











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Las algas bentónicas son una de las comunidades más productivas en lagos y ríos (Passy, 2008). En particular las diatomeas bentónicas han mostrado una gran eficiencia en la regulación y flujo de nutrientes (Van Luijin et al., 1995). Algunos estudios en humedales han mostrado que las comunidades bentónicas ricas en diatomeas absorben fósforo reactivo soluble (FRS) directamente desde el sedimento superficial (Ibarra et al., 2009; Novelo y Tavera, 2003). La absorción de fósforo por diatomeas bentónicas ocurre en episodios constantes y rápidos (menos de 1 hora), produciendo aumentos en la dimensión de la capa oxigenada de los sedimentos e inhibiendo la liberación de FRS (Zhang y Mei, 2015), lo que contribuye a depauperar la columna de agua. En lagos eutrofizados, como el lago Chalco (Caballero, 1995), no es sorprendente que las diatomeas más abundantes en la comunidad bentónica sean especies con estrategias de alta asimilación de nutrientes. Este rasgo podría ser un factor común de ambientes someros y eutrofizados, especialmente si reciben un impacto antropogénico, y aunque este conocimiento hasta ahora no ha sido homologado para México, algunos autores han propuesto que ocurre en regiones tropicales, como resultado de una preferencia autóctona de las diatomeas de cada localidad, sugiriendo que su análisis podría requerir la modificación de índices históricamente utilizados para evaluar riqueza y diversidad (Bellinger et al., 2006). En el manejo de nuestros datos en el lago Chalco, utilizamos los índices reconocidos en la literatura y mostramos su pertinencia para el análisis de la estructura de una comunidad como las diatomeas bentónicas. El resultado de estos análisis se confrontó con la biología y ecología de algunas de las especies más abundantes de esta comunidad, que mostró rasgos comunes con otros ecosistemas lénticos del sur de la Ciudad de México (Buendía-Flores et al., 2015). Sin embargo, la comparación con otras regiones de nuestro país está limitada porque el estudio de diatomeas que habitan en sedimentos de ambientes continentales lénticos ha recibido una atención desproporcionadamente menor en relación con su importancia a nivel global o nacional para comunidades bentónicas de lagos. No se cuenta entonces con datos históricos suficientes para identificar límites de la diversidad alfa, ni patrones de distribución de las especies, ni afinidades ecológicas de las poblaciones mexicanas.
En términos amplios, el estudio de diatomeas bentónicas de ambientes eutrofizados es importante, pues la tolerancia con la que responden ante factores de estrés permite su crecimiento sustancial, lo que las hace dominantes. Al mismo tiempo, mientras mayor sea el número de recursos, aumenta la oportunidad de que exista mayor riqueza. Por ello se considera que la comunidad en la que ocurre eso ofrece una mayor dimensionalidad de nicho (Passy, 2008). Desde luego, esta dinámica aporta argumentos medulares de sostenibilidad de los recursos acuáticos pues en ambientes con múltiples disturbios además de la eutrofización, no se conocen los límites que excluyen a más especies de las que se posibilita que existan. El estudio de las diatomeas bentónicas ofrece un alto valor en el contexto del lago Chalco pues se trata de una zona con alto disturbio urbano y agropecuario y con sobreexplotación del manto acuífero, lo que deriva en una acentuada subsidencia, generando varios problemas de tipo ambiental (Ortiz-Zamora y Ortega-Guerrero, 2007). Por otra parte, se ha comprobado que los cuerpos de agua en el Valle de México actúan como reguladores de las fluctuaciones estacionales de temperatura (Ruiz-Angulo y López-Espinoza, 2015), por lo que es necesaria su conservación, especialmente cuando se trata de lagos urbanos. Esta situación resalta el valor de estudiar a las comunidades bióticas como la que forman las diatomeas, pues el aspecto biológico de la conservación de ecosistemas acuáticos debe atenderse para mantener los servicios ecosistémicos que ofrecen, como son la regulación térmica y calidad del agua.
Los aspectos que sobresalen de esta investigación son la documentación detallada de la composición y dominancia de especies de diatomeas bentónicas y un análisis de la posible funcionalidad del ambiente basada en la diversidad y equitatividad de la comunidad y en la biología de las especies más abundantes del componente florístico, principalmente de las diatomeas centrales de talla pequeña, y que coinciden con los registrados en otras investigaciones de ambientes similares, a nivel mundial (Jewson et al., 2015; Malik y Saros, 2016; Saros y Anderson, 2015; Saros et al., 2016).
Materiales y métodos
El lago Chalco se encuentra al sureste de la cuenca de México (fig. 1) en la porción centro oriental de la Faja Volcánica Transversal Mexicana a una altura media de 2,240 m snm. La precipitación anual es de 552 mm y el potencial de evaporación es de 1,825 mm anuales (SMN, 2019); predomina un clima semiseco templado (García, 1998). Presenta una longitud máxima de 4 km, ancho máximo de 3.2 km, una línea de costa de 19.8 km y una profundidad máxima de 3 m. Está limitado al norte por los domos volcánicos de la sierra de Santa Catarina, al este por la sierra Nevada, al sur por el campo volcánico monogenético de la sierra de Chichinautzin y al oeste por la divisoria por la que corre la avenida Tláhuac-Chalco (Herrera-Hernández, 2011). Alrededor del cuerpo de agua se pueden observar 2 tipos principales de uso de suelo (fig. 1), la zona urbana constituida por las delegaciones Tláhuac (344,106 habitantes) y Xochimilco (402,056 habitantes) en la Ciudad de México y el municipio de Valle de Chalco Solidaridad (332,279 habitantes) en el Estado de México y la zona agrícola con cultivos de maíz, alfalfa, frijol y trigo. Además, algunas zonas se dedican a la ganadería (Bouvier et al., 1993).

