a crucial cornerstone in furthering our understanding of the natural world (Bradley et al., 2014; Holmes et al., 2016; Rowe et al., 2011; Winker, 2004). The benefits that scientific collecting and natural history collections provide to science and society are beyond debate (Graham et al., 2004; Suárez & Tsutsui, 2004). Even so, cutting the budgets for field-based research, scientific collecting, and maintaining and growing collections is becoming more frequent around the world (Kemp, 2015). Increasing the scientific collections is therefore a challenging issue, whose resolution will require new creativity for justifying and planning scientific collecting (Cook & Light, 2019). This is especially crucial in biodiversity-rich regions of the planet where knowledge gaps, threats to biodiversity, and funding limitations predominate (Dunnum et al., 2018).
Here, I show the use of ecological niche models and paleodistributions (or past distributions) as a framework for planning fieldwork and natural history collection growth based on spatio-temporal hypotheses. To do so, I emphasize why I consider that the usefulness of considering paleodistributions goes beyond suggesting areas that could harbor new records to increase scientific collections, by helping to identify biodiversity data gaps that can be crucial to studying evolutionary and ecological processes. This strategy would allow for well-planned growth of collections and stimulate future long-term evolutionary research.
Importantly, this contribution makes no attempt to debate the ethical aspects of scientific collecting, which have been dealt with extensively in the literature (Patterson, 2002; Sikes et al., 2011). Rather, I consider that acquiring necessary scientific knowledge about the biodiversity of our planet requires that natural history collections continue to grow through scientific collecting, and that this venture must continue to be supported by the scientific community and funding agencies (Cook & Light, 2019; Gippoliti, 2018).
Identifying data gaps and research needs
One priority of natural history museums is to capture the widest genetic and phenotypic variability in their collections as possible since an objective study of variability (e.g., genetic adaptations and phenotypic plasticity) is only possible through adequate geographic sampling (Gippoliti, 2018). An adequate geographical coverage within a scientific collection allows for a better understanding of the response of the species to past pressures such as climate change and the identification of populations that could assume more distinct ecological roles in the face of current global change (Ceballos & Ehrlich, 2009.).
However, given that 1) genetic and phenotypic variability is not distributed homogeneously throughout the distribution area of a species, and 2) biological sampling is focused toward more easily accessible areas, it is inevitable that the current collections represent only a biased sample of the variability existing in nature (Illoldi-Rangel et al., 2004; Jarnevich et al., 2015; Meineke et al., 2018; Winker, 2004; Zamudio et al., 2016). This may be the case even for species that are well-represented in terms of the number of specimens in collections. Predicting where we might find natural variability that is not yet preserved in collections would be very helpful for making decisions about what and where to collect, allowing better use of shrinking resources.
Niche modelling and paleodistributions
Ecological -correlative- Niche Modeling (ENM) estimates the current potential species distribution by identifying geographical regions with environmental conditions similar to those where the presence of the species has been confirmed (Soberón & Peterson, 2005). Among its numerous applications (for an example, see Araújo et al., 2019), ENM can be used to identify regions on the planet, mainly those that are remote and poorly sampled, that could harbor yet unknown components of biodiversity (e.g., new populations of known species, or even species that are not yet known; Raxworthy et al., 2003).
Niche models can also be projected onto Quaternary climates simulated with Global Circulation Models to estimate the potential distribution of species through time (i.e., paleodistributions; Peterson & Lieberman, 2012). For example, ENM have been used to estimate geographic paleodistributions through different epochs from the Last Glacial Maximum (ca. 21,000 years) to the present, in order to track historical species distributions under different climates (Nogués-Bravo, 2009; Peterson & Lieberman, 2012). Specifically, paleodistributions through the last glacial-interglacial cycle are relevant in evolutionary biology, since this time period has been a causal factor in current geographical distribution patterns. The abrupt alternation between hot (even warmer than the present) and cold periods is thought to have caused expansion/ contraction and fragmentation/historical connectivity events that should leave signals on the distributions of current lineages; it is therefore theoretically possible to predict the expected genetic and phenotypic variability of known or still unknown lineages (Blois et al., 2010; Chan et al., 2011; Hewitt, 2000; Zamudio et al., 2016).
