











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La familia Buthidae es la más diversa entre los escorpiones en el mundo con 90 géneros actuales y 1,155 especies que representan 47.7% de las 2,423 descritas (Rein, 2019). El género Centruroides Marx, 1890, actualmente contiene 91 especies, 45 de las cuales se han descrito o registrado para México; sin embargo, en este conteo se considera a Centruroides elegans insularis como una especie (Ponce-Saavedra et al., 2016; Teruel y Myers, 2017), a pesar de que no se tiene ninguna propuesta ni evidencia formal al respecto. Por lo anterior, el número actual de especies del género en México debe ser de 44 (48.4%) y 2 con estatus subespecífico.
A partir del trabajo de Hoffmann (1932), el género Centruroides se divide en grupos de especies rayadas y no rayadas. Posteriormente, se han reconocido 4 grupos que se resumen en: a) grupo “gracilis”, b) grupo “bertholdi”, c) grupo “thorelli” y d) grupo de alacranes “rayados” (González-Santillán, 2001; Ponce-Saavedra y Francke, 2011a, b; Ponce-Saavedra y Moreno-Barajas, 2005). En este último, se han reconocido diferentes subgrupos con diferentes nombres (González-Santillán, 2001; Ponce-Saavedra y Moreno-Barajas, 2005; Santibáñez-López y Ponce-Saavedra, 2009) con base en las diferencias de coloración del carapacho, sin que a la fecha exista una propuesta que incluya a todas las especies conocidas en el país; por esta razón, en este trabajo se resume la información existente y se propone se incluya a los alacranes hasta ahora conocidos como subgrupos de rayados, en 3 grupos: “elegans”, “infamatus” y “nigrovariatus”, que se adicionan a los grupos “bertholdii”, “gracilis” y “thorelli”, generalmente aceptados (tabla 1).
Tabla 1 Especies del género Centruroides ubicadas en grupos. Modificado de Hoffmann (1932), González-Santillán (2001), Ponce-Saavedra y Moreno-Barajas (2005), Martín-Frías et al. (2007), Santibañez-López y Ponce-Saavedra (2009), Teruel et al. (2015).
| Grupo | Especie | Importancia para la salud pública * |
| “bertholdii” | C. bertholdii (Thorell, 1876) * | |
| Especies de tamaño variable, con 8 hileras o series de granulaciones en el borde interno de los dedos de la que la de los pedipalpos; coloración uniforme en carapacho y mesosoma, obscura, ocre o castaña. Los pedipalpos y patas pueden ser de coloración más clara | C. chiapanensis Hoffmann, 1932* | |
| C. edwardsii (Gervais, 1843) | ||
| C. flavopictus (Pocock, 1898) * | ||
| • C. flavopictus meridionalis Hoffmann, 1932 * | ||
| C. huichol Teruel, Ponce Saavedra y Quijano-Ravell, 2015 | X | |
| C. noxius Hoffman, 1932 * | X | |
| C. ochraceus (Pocock, 1898) * | ||
| C. tapachulaensis Hoffmann, 1932 | ||
| 8 especies y 1 subespecie | ||
| “elegans” | C. balsasensis Ponce-Saavedra y Francke, 2004 * | X |
| Especies de tamaño mediano (40 a 70 mm) con 7-8 hileras o series de granulaciones en el borde interno de los dedos de la quela de los pedipalpos; presentan 4 líneas longitudinales en el carapacho, siempre bien definidas en la parte anterior por debajo de los ojos laterales y 2 anteriores a los ojos medios. Posteriormente, las líneas medias pueden estar reducidas o poco evidentes. Las externas pueden variar en grosor a lo largo del carapacho e incluso estar reducidas. El mesosoma con 2 bandas de pigmento separadas por una banda mesal clara, estas se forman por manchas en los terguitos | C. bonito Quijano-Ravell, Treuel y Ponce-Saavedra, 2016 | X |
| C. chamela Ponce-Saavedra y Francke, 2011 * | X | |