Figura 1 Ubicación de los puntos de recolecta (*) en el lago Chalco. Modificado de www.gaia.inegi.org.mx
Actualmente, el lago recibe solo el agua de lluvia que se acumula lo suficiente para llenarlo debido a una progresiva subsidencia del terreno; ésta es ocasionada por la sobreexplotación del manto acuífero como parte del suministro a la Ciudad de México (Ortiz-Zamora y Ortega- Guerrero, 2007).
Se realizaron recolecciones de muestras de agua en los meses de marzo a agosto, noviembre a diciembre de 2015 y de enero a octubre de 2016 con una frecuencia mensual, en 3 puntos geoposicionados. Se tomaron muestras de agua para determinar nutrimentos como fósforo reactivo (ortofosfato) y fósforo total y las formas de nitrógeno inorgánico disuelto (NID expresado en mg L-1, como suma de nitrato, nitrito y amonio) con métodos espectrofotométricos (Hach DR-1900, Loveland CO, EUA), con base en los estándares aprobados para el análisis de agua dulce de la Agencia de Protección Ambiental de los Estados Unidos (USEPA, 2012). Se midieron algunos factores fisicoquímicos (temperatura, pH, conductividad K25, oxígeno disuelto, clorofila-a) con una sonda multiparamétrica Hydrolab D-5 (Danaher Corp., Loveland CO, EUA). Para muestrear la comunidad bentónica de diatomeas, en cada punto se tomó sedimento con una draga Eckman (Analab, S.A. Barna, España), de la cual se recolectó solo la parte más superficial (≈ 2 mm de grosor de capa) con una espátula. Las muestras se mantuvieron en frío (≈ 4-6 °C) hasta su procesamiento en el laboratorio. Se tomaron 0.5 gr de sedimento liofilizado (Labconco Corporation FreeZoneTM, Kansas City, MO, EUA) y se sometieron a un proceso de digestión lenta de acuerdo con Stroemer et al. (1995), conservando la muestra final en 2.0 ml de agua destilada. Agitando suavemente, se tomaron 200 µl de la muestra por cada preparación montada con resina Naphrax® (Chippenham, RU). Para la documentación fotográfica y conteo de valvas se utilizó un microscopio Nikon-Optiphot (Nikon Co.) equipado con una cámara Nikon Coolpix S10 (Nikon Co., Tokio, Japón). El cálculo de la abundancia de cada especie sobre un conteo mínimo de 300 valvas se expresó como número de valvas por gramo de sedimento seco (gss), dividido por 2 para considerar frústulas en el cálculo final, de acuerdo con Scherer (1994). El cálculo de biomasa como peso fresco expresado en μm3 gss se obtuvo sobre las frústulas contadas, de acuerdo con Hillebrand et al. (1999) y se aplicó a las especies dominantes con el ajuste morfométrico correspondiente (formas geométricas más cercanas). La mitad de volumen del material limpio (1.0 mL) se sometió a un proceso de flotación con metatugstato de litio, de acuerdo con Morley et al. (2004), como paso previo a la preparación para su documentación en un microscopio electrónico de barrido Jeol JSM-5310LV, (JEOL USA, Inc., Peabody, MA, EUA) a 20 o 25 kV. El material obtenido se incorporó a la colección EMexChal, de FCME (Herbario de la Facultad de Ciencias, UNAM). La determinación taxonómica y la búsqueda de afinidades ecológicas de cada especie se realizó con base en literatura especializada: Krammer (1997a, b, 2000); Krammer y Lange-Bertalot (1985, 1986, 1988, 1991a, b); Lange-Bertalot (1980); López-Fuerte y Siqueiros- Beltrones (2006); Patrick y Reimer (1966, 1975). La clasificación taxonómica se basó en Medlin y Kaczmarska (2004) confrontada con Guiry y Guiry (2019); INA (2019) y Novelo (2012).
Los análisis para determinar la estructura de la comunidad de diatomeas bentónicas incluyeron la determinación del esfuerzo de muestreo (rarefacción) por medio de curvas de acumulación de especies (Kraker-Castañeda y Cóbar-Carranza, 2011) con el programa StatSoft Statistica. Versión 6.0 (2001). Siguiendo a Ávalos (2007), la curva de acumulación fue calculada de acuerdo con el modelo paramétrico de Clench para ajustar la asíntota. Se estimó la diversidad de especies con el índice de Shannon-Wiener (H´) y el valor de dominancia con el índice de Simpson (D) para detectar procesos empobrecedores de la comunidad (Magurran, 1996); la estimación de la equitatividad de la comunidad se realizó con el índice de equidad de Pielou (J´). Los cálculos de los índices se hicieron en el software PAST versión 3.17 (Hammer et al., 2001). Los resultados obtenidos se evaluaron independientemente para 2015 y 2016.
Resultados
Nutrientes y factores abióticos. La concentración de fósforo total estimada fue en promedio 0.43 mg L-1 en 2015 y de 1.1 mg L-1 en 2016; la concentración de clorofila-a promedio fue 75.6 μg L-1 en 2015 y de 102.5 μg L-1 en 2016. La concentración de fósforo total asumiendo la variación, indicó un lago eutrófico a hipertrófico; este resultado no fue consecuente con la concentración de clorofila-a, que mostró una columna oligotrófica (Lampert y Sommer, 2007). La concentración promedio de nitrógeno inorgánico disuelto total (NID) fue de 1.0 mg L-1 en ambos años de estudio. Se determinó una columna homogénea de agua tibia (temperatura promedio anual de 21.7 °C en 2015 y 20.8 °C en 2016). Los valores de conductividad fueron en promedio de 1,567 μS cm-1 (K25) en 2015 y 1,961 Μs cm-1 (K25) en 2016. Los valores de pH tanto en 2015 (9.6) como en 2016 (10.0) caracterizaron una columna de agua con basicidad importante, lo que se correspondió también con los valores registrados de alcalinidad de carbonatos, 350.2 mg L-1 en 2015 y 439.0 mg L-1 en 2016.