Beyond new records
To illustrate the utility of paleodistribution modelling for planning the growth of natural history collections, I used the small-eared shrews of the Cryptotis mexicanus group (Mammalia, Eulipotyphla) as an example. This group is mainly distributed in the tropical montane cloud forest of Mexico (Cryptotis magnus, C. mexicanus, C. nelsoni, and C. phillipsii). It is a relatively well-known clade, with a large number of specimens in collections and with georeferenced localities (Guevara et al., 2015; Guevara & Sánchez-Cordero, 2018). Using maxent, an open source software for modeling species distributions (Phillips et al., 2017), the ecological niche of this clade was estimated to predict its potential distribution at present and at the following times of the last glacial-interglacial cycle under the CCSM3 climate scenario (Community Climate System Model; Collins et al., 2006): Late-Holocene (4.2 - 0.3 ka), Mid-Holocene (8.326 - 4.2 ka), Early-Holocene (11.7 - 8.326 ka), Younger Dryas Stadial (12.9-11.7 ka), Bølling-Allerød Interstadial (14.7-12.9 ka), Heinrich Stadial 1 (17.0-14.7 ka), and Last Glacial Maximum (ca. 21 ka) (Fig. 1).
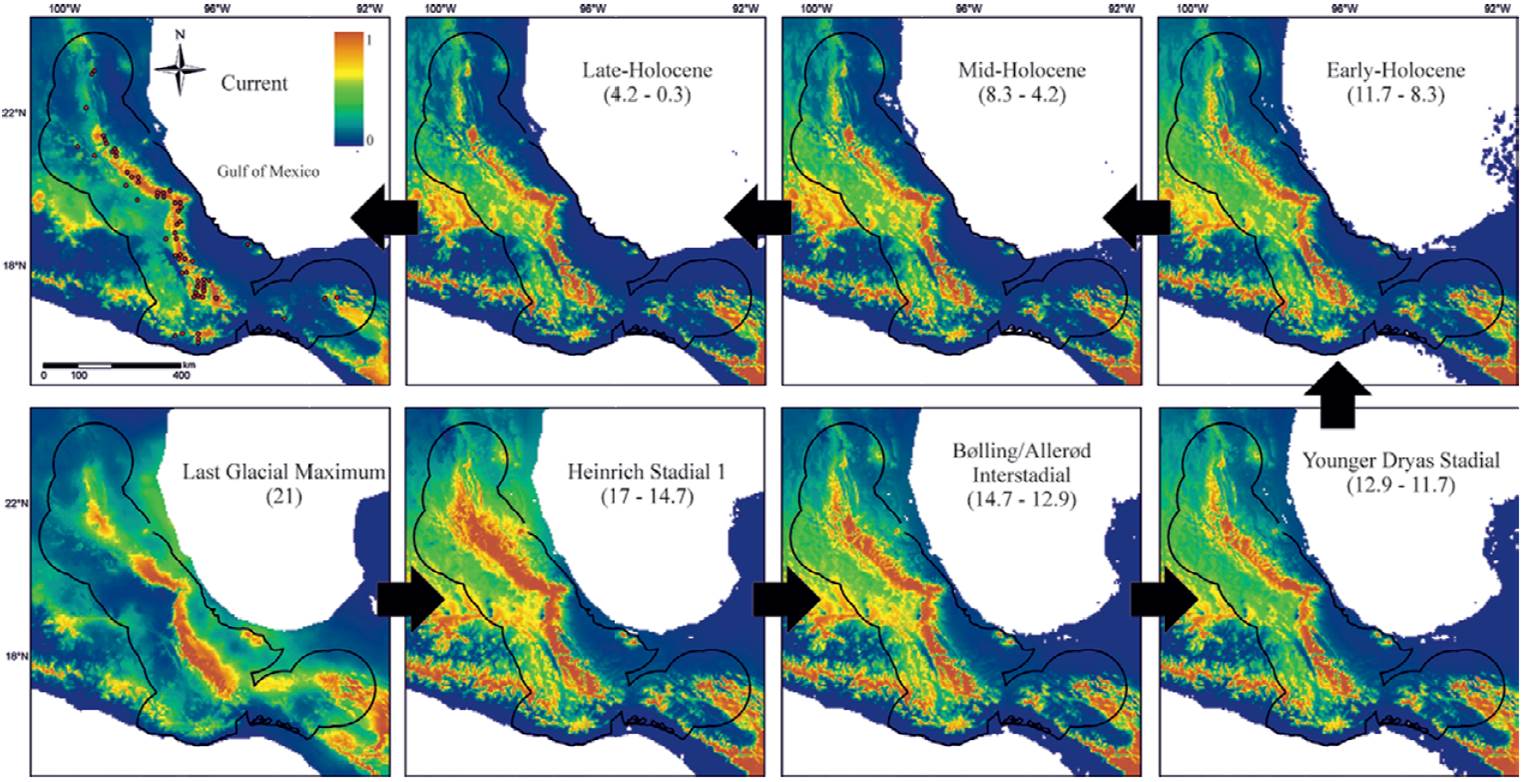
Figure 1 Maps describing the potential historical geographic distributions of the group of shrews Cryptotis mexicanus from the Last Glacial Maximum to the present. The number in parentheses indicates thousands of years before the present. The area inside the black outline is the area that has been accessible via dispersion during this time period and which was used to calibrate the ecological niche model (accessible area hypothesis according to Barve et al., 2011). The color indicates environmental suitability (redder colors are more suitable; see key in the upper left panel). The arrows indicate the sequence of events from past to present.
By superimposing the records of the Cryptotis mexicanus group that are currently available in natural history collections, it is possible not only to detect regions that could harbor new records, but also to identify if they come from areas of postglacial colonization, areas of predicted stability (refugial zones), and if they have maintained contact that could allow gene flow within this group of species (Chan et al., 2011).
As an example, I highlight 2 relatively poorly sampled regions for small mammals in Mexico (Briones-Salas & Sánchez-Cordero, 2004; Guevara & Sánchez-Cordero, 2018; Fig. 2). According tothe paleodistribution hypothesis, region 1 or Sierra de Otontepec has been climatically suitable for the C. mexicanus group through the last glacial-interglacial cycle, so it is reasonable to predict the existence of new populations. Notably, a recent expedition found 6 individuals of C. mexicanus in this region (Mayen-Zaragoza et al., 2019). Going one step beyond new records, we could also predict this population to be the result of postglacial colonization from the mountain range to the west, specifically during the Heinrich Stadial 1.