| C. elegans (Thorell, 1876) * | X | |
| • C. elegans insularis Pocock, 1902 * | ||
| C. exilicauda (Wood, 1863) | ||
| C. hirsutipalpus Ponce-Saavedra y Francke, 2009 * | X | |
| C. limpidus (Karsch, 1879) * | X | |
| C. mascota Ponce-Saavedra y Francke, 2011* | X | |
| C. meisei Hoffmann, 1939 * | X | |
| C. pallidiceps Pocock, 1902 * | ||
| C. sculpturatus Ewing, 1928 | X | |
| C. tecomanus Hoffmann, 1932 * | X | |
| 12 especies y 1 subespecie | ||
| “gracilis” | C. caribbeanus Teruel y Myers, 2017 * | |
| Especies de tamaño grande (< 70 mm) con 9 series de granulaciones en el borde interno de los dedos de los pedipalpos; coloración uniforme castaña a negra con patas y que las variables, generalmente de tonalidad más clara. Los peines pueden ser blancos y contrastantes con el color general | C. chamulaensis Hoffmann, 1932 * | |
| C. fulvipes (Pocock, 1898) * | ||
| C. gracilis (Latreille, 1804) | ||
| C. nigrescens (Pocock, 1898) * | ||
| C. nigrimanus (Pocock, 1898) * | ||
| 6 especies | ||
| “infamatus”: | C. infamatus (C.L. Koch, 1844) * | |
| Especies de tamaño mediano (40-70 mm) con 7-8 hileras o series de granulaciones en el borde interno de los dedos de la quela de los pedipalpos; carapacho con pigentación difusa a intensa en la mayor parte, formando diferentes patrones que pueden ser característicos de las especies. El mesosoma con bandas de pigmento de grosor variable, formadas por manchas en los terguitos. El mesosoma con 2 bandas de pigmento separadas por una banda mesal clara que se forman por manchas en los terguitos | C. lauriadnae n. sp. * | |
| C. ornatus Pocock, 1902 * | ||
| C. ruana Quijano-Ravell y Ponce-Saavedra, 2016 | X | |
| C. suffusus (Pocock, 1902) * | X | |
| C. vittatus (Say, 1821) | X | |
| 6 especies | X | |
| X | ||
| “nigrovariatus” | C. baergi Hoffmann, 1932* | X |
| Especies de tamaño pequeño a mediano (30-60 mm) con el cuerpo y apéndices densamente manchados; el mesosoma con 2 bandas de pigmento obscuro que pueden ser poco conspicuas; quela del pedipalpo delgada y densamente granulosa, sobre todo en machos; metasoma más ancho de manera progresiva hacia la vesícula, claramente más ancho y alto que los últimos segmentos metasomales, más evidente en hembras; placa basal de peines de las hembras que puede tener un orificio central. Coloración variable. | C. franckei Santibáñez-López y Contreras-Félix, 2013* | |
| C. hoffmanni Armas, 1996* | ||
| C. nigrovariatus (Pocock, 1898)* | X | |
| C. orizaba Armas y Martin-Frías, 2003* | ||
| C. poncei Teruel, Kovařík, Baldazo-Monsivaiz y Hoferek, 2015* | ||
| C. rodolfoi Santibáñez-López y Contreras-Félix, 2013* | ||
| C. serrano Santibáñez-López y Ponce-Saavedra, 2009* | ||
| C. villegasi Baldazo-Monsivaiz, Ponce-Saavedra y Flores-Moreno, 2013* | ||
| 9 especies | ||
| “thorelli” | C. rileyi Sissom, 1995 * | |
| Especies de tamaño pequeño (> 40 mm) con pigmentación en todo el cuerpo en forma de manchas, sin llegar a constituir bandas en el mesosoma; el segmento metasomal V puede ser de color más obscuro y por lo menos las especies hasta ahora conocidas, con hábitos arborícolas. | C. schmidti Sissom, 1995 | |
| C. sissomi Armas, 1996 * | ||
| C. tuxtla Armas, 1999 * | ||
| 4 especies | ||
* Especies endémicas de México;
• estatus subespecífico.
Las especies del género Centruroides hasta ahora registradas en el norte de México son C. vittatus (Say) y C. sufussus (Pocock) que se ubicarían en el grupo “infamatus”; además de C. pallidiceps Pocock, C. sculpturatus Ewing y C. exilicauda (Wood) del grupo “elegans”.