Se presentan las características morfológicas, comentarios taxonómicos y algunos comentarios ecológicos de las especies que constituyen registros nuevos para México, incluyendo la documentación de herbario y se ilustran los 70 taxones de diatomeas del lago Chalco, refiriendo la presencia particular en cada año o en ambos años de estudio (tabla 1; figs. 2-98). Tres taxones fueron registros nuevos: Thalassiosira aff. duostra que corresponde también con el primer registro para América; Nitzschia graciliformis y Epithemia argus var. alpestris.
Tabla 1 Listado y presencia de especies exclusivas en 2015, en 2016 y compartidas.
| Bacillariophyta | 2015 | 2016 |
|---|---|---|
| Mediophyceae | ||
| Thalassiosirales, Stephanodiscaceae | ||
| Cyclotella meneghiniana Kützing, 1844 | ✓ | ✓ |
| Discostella pseudostelligera (Hustedt) Houk et Klee, 2004 | ✓ | ✓ |
| Thalassiosirales, Thalassiosiraceae | ||
| Thalassiosira aff. duostra Pienaar et Pieterse, 1990 | ✓ | ✓ |
| Bacillariophyceae | ||
| Fragilariales, Fragilariaceae | ||
| Fragilaria capucina Desmazières, 1830 | ✓ | ✓ |
| Fragilariforma virescens (Ralfs) Williams et Round, 1988 | ✓ | ✓ |
| Pseudostaurosira brevistriata (Grunow) Williams et Round, 1987 | ✓ | ✓ |
| P. subsalina (Hustedt) E.A. Morales, 2005 | ✓ | ✓ |
| Staurosira construens Ehrenberg, 1843 | ✓ | ✓ |
| S. neoproducta (Lange-Bertalot) Chudaev et Gololobova, 2012 | ✓ | ✓ |
| S. venter Cleve et J.D.Möller, 1879 | ✓ | ✓ |
| Punctastriata mimetica E.A. Morales, 2005 | ✓ | ✓ |
| Tabularia fasciculata (C. Agardh) D.M. Williams et Round, 1986 | ✓ | ✓ |
| Achnanthales, Achnanthidiaceae | ||
| Achananthidium exiguum (Grunow) Czarnecki, 1994 | ✓ | ✓ |
| Achnanthidium minutissimum (Kützing) Czarnecki, 1994 | ✓ | ✓ |
| Lemnicola hungarica (Grunow) F.E. Round et P.W. Basson, 1997 | ✓ | ✓ |
| Achnanthales, Cocconeidaceae | ||
| Cocconeis placentula Ehrenberg, 1838 | ✓ | ✓ |
| Cymbellales, Anomoeoneidaceae | ||
| Anomoeoneis costata (Kützing) Hustedt, 1927 | ✓ | ✓ |
| A. sphaerophora E. Pfitzer, 1871 | ✓ | ✓ |
| Cymbellales, Cymbellaceae | ||
| Cymbella aspera (Ehrenberg) Cleve, 1894 | ✓ | ✓ |
| C. mexicana (Ehrenberg) Cleve, 1894 | ✓ | ✓ |
| Encyonema mesianum (Cholnoky) D.G. Mann, 1990 | ✓ | ✓ |
| E. silesiacum (Bleisch) D.G. Mann, 1990 | ✓ | ✓ |
| Navicymbulla pusilla (Grunow) Krammer, 2003 | ✓ | ✓ |
| Cymbellales, Gomphonemataceae | ||
| Gomphonema affine Kützing, 1844 | ✓ | ✓ |
| G. angustatum (Kützing) Rabenhorst, 1864 | ✓ | |
| G. clavatum Ehrenberg, 1832 | ✓ | |
| G. gracile Ehrenberg, 1838 | ✓ | ✓ |
| G. hebridense W. Gregory, 1854 | ✓ | ✓ |
| G. parvulum (Kützing) Kützing, 1849 | ✓ | ✓ |
| G. truncatum Ehrenberg, 1832 | ✓ | ✓ |
| Cymbellales, Rhoicospheniaceae | ||
| Rhoicosphenia abreviata (C.Agardh) Lange-Bertalot, 1980 | ✓ | ✓ |
| Naviculales, Diadesmidaceae | ||
| Diadesmis confervacea Kützing, 1844 | ✓ | ✓ |
| Luticola goeppertiana (Bleisch) D.G. Mann, 1990 | ✓ | ✓ |
| L. mutica (Kützing) D.G. Mann, 1990 | ✓ | ✓ |
| Naviculales, Diploneidaceae | ||
| Diploneis pseudovalis Hustedt, 1930 | ✓ | ✓ |
| Naviculales, Naviculaceae | ||
| Eolimna subminuscula (Manguin) G. Moser, Lange-Bertalot et Metzeltin, 1998 | ✓ | ✓ |
| Geissleria tagensis Novais et Ector, 2013 | ✓ | ✓ |
| Hippodonta hungarica (Grunow) Lange-Bertalot, Metzeltin et Witkowski, 1996 | ✓ | ✓ |
| Navicula cryptocephala Kützing, 1844 | ✓ | ✓ |
| N. veneta Kützing, 1844 | ✓ | ✓ |
| Naviculales, Pinnulariaceae | ||
| Pinnularia abaujensis var. linearis R.M. Patrick, 1966 | ✓ | ✓ |
| Naviculales, Sellaphoraceae | ||
| Fallacia pygmaea (Kützing) A.J. Stickle et D.G. Mann, 1990 | ✓ | ✓ |
| Sellaphora laevissima (Kützing) D.G. Mann, 1989 | ✓ | ✓ |
| S. pseudoventralis (Hustedt) Chudaev et Gololobova, 2015 | ✓ | ✓ |
| S. pupula (Kützing) Mereschkovsky, 1902 | ✓ | ✓ |