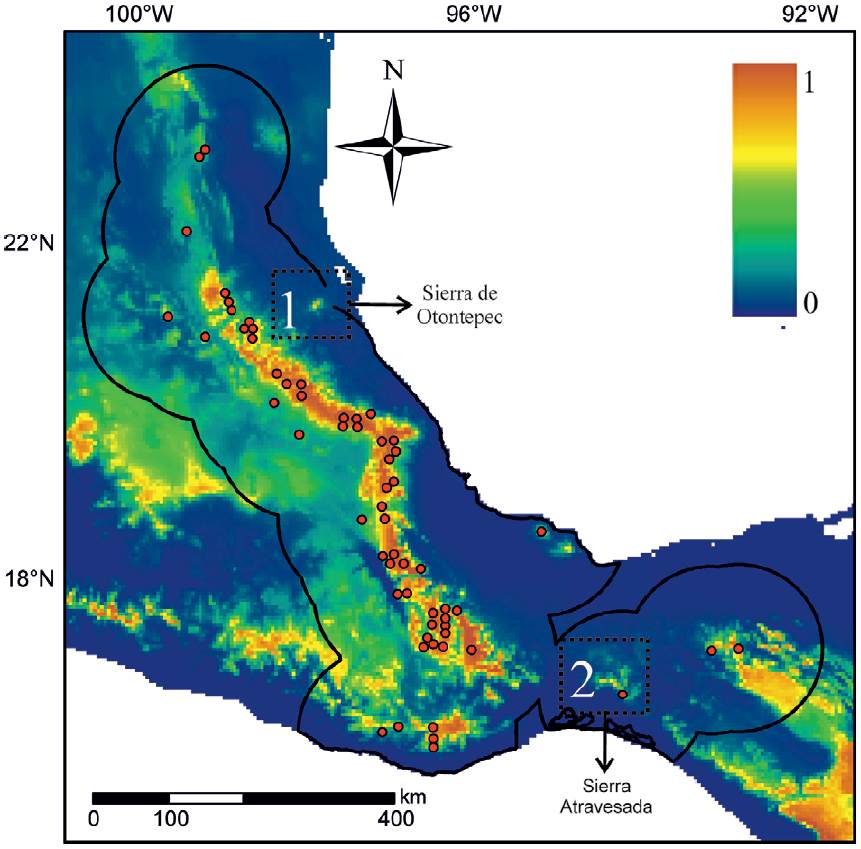
Figure 2 Current potential distribution map indicating 2 areas that could contain more records for the group of species of shrews Cryptotis mexicanus (see text for details). The red points indicate occurrence localities in biological collections. The area inside the black outline is the area that has been accessible via dispersion during this time period (accessible area hypothesis according to Barve et al., 2011). The color indicates the degree of environmental suitability (redder colors are more suitable, see key in the upper right corner of the panel). The arrows indicate the sequence of events from past to present.
Region 2 borders the "Sierra Atravesada" that lies in the middle of the Isthmus of Tehuantepec, which represents a long-standing biogeographical dilemma about its role as barrier for montane species (Peterson et al., 1999). The "Sierra Atravesada" has a record supported by a single individual collected more than 50 years ago and for which no tissue samples are available (California Academy of Science, CAS 14636). Based on the paleodistribution hypothesis, this region did not have adequate climatic conditions for the group C. mexicanus during the Last Glacial Maximum, but the populations had to move north and formed a bridge along the Isthmus, promoting gene flow between populations on either side of the Isthmus. We could then expect that this population is not highly differentiated from the populations on either side of the Isthmus. The 2 examples described here can be later tested through obtaining more specimens or samples and using genetic and morphological analyses.
I aim to illustrate using this simple example that paleodistributions have the potential to provide information about the causes or events that could explain current distributional patterns, as well as to predict patterns of genetic and phenotypic variability of still-unknown biodiversity. The framework outlined here can certainly be applied to questions on broader taxonomic and geographic scales. For example, including more species with similar ecological characteristics could provide information about the dynamics of communities over time. This example also shows that even for relatively well-known areas and taxa, there are both gaps in knowledge and opportunities for field-based research that if addressed could greatly enrich our integrated understanding of the evolution, ecology, and conservation of biodiversity.
Perspective
The challenges that humanity faces require that collections continue to grow. However, they do not just need the preservation of more specimens per se, but the preservation of more historical events that allow us to better understand complex biological processes at the species and community levels (Schindel & Cook, 2018). The framework presented in this opinion is a proposal, based on spatio-temporal hypotheses, for the kind of planning that should go into collections. As a parallel step, these specimens or samples collected following this framework will serve to continue the validation of ecological niche models and paleodistributions, a task that urgently needs to be expanded and improved (Araújo et al., 2019). I hope this opinion also serves to stimulate more integrated and cooperative long-term research among niche modelers, taxonomists, ecologists, and biogeographers, or, as Futuyma (1998) said, the creation of future scientific naturalists.