En Sonora se tiene registro de 5 familias, 13 géneros y 30 especies de escorpiones, 3 de ellas con poblaciones con estatus subespecífico (tabla 2).
Tabla 2 Alacranes registrados para Sonora. Fuente: datos no publicados de O. F. Francke.
| Familia | Género | Especie | Subespecie |
| Buthidae | Centruroides Marx, 1890 | pallidiceps Pocock, 1902 | |
| sculpturatus Ewing, 1928 | |||
| Diplocentridae | Diplocentrus Peters, 1861 | gertschi Sissom et Walker, 1992 | |
| peloncillensis Francke, 1975 | |||
| spitzeri Stahnke, 1970 | |||
| williamsi Sissom et Wheeler, 1995 | |||
| Caraboctonidae | Hadrurus Thorell, 1876 | arizonensis | arizonensis Ewing, 1928 |
| pallidus Williams, 1970 | |||
| Superstitioniidae | Superstitionia Stahnke, 1940 | donensis Stahnke, 1970 | |
| Vaejovidae | Chihuahuanus González et Prendini 2013 | russelli (Williams, 1971) | |
| Gertschius Graham et Soleglad 2006 | agilis (Sissom et Stockwell, 1991) | ||
| crassicorpus Graham et Soleglad, 2007 | |||
| Kochius Soleglad et Fet, 2008 | sonorae (Williams, 1971) | ||
| Paravaejovis Williams, 1980 | confusus (Stahnke, 1940) | ||
| spinigerus (Wood, 1863) | |||
| Paruroctonus Werner, 1934 | baergi (Williams et Hadley, 1967) | ||
| borregoensis | borregoensis Williams, 1972 | ||
| stahnkei (Gertsch et Soleglad, 1966) | |||
| xanthus (Gertsch et Soleglad, 1966) | |||
| Stahnkeus Soleglad et Fet, 2006 | allredi (Sissom et Stockwell, 1991) | ||
| polisi (Sissom et Stockwell, 1991) | |||
| subtilimanus (Soleglad, 1972) | |||
| Serradigitus Stahnke, 1974 | hearnei (Williams, 1980) | ||
| yaqui Sissom et Stockwell, 1991 | |||
| Smeringurus Haradon, 1983 | aridus (Soleglad, 1972) | ||
| mesaensis (Stahnke, 1957) | |||
| Vaejovis C. L. Koch, 1836 | decipiens Hoffmann, 1931 | ||
| mauryi Capes, 2001 | |||
| pequeno Hendrixson, 2001 |
En la presente contribución se describe Centruroides lauriadnae sp. n., alacrán rayado que se incluye en el grupo “infamatus” por su patrón de coloración en el carapacho, incrementando a 31 el total de especies para el estado y a 45 las especies del género Centruroides en México.
Materiales y métodos
Para la nomenclatura y medidas se siguió a Stahnke (1970), excepto para tricobotrias (Vachon, 1974), carenas del metasoma (Francke, 1977), carenas de la mano (Acosta et al., 2008) y superficies del carapacho (Prendini et al., 2003).
Las observaciones se hicieron con un microscopio estereoscópico Stemi DV4. Las medidas fueron tomadas en milímetros (mm) con un micrómetro calibrado a 10X. Se tomaron imágenes digitales con luz visible con una cámara Cannon SX700 HS que posteriormente fueron editadas con Adobe Photoshop® CS5 para mejorar contraste y eliminar fondo. Para las ornamentaciones del carapacho se tomaron fotografías bajo luz negra cercana a UV y se editaron para obtener el mejor contraste y definición. Para estructuras específicas se utilizó una cámara MU100 con lente FMA050 de AmScope de 10 megapixeles de resolución, fotografías posteriormente acopladas con el software Combine ZP (Hadley, 2010) y editadas con Adobe Photoshop® CS5 para mejorar contraste y eliminar fondo. El mapa de distribución se generó utilizando el programa Maptool (http://www.seaturtle.org/maptool/). El material tipo será depositado en la Colección Nacional de Arácnidos (CNAN) del Instituto de Biología de la Universidad Nacional Autónoma de México y en la Colección Aracnológica de la Facultad de Biología de la Universidad Michoacana de San Nicolás de Hidalgo (CAFBUM).