| Naviculales, Stauroneidaceae | ||
| Craticula halophila (Grunow) D.G. Mann, 1990 | ✓ | ✓ |
| Stauroneis phoenicenteron (Nitzsch) Ehrenberg, 1843 | ✓ | |
| Thalassiophysales, Catenulaceae | ||
| Amphora copulata (Kützing) Schoeman et R.E.M. Archibald, 1986 | ✓ | ✓ |
| Halamphora coffeiformis (Agardh) Levkov, 2009 | ✓ | ✓ |
| H. veneta (Kützing) Levkov, 2009 | ✓ | ✓ |
| Bacillariales, Bacillariaceae | ||
| Denticula valida (Pedicino) Grunow, 1882-1885 | ✓ | ✓ |
| Hantzschia amphioxys (Ehrenberg) Grunow, 1880 | ✓ | ✓ |
| Nitzschia amphibia Grunow, 1862 | ✓ | ✓ |
| N. communis Rabenhorst, 1848-1860 | ✓ | ✓ |
| N. frustulum (Kützing) Grunow, 1880 | ✓ | ✓ |
| N. gandersheimiensis var. tenuirostris Grunow, 1881 | ✓ | ✓ |
| N. graciliformis Lange-Bertalot et Simonsen, 1978 | ✓ | ✓ |
| N. hantzschiana Rabenhorst, 1860 | ✓ | ✓ |
| N. palea (Kützing) W. Smith, 1856 | ✓ | ✓ |
| N. umbonata (Ehrenberg) Lange-Bertalot, 1978 | ✓ | ✓ |
| Tryblionella apiculata W. Gregory, 1857 | ✓ | |
| Rhopalodiales, Rhopalodiaceae | ||
| Epithemia adnata (Kützing) Brébisson, 1838 | ✓ | ✓ |
| E. argus (Ehrenberg) Kützing, 1844 | ✓ | ✓ |
| E. argus var. alpestris (W. Smith) Grunow, 1860 | ✓ | ✓ |
| E. sorex Kützing, 1844 | ✓ | ✓ |
| E. turgida (Ehrenberg) Kützing, 1844 | ✓ | ✓ |
| E. turgida var. granulata (Ehrenberg) Brun, 1880 | ✓ | ✓ |
| Rhopalodia gibba (Ehrenberg) O.F. Müller, 1895 | ✓ | ✓ |
| R. operculata (Agardh) Håkasson, 1979 | ✓ | ✓ |

Figuras 2-24 2) Cyclotella meneghiniana; 3) Discostella pseudostelligera; 4, 5) Thalassiosira aff. duostra; 6) Fragilaria capucina; 7) Fragilaria sp. 8) Fragilariforma virescens; 9) Pseudostaurosira brevistriata; 10-12) Pseudostaurosira subsalina; 13) Staurosira construens; 14) Staurosira venter; 15) Staurosira neoproducta; 16, 17) Punctastriata mimetica; 18) Tabularia fasciculata; 19) Achnanthidium exiguum; 20) Achnanthidium minutissimum; 21) Lemicola hungarica; 22, 23) Cocconeis placentula; 24) Anomoeoneis costata. Barra = 10 μm.

Figuras 25-35 25) Anomoeoneis sphaerophora; 26) Cymbella mexicana; 27) Cymbella aspera; 28) Encyonema mesianum; 29) Encyonema silesiacum; 30) Navicymbula pusilla; 31) Gomphonema affine; 32) Gomphonema angustatum; 33) Gomphonema clavatum; 34) Gomphonema gracile; 35) Gomphonema hebridense. Barra = 10 μm; 26 y 27) = 25 μm.

Figuras 36-53 36) Gomphonema parvulum; 37) Gomphonema truncatum; 38-39) Rhoicosphenia abbreviata; 40) Diadesmis confervacea; 41) Luticola goeppertiana; 42) Luticola mutica; 43) Diploneis pseudovalis; 44) Eolimna subminuscula; 45) Geissleria tagensis; 46) Hippodonta hungarica; 47) Navicula cryptocephala; 48) Navicula veneta; 49) Pinnularia abaujensis var. linnearis; 50) Fallacia pygmaea; 51) Sellaphora pseudoventralis; 52) Sellaphora laevissima; 53) Sellaphora pupula. Barra = 10 μm.

Figuras 54-70 54) Craticula halophila; 55) Stauroneis phoenicenteron; 56) Amphora copulata; 57) Halamphora coffeiformis; 58) Halamphora veneta; 59, 60) Denticula valida; 61) Hantzschia amphyoxis; 62) Nitzschia amphibia; 63) Nitzschia communis; 64, 65) Nitzschia frustulum; 66) Nitzschia graciliformis; 67) Nitzschia palea; 68) Nitzschia gandersheimiensis var. tenuirostris; 69) Nitzschia hantzschiana; 70) Nitzschia umbonata. Barra = 10 μm.

Figuras 71-79 71) Tryblionella apiculata; 72) Epithemia adnata; 73) Epithemia argus; 74) Epithemia argus var. alpestris; 75) Epithemia sorex; 76) Epithemia turgida; 77) Epithemia turgida var. granulata; 78) Rhopalodia gibba; 79) Rhopalodia operculata. Barra = 10 μm.

Figuras 80-88 Fotografías de MEB por método de electrones secundarios. 80-83) Cyclotella meneghiniana; 84) Thalassiosira aff. duostra; 85) Pseudostaurosira brevistriata; 86) Staurosira construens; 87) Staurosira venter; 88) Punctastriata mimetica. Barras = 1 μm.
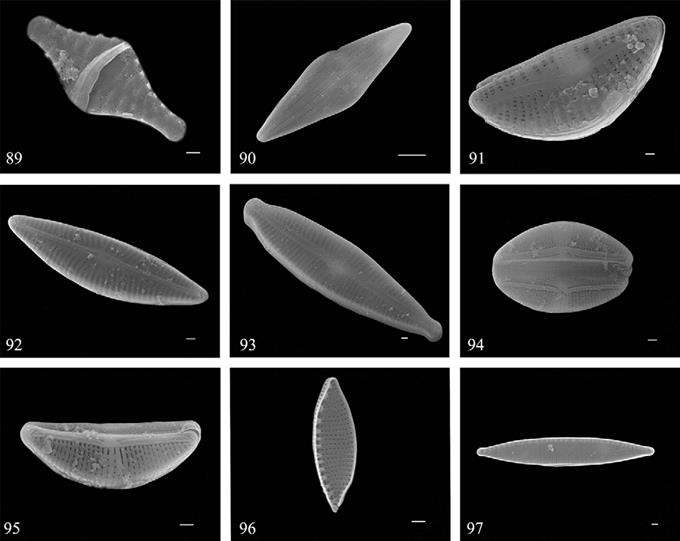
Figuras 89-97 Fotografías de MEB por método de electrones secundarios. 89) Punctastriata mimetica; 90) Anomoeoneis costata; 91) Encyonema mesianum; 92) Navicymbulla pusilla; 93) Gomphonema parvulum; 94, 95) Halamphora veneta; 96) Nitzschia frustulum; 97) Nitzschia palea. Barras = 1 μm.
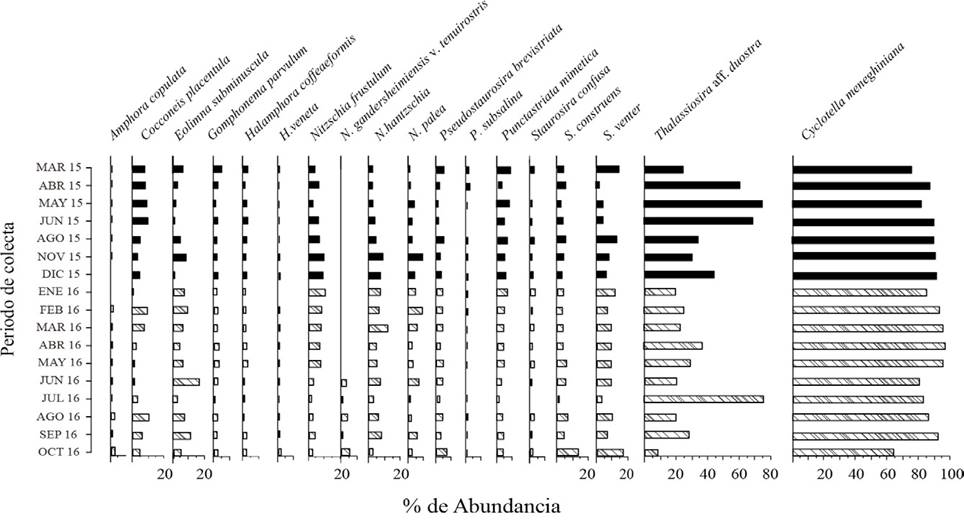
Figura 98 Porcentaje de abundancia (frústulas totales) de diatomeas bentónicas dominantes del lago Chalco durante 2015 y 2016. Las abundancias de las diatomeas dominantes fueron ≥ 2% del componente bentónico total. Los datos de Thalassiosira aff. duostra y Cyclotella meneghiniana son gráficas independientes para facilitar una visualización comparativa.
Familia: Thalassiosiraceae
Thalassiosira aff. duostra Pienaar y Pieterse 1990; figuras 4-5, 84
Diatom Res. 5: 106, figs. 1-11. 1990.
Valvas circulares con la cara valvar plana. Estrías no agrupadas en fascículos. Areolas grandes, loculadas cubiertas exteriormente por foramina, cubiertas internamente por una cribra. En la cara valvar se presentan 28 a 33 fultopórtulas marginales como proyecciones alargadas y 4 fultopórtulas, distantes entre sí, aproximadamente a la mitad entre el centro y el margen de la valva. Las poblaciones observadas se consideraron afines a la especie T. duostra porque las areolas difieren en lo siguiente: son circulares solo en el centro y hasta 2/3 de la superficie de la valva y alargadas e irregulares en el margen y en la vista conectiva; además, no mostraron ningún arreglo radial evidente. Diámetro de la valva: 13.8-16.0 µm; densidad de areolas: 20-25/10 µm. Esta especie se presume habitante de aguas contaminadas, pero su ecología no está claramente documentada. Referencias de herbario: CDMXChl 6-CDMXChl 8, CDMXChl 12-CDMXChl 14, CDMXChl 19-CDMXChl 21, CDMXChl 26-CDMXChl 28, CDMXChl 33-CDMXChl 35, CDMXChl 42-CDMXChl 44, CDMXChl 47-CDMXChl 49, CDMXChl 52-CDMXChl 54, CDMXChl 58-CDMXChl 60, CDMXChl 63-CDMXChl 65, CDMXChl 73-CDMXChl 75, CDMXChl 76-CDMXChl 78, CDMXChl 79-CDMXChl 81, CDMXChl 83-CDMXChl 85, CDMXChl 94-CDMXChl 96, CDMXChl 99-CDMXChl 101, CDMXChl 103-CDMXChl 105.
Familia: Bacillariaceae
Nitzschia graciliformis Lange-Bertalot y Simonsen 1978; figura 66
Bacillaria 1: 33.1978.
Valvas linear lanceoladas con una constricción ligera en el centro de la valva. Ápices alargados estrechamente subcapitados y redondeados. El rafe es marginal con fíbulas cortas, rectangulares, equidistantes. Las estrías no son visibles en microscopia de luz. Largo de la valva: 38.0-65.5µm; ancho de la valva: 3.3µm; densidad de fíbulas: 20-22/10µm. Ha sido registrada como perifítica y epipélica en estanques, observada en ambientes eutróficos, subsalinos, alcalífila, β-mesosapróbica. Referencias de herbario: CDMXChl 19, CDMXChl 21, CDMXChl 26, CDMXChl 27, CDMXChl 59, CDMXChl 73, CDMXChl 75, CDMXChl 76, CDMXChl 83, CDMXChl 100.