Descripción
Familia Buthidae C.L. Koch, 1837
Género Centruroides Marx, 1890
Centruroides lauriadnae n. sp.
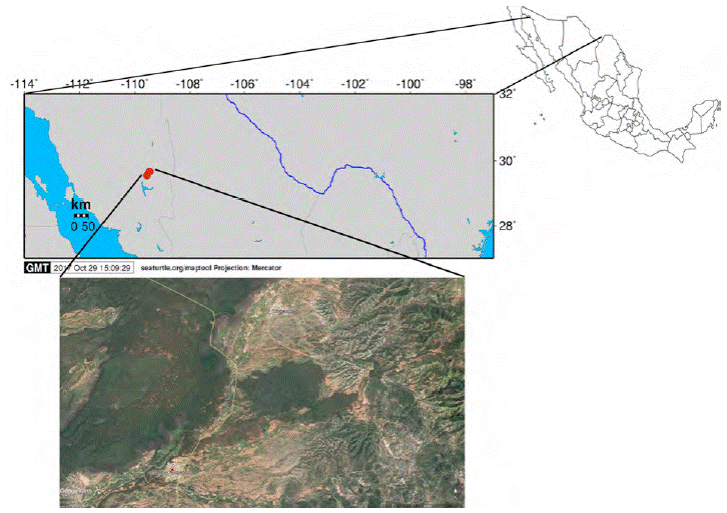
Figura 1. Ubicación de las localidades en que se colectaron los ejemplares tipo de Centruroides lauriadnae sp. n.

Figura 2. Habitus de Centruroides lauriadnae sp. n. Macho: a) vista dorsal, b) ventral, hembra: c) vista dorsal, d) ventral.

Figura 3. Centruroides lauriadnae sp. n. Carapacho, mesosoma y pedipalpo; vista dorsal. Macho holotipo: a, c, e; hembra paratipo: b, d, f.

Figura 4. Centruroides lauriadnae sp. n. Macho holotipo: a) pedipalpo en vista ventral, b) mano en vista dorsal, c) terguito VII, d) esternitos VI y VII, e) vista ventrolateral de la pata IV; f-k, metasoma: f), segmento I en vista dorsal, g) segmento II en vista dorsolateral, h) segmento III en vista dorsolateral, i) segmento IV en vista lateral, j) segmento V y telson en vista lateral, k) vesícula y aguijón en vista lateral.
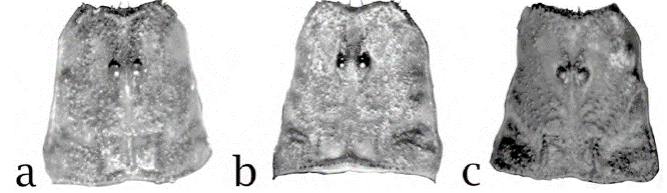
Figura 5. Carapacho bajo luz UV, a) macho holotipo de Centruroides lauriadnae sp. n., b) macho de C. sculpturatus, c) macho de C. pallidiceps.

Figura 6. a) Esternón, opérculo genital y estructuras pectinales del macho holotipo de Centruroides lauriadnae sp. n.; b) acercamiento del opérculo genital y placa basal de los peines. p) parche blanquecino en esternito III.

Figura 7. Esternón, opérculo genital y estructuras pectinales de la hembra paratipo de Centruroides lauriadnae sp. n.

Figura 8. Habitus de Centruroides sculpturatus en vista dorsal y ventral: a) y b) hembra; c) y d) macho. Fotografías de T. R. Van Devender en abril 14 de 2016 en sierra Huérfana, Sonora.

Figura 9. Habitus de Centruroides pallidiceps en vistas dorsal y ventral: a) y b) hembra; c) y d) macho.