Familia: Rhopalodiaceae
Epithemia argus var. alpestris (W. Smith) Grunow 1860; figura 74
Verh. K. Zool.-Bot. Ges. Wien 10: 507. 1860
En vista valvar. Valvas dorsiventrales, estrechas, con el margen ventral cóncavo y el dorsal convexo. Esta variedad es diferente a la nominal por sus ápices redondeados y una ligera apariencia curvada hacia el lado dorsal. El canal del rafe se observa alineado al margen ventral y el arco central se extiende hasta casi llegar al margen dorsal. Costillas y estrías ligeramente radiadas. Largo de la valva: 40.0-44.8 µm; ancho de la valva: 7.3 µm; densidad de costillas: 5/10 µm; densidad de estrías entre costillas: 3-4; densidad de estrías: 11-13/10 µm. Se ha observado en arroyos, manantiales, lagos, estanques, y pantanos en tierras de cultivo, con un pH 7.3-8.6. Referencia de herbario: CDMXChl 95.
Análisis ecológico de las especies. A lo largo del estudio destacó la presencia constante de un grupo de 18 especies que reunieron abundancias con un mínimo de 13,560 frústulas totales/ggs en 2015 y 62,2458 frústulas totales/gss en 2016 (fig. 98). Esta diferencia en abundancia fue mayor a 45 veces, lo que justificó los análisis separados para cada año de estudio. En este conjunto de especies sobresalió Cyclotella meneghiniana que presentó más de 95% de la biomasa total (fig. 99 A, B). Solo Thalassiosira aff. duostra destacó con una biomasa total de 2-3%, como co- dominante de C. meneghiniana junto con el resto del grupo integrado por Amphora copulata, Cocconeis placentula, Eolimna subminuscula, Gomphonema parvulum, Halamphora coffeiformis, H. veneta, Nitzschia frustulum, N. gandersheimiensis v. tenuirostris, N. hantzschiana, N. palea, Pseudostaurosira brevistriata, Punctastriata mimetica, Staurosira neoproducta, S. construens y S. venter, todas con una biomasa total < 1%. El panorama presentado por la biomasa de estas especies fue similar al observado en la abundancia (figs. 98, 99 A, B).
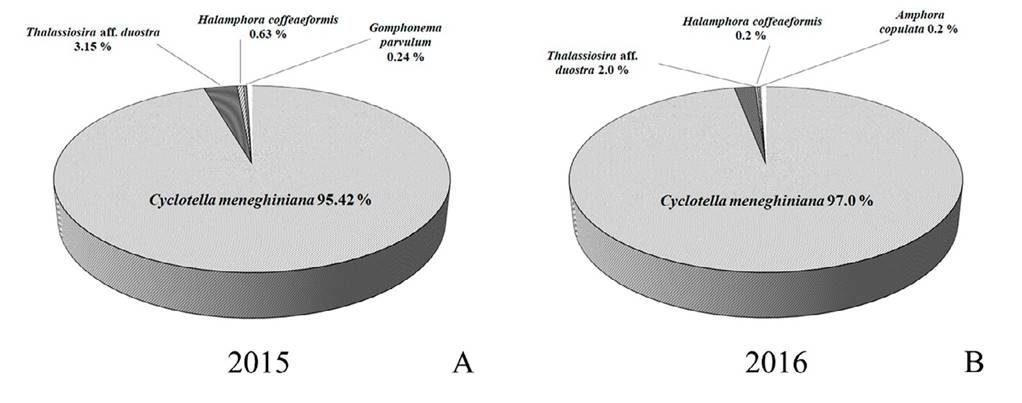
Figura 99 Porcentaje de biomasa obtenida como peso fresco (μm3 gss) para el total de diatomeas dominantes. A, 2015; B, 2016. Se destacan las especies cuya biomasa como peso fresco fue > 0.2%
El modelo de Clench mostró una estimación de 87% en 2015 (R2 = 0.99) y 93% en 2016 (R2= 0.98), por lo que las curvas asintóticas de acumulación de especies (rarefacción) (fig. 100 A, B), calculadas para los 2 años de recolecciones, comprobaron que el esfuerzo de muestreo fue suficiente, lo que respaldó el cálculo de la diversidad de Shannon-Wiener (H´), que mostró valores bajos (fig. 101); en 2015 fue H´= 2.08 (Hmax = 4.1), mayor que en 2016, H´= 0.5 (Hmax = 4.2); esto indicó, además, una tendencia evidente a su disminución con respecto al tiempo.
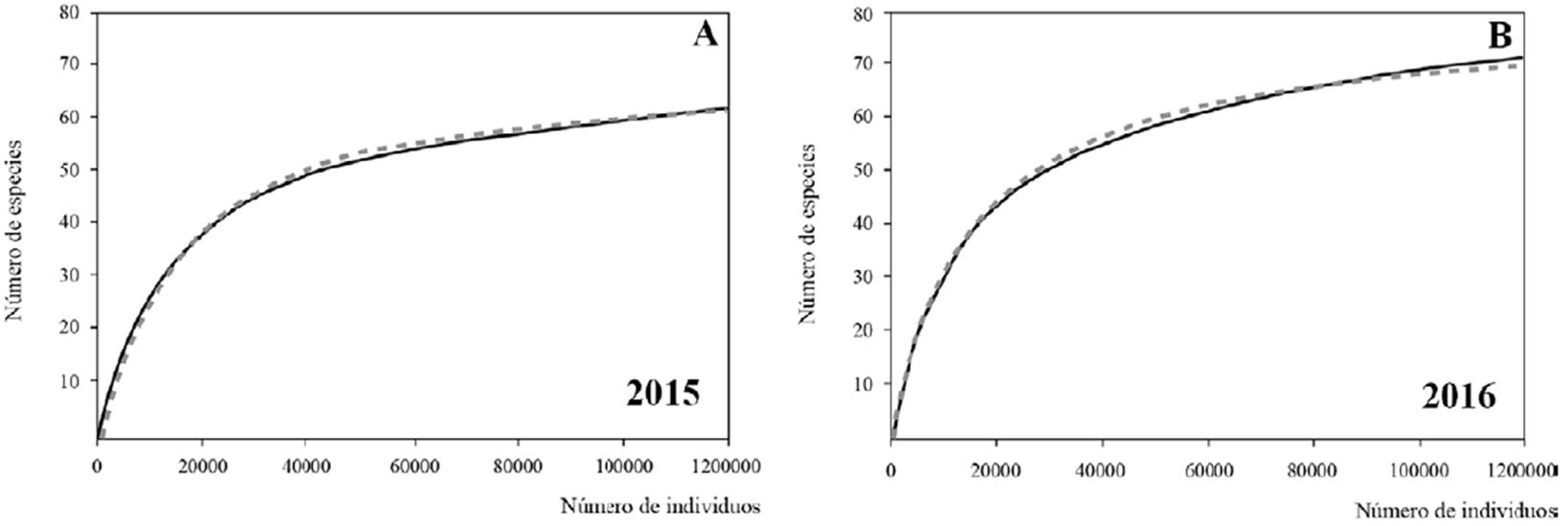
Figura 100 Curvas de rarefacción observadas (línea continua) y esperadas (línea punteada) de máximo número de frústulas (individuos). A, 61 especies en 2015; B, 70 especies en 2016.
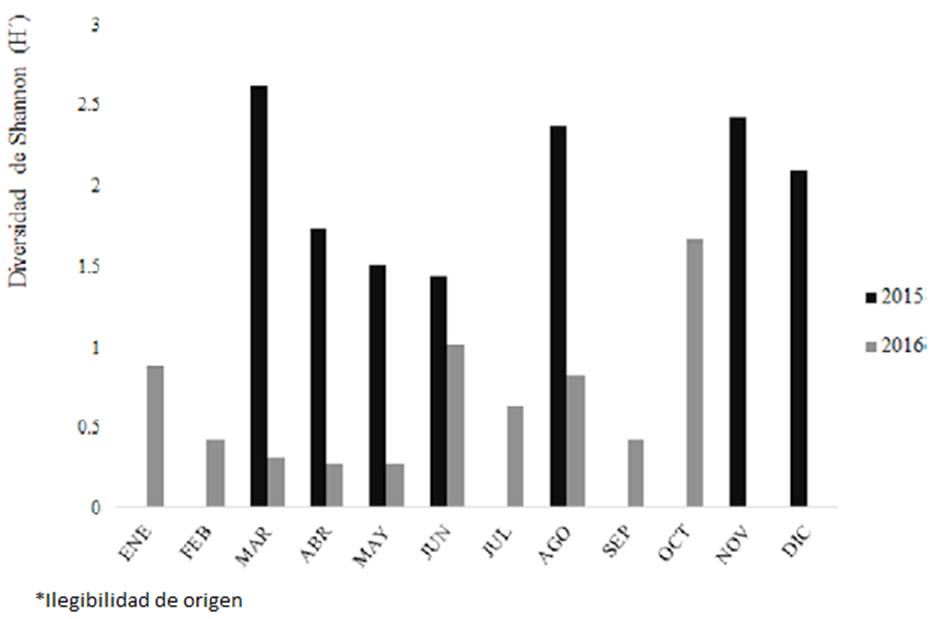
Figura 101 Valores obtenidos a partir del índice de Shannon-Wiener para la diversidad mensual en 2015 y 2016.
El índice de Simpson (D) mostró menor dominancia en 2015 (D = 0.3), que en 2016 (D = 0.8), lo que se correspondió con Shannon y con el valor del índice de Pielou (J´), que mostró mayor equitatividad en 2015 (0.5) que en 2016 (0.1) (fig. 102).

Discusión
Las concentraciones de nutrientes asimilables (NID y fósforo reactivo soluble) en la columna de agua, se mantuvieron en una proporción baja con respecto al óptimo estequiométrico considerado en la literatura (Falkowski, 2000) que, en promedio, en el lago Chalco fue de 10 veces. Esto sugiere que el fitobentos podría controlar una proporción importante de los nutrimentos hacia la columna de agua, lo que de acuerdo con la literatura podría influir en la densidad del fitoplancton por desbalance entre nutrientes (Zhang y Mei, 2015).
El conjunto de diatomeas bentónicas dominantes del lago Chalco se caracterizó por presentar especies con altos requerimientos de nitrógeno, que habitan en pH básico y comunes en cuerpos de agua con desechos sanitarios o agrícolas, por lo que se consideran tolerantes a la contaminación. Durante 2016, también se observaron especies cuyas preferencias ecológicas incluyen ambientes alcalinos, eurihalobios, de alta evaporación y elevada conductividad (Van Dam et al., 1994). Estos cambios en la composición de especies, con un abundante desarrollo en el lago Chalco sugieren una relación con la variación observada en las concentraciones de sales, que podría ser producto de fluctuaciones en el nivel de agua. Esta situación se facilita en la región porque predomina un clima semiseco, así que la composición florística y la abundancia poblacional de diatomeas dominantes de este lago sería una característica típica asociada al clima actual del valle de México, que se ha modificado fuertemente en los últimos 700 años (Ruiz-Angulo y López-Espinoza, 2015). De acuerdo con Caballero (1995), la composición de especies de diatomeas bentónicas que se registró hace 20 años tuvo un componente importante (47%) de las especies dominantes que observamos en este estudio, cuando el lago Chalco tenía un valor de precipitación anual de 662 mm y un potencial de evaporación de 1,695 mm anuales, es decir, valores similares a los que se registran actualmente, lo que implica que en Chalco se mantiene un balance negativo de precipitación-evaporación. Es notable que estos datos revelen una relación entre la composición de especies de diatomeas y las condiciones climáticas.