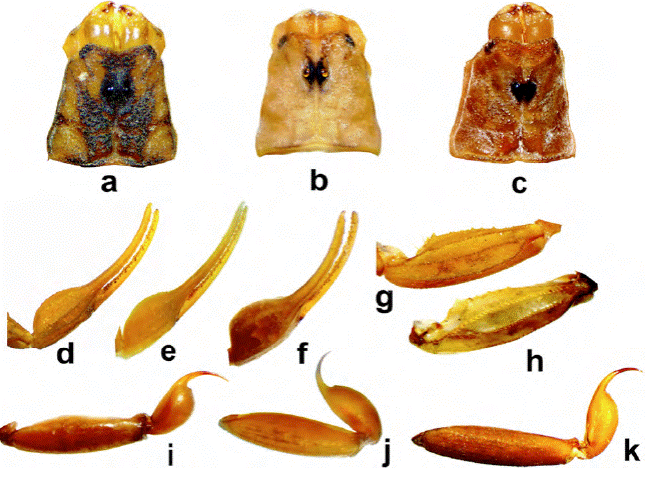
Figura 10. Comparación de estructuras entre machos de diferentes especies; carapacho: a) C. lauriadnae sp. n.; b) C. sculpturatus; c) C. pallidiceps. Vista dorsal de la mano: d) C. lauriadnae sp. n.; e) C. sculpturatus; f) C. pallidiceps; vista dorsal de la patela pedipalpal: g) C. lauriadnae sp.n.; h) C. pallidiceps; vista lateral del segmento metasomal V y vesícula: i) C. pallidiceps; j) C. sculpturatus; k) C. lauriadnae sp. n.
Tabla 3 Medidas de los ejemplares tipo de Centruroides lauriadnae sp. n. Las medidas están expresadas en mm. CDp = Conteo de dientes pectinales; Ltot = longitud total; LCtx = longitud del carapacho; AantCtx = ancho anterior del carapacho; ApostCtx = ancho posterior del carapacho; Lmeso = longitud del mesosoma; LSMI = longitud del segmento metasomal I; LSMII = longitud del segmento metasomal II; LSMIII = longitud del segmento metasomal III; LSMIV = longitud del segmento metasomal IV; LSMV = longitud del segmento metasomal V; Lves = longitud de la vesícula; Lfem = longitud del fémur del pedipalpo; Lpat = longitud de la patela del pedipalpo; Lman = longitud de la mano del pedipalpo; Ldf = longitud del dedo fijo de la quela del pedipalpo; Ldm = longitud del dedo móvil de la quela del pedipalpo; LSMesVII = longitud del segmento mesosomal VII; LBpei = longitud de la base de los peines; ABpei = ancho de la base de los peines; Aves = ancho de la vesícula; Hves = alto de la vesícula.
| Ejemplar | CDp | Ltot | LCtx | Aant Ctx | Apost Ctx | Lmeso | LSMI | LSMII | LSMIII | LSMIV | LSMV |
| Macho holotipo | 24-24 | 42.24 | 4.42 | 1.78 | 4.45 | 11.38 | 3.96 | 4.98 | 5.28 | 5.61 | 6.60 |
| Macho paratipo 1 | 24-25 | 45.14 | 4.83 | 2.08 | 4.98 | 12.55 | 4.29 | 5.08 | 5.56 | 5.94 | 6.88 |
| Macho paratipo 2 | 23-xx | 36.75 | 4.01 | 1.57 | 4.11 | 10.39 | 3.56 | 4.01 | 4.42 | 4.67 | 5.69 |
| Macho paratipo 3 | 24-23 | 40.54 | 4.57 | 1.93 | 4.42 | 10.74 | 3.96 | 4.65 | 4.98 | 5.41 | 6.22 |
| Macho paratipo 4 | 24-24 | 40.16 | 4.50 | 1.83 | 4.42 | 11.86 | 3.68 | 4.45 | 4.75 | 4.75 | 6.17 |
| Macho paratipo 5 | 25-24 | 40.03 | 4.62 | 2.13 | 4.32 | 10.29 | 4.14 | 4.67 | 4.67 | 5.13 | 6.50 |
| Macho paratipo 6 | 24-24 | 39.88 | 4.32 | 1.98 | 4.27 | 10.03 | 4.14 | 4.50 | 4.98 | 5.38 | 6.53 |
| Macho paratipo 7 | 25-25 | 39.52 | 4.09 | 1.96 | 4.09 | 10.49 | 3.63 | 4.75 | 4.85 | 5.44 | 6.27 |
| Macho paratipo 8 | 25-25 | 35.79 | 4.11 | 1.73 | 3.71 | 10.57 | 3.28 | 3.89 | 4.24 | 4.39 | 5.31 |
| Moda= 24 | |||||||||||