Una característica particular de la comunidad bentónica estudiada en Chalco es la presencia de especies de diatomeas centrales como Cyclotella meneghiniana y Thalassiosira aff. duostra, así como de Discostella pseudostelligera, una especie rara (biomasa < 0.2%) que se observó en los conteos de 2016 (tabla 1). Estas especies de diatomeas centrales en las que destacó la dominancia de C. meneghiniana, han sido recientemente estudiadas en relación con el cambio climático (Malik y Saros, 2016) y se ha visto que, independientemente de su tamaño, no necesariamente son habitantes del fitoplancton. Los cambios en temperatura, así como en profundidad de mezcla de la columna de agua afectan tanto la posición de varias especies de estos géneros como su tasa de división celular, por lo que influyen en su dominancia, pero la variación y selección de tamaño depende también de respuestas específicas a luz y nutrientes (Jewson et al., 2015; Saros et al., 2016; Vilmi et al., 2015), que sería el caso de C. meneghiniana y D. pseudostelligera. En el lago Chalco tenemos una columna de agua tibia, somera y eutrófica y C. meneghiniana y T. duostra han sido registradas como habitantes del perifiton y del bentos en regiones templadas (Altuner, 2017; Szabó et al., 2004); en México existen registros de perifiton y de bentos en los que C. meneghiniana y D. stelligera crecen con ecología similar a D. pseudostelligera (Buendía-Flores et al., 2015; García-Meza, 1999; López-Fuerte, 2010). Asimismo, C. meneghiniana ha sido reportada como afín a lagos salinos y eutróficos (Bolgovics et al., 2017), algunos con contaminación orgánica (Bere, 2014); esta especie también ha sido registrada como acalífila y halófila, dominante en la cuenca del antiguo lago Chalco (Ortega-Guerrero et al., 2017), lo que coincide con la situación ambiental que hemos observado. T. duostra se presume habitante de aguas contaminadas (Szabó et al., 2004), pero desde su descripción en ríos de África (Pienaar y Pieterse, 1990), no se ha documentado en detalle su ecología y tampoco ha sido registrada con abundancia notable. De acuerdo con algunos autores (Malik y Saros, 2016; Saros y Anderson, 2015), la presencia de diatomeas centrales sugiere la importancia de descifrar su autoecología para interpretar la distribución regional que presentan y su asociación con modificaciones climáticas a gran escala, pero sus comportamientos poblacionales no son unidireccionales ni sincrónicos, aún dentro de una misma región.
En el lago Chalco en particular, es evidente el contraste de la dominancia de Cyclotella meneghiniana con el grupo de especies más abundantes (fig. 98). Esta disparidad corresponde con los valores de dominancia y equitatividad de la comunidad (fig. 102 A, B). Asimismo, también es consecuente con los valores bajos de la diversidad (H’). La baja diversidad de una comunidad se considera un resultado de la respuesta resiliente de especies, en situaciones ambientales de disturbio frecuente (Lengyel et al., 2014). Ciertamente, la diversidad de diatomeas bentónicas de Chalco es baja en contraste con la que se ha registrado en otros estudios (López-Fuerte y Siqueiros- Beltrones, 2006; Pérez et al., 2013; Vázquez y Caballero, 2013), con valores entre H’ = 2 y H’ = 3. Si acaso este pequeño grupo de especies dominantes y codominantes reflejaran una respuesta resiliente, sería posible asociarla con estados de equilibrio del lago porque cumple con los paradigmas de estabilidad y equilibrio establecidos para el fitoplancton (Komárková y Tavera, 2003; López-Mendoza et al., 2015; Naselli-Flores et al., 2003; Sommer, 1993). Sin embargo, algunos autores consideran que, desde el punto de vista de la biomasa el panorama es diferente porque el número de individuos puede ser elevado, pero por su morfometría, este número puede corresponder con una biomasa pequeña (Hillebrand et al., 1999). Si una sola especie tiene la mayor contribución a la biomasa total de una comunidad (80% mínimo), como ocurre en el caso de Chalco con C. meneghiniana (fig. 99 A, B), nos encontramos ante un sistema altamente competitivo, que implica períodos de estabilidad muy cortos, por lo que no debe considerarse un sistema en estado de verdadero equilibrio (Leitão et al., 2003). Entonces el concepto de resiliencia no aplica adecuadamente para Chalco. Si bien la composición y la biomasa de las diatomeas bentónicas en Chalco indican una relación entre disturbio del ambiente y estados efímeros de equilibrio, no reflejan cambios rápidos de las condiciones ambientales, que podrían ser entonces en lapsos prolongados (O’Farrell et al., 2003). Por tanto, las diatomeas bentónicas estudiadas señalan una tendencia sostenida hacia la disminución de la diversidad en el lago Chalco, indicando que hay respuestas de cierto ajuste de las especies dominantes a las condiciones de alteración urbana y agropecuaria del sitio, que son continuas. Es importante añadir este resultado al conocimiento de la autoecología de diatomeas centrales en México, para entender los cambios climáticos a niveles regionales y aun cuando faltan elementos para explicar completamente la ecología de las diatomeas bentónicas del lago Chalco, en este estudio concluimos que existe una correspondencia entre la composición de especies y el ecosistema, que nos sugiere que las variaciones de la comunidad (diversidad, dominancia y equidad de especies) podrían ser fluctuantes y episódicas y que al menos las diatomeas dominantes exhiben respuestas que se relacionan de manera directa con las características ecológicas y climáticas particulares del ambiente.