| n=17 | 5.59 | ||||||||||
| Hembra paratipo 1 | 22-21 | 41.15 | 4.90 | 2.67 | 5.23 | 12.65 | 5.23 | 4.06 | 4.22 | 4.50 | 5.99 |
| Hembra paratipo 2 | 21-23 | 41.48 | 5.33 | 2.59 | 5.59 | 12.60 | 3.71 | 4.27 | 4.60 | 4.98 | 5.44 |
| Hembra paratipo 3 | 24-24 | 37.64 | 4.98 | 2.18 | 5.00 | 11.61 | 3.20 | 3.84 | 4.17 | 4.42 | 5.66 |
| Hembra paratipo 4 | 22-21 | 39.60 | 5.08 | 2.24 | 5.38 | 12.42 | 3.38 | 4.17 | 4.45 | 4.45 | 5.21 |
| Hembra paratipo 5 | 22-22 | 36.73 | 4.70 | 1.98 | 5.03 | 11.18 | 3.25 | 3.96 | 4.06 | 4.37 | 5.84 |
| Hembra paratipo 6 | 23-22 | 43.46 | 5.61 | 2.34 | 5.84 | 14.66 | 3.76 | 4.39 | 4.57 | 4.62 | 5.74 |
| Hembra paratipo 7 | 22-22 | 39.24 | 5.18 | 1.85 | 5.79 | 11.89 | 3.25 | 3.94 | 4.45 | 4.80 | 5.41 |
| Hembra paratipo 8 | 23-xx | 37.26 | 4.98 | 2.24 | 4.83 | 11.28 | 3.18 | 3.91 | 4.11 | 4.39 | 5.44 |
| Hembra paratipo 9 | 22-21 | 40.18 | 4.85 | 2.21 | 4.65 | 13.94 | 3.43 | 4.04 | 4.24 | 4.24 | ¿???? |
| Moda= 22 | |||||||||||
| n=17 | |||||||||||
| Ejemplar | Lves | Aves | Hves | Lfem | Lpat | Lman | Ldf | Ldm | LSMesVII | LBpei | ABpei |
| Macho holotipo | 2.49 | 1.78 | 1.47 | 4.19 | 5.13 | 3.20 | 4.57 | 5.13 | 3.20 | 0.58 | 0.94 |
| Macho paratipo 1 | 3.10 | 1.52 | 1.6 | 4.57 | 5.33 | 2.69 | 5.00 | 5.59 | 3.68 | 0.53 | 1.02 |
| Macho paratipo 2 | 2.79 | 1.40 | 1.35 | 3.76 | 4.55 | 2.34 | 4.27 | 4.65 | 3.00 | 0.53 | 1.02 |
| Macho paratipo 3 | 3.00 | 1.47 | 1.37 | 4.19 | 5.11 | 3.07 | 4.98 | 5.05 | 3.48 | 0.56 | 1.07 |
| Macho paratipo 4 | 2.57 | 1.42 | 1.40 | 4.42 | 4.83 | 2.82 | 4.83 | 5.08 | 3.23 | 0.48 | 0.99 |
| Macho paratipo 5 | 2.62 | 1.45 | 1.49 | 4.29 | 4.98 | 2.97 | 4.93 | 5.46 | 3.15 | 0.53 | 1.09 |
| Macho paratipo 6 | 3.00 | 1.50 | 1.52 | 4.17 | 4.90 | 3.12 | 4.67 | 5.38 | 3.12 | 0.61 | 1.12 |
| Macho paratipo 7 | 2.74 | 1.40 | 1.42 | 4.01 | 4.42 | 3.18 | 4.57 | 5.33 | 3.02 | 0.48 | 0.79 |
| Macho paratipo 8 | 2.44 | 1.35 | 1.35 | 3.73 | 4.19 | 2.74 | 4.01 | 4.62 | 3.05 | 0.51 | 0.89 |
| Hembra paratipo 1 | 4.88 | 1.63 | 1.68 | 4.78 | 5.38 | 2.82 | 5.28 | 5.89 | 3.28 | 0.71 | 1.04 |
| Hembra paratipo 2 | 2.97 | 1.68 | 1.80 | 4.90 | 5.61 | 3.07 | 6.30 | 6.38 | 3.71 | 0.86 | 1.09 |
| Hembra paratipo 3 | 2.18 | 1.55 | 1.73 | 4.14 | 4.93 | 2.92 | 4.93 | 5.54 | 3.15 | 0.81 | 1.07 |
| Hembra paratipo 4 | 2.57 | 1.57 | 1.67 | 4.57 | 5.28 | 2.62 | 5.13 | 5.79 | 3.43 | 0.94 | 1.09 |
| Hembra paratipo 5 | 2.31 | 1.55 | 1.55 | 3.91 | 4.83 | 2.72 | 4.88 | 5.38 | 3.00 | 0.91 | 1.04 |
| Hembra paratipo 6 | 2.67 | 1.73 | 1.78 | 4.98 | 5.69 | 3.18 | 5.82 | 15.75 | 3.35 | 0.84 | 1.30 |
| Hembra paratipo 7 | 2.59 | 1.70 | 1.68 | 4.37 | 5.44 | 3.25 | 5.84 | 6.17 | 4.17 | 0.91 | 1.12 |
| Hembra paratipo 8 | 2.54 | 1.50 | 1.65 | 4.37 | 4.95 | 2.64 | 5.31 | 5.66 | 3.07 | 0.74 | 1.02 |
| Hembra paratipo 9 | 2.36 | 1.52 | 1.55 | 4.01 | 2.29 | 3.05 | 5.26 | 5.61 | 3.66 | 0.79 | 1.09 |
Diagnosis. La siguiente combinación de caracteres es diagnóstica para Centruroides lauriadnae sp. n. Longitud total de los adultos entre 35 y 46 mm. Coloración típica de un alacrán “rayado”, con el carapacho fuertemente pigmentado y el patrón del subgrupo “infamatus”: dorsalmente ocre y ventralmente amarillo muy claro, con metasoma y patas amarillo más claro. Pedipalpos con coloración ligeramente más clara y débilmente infuscados. Superficies intercarenales del carapacho con granulación gruesa e irregular; en terguitos mesosomales con granulación más fina y en metasoma más dispersa y fina, pero presente en todos los segmentos. Conteo de dientes pectinales de 23-25 (moda = 24, n = 17) en los machos y un poco más bajo (21-24, moda = 22, n = 17) en hembras. Carenas metasomales moderadas a bien desarrolladas y granulares, excepto en el segmento V del macho, débiles basalmente y muy débiles en el resto del segmento. Segmento metasomal V de 1.3 a 1.5 veces más largo que el carapacho y la longitud del metasoma es menor de 1.6 veces la longitud del resto del cuerpo. La vesícula es alargada y con el tubérculo subaculear cónico y ubicado cerca de la base del aguijón, a diferencia de C. sculpturatus y C. pallidiceps, en los que dicho tubérculo se encuentra más alejado de la base del aguijón. Entre los alacranes que habitan en el norte de México, C. lauriadnae sp. n. es similar a C. suffusus en coloración y conteo de dientes pectinales con 23-25 (moda = 24) en machos por 22-25 (moda = 23) en C. suffusus y 21-24 (moda = 22) en hembras por 21-23 (moda 23) en C. suffusus; sin embargo, la especie nueva es de menor tamaño, ya que sus ejemplares más grandes apenas se acercan a los 50 mm, mientras que los de C. suffusus, en general, son mayores a 55 mm; además, el metasoma de los machos de C. suffusus es 1.8 veces más largo que el resto del cuerpo, mientras que esta relación es menor a 1.6 veces en C. lauriadnae sp. n. El tubérculo subaculear es cónico y su ubicación cercana a la base del aguijón es distinto al de C. suffusus, que es poco desarrollado y se ubica ligeramente más alejado de la base del aguijón. Las otras 2 especies son menos similares en apariencia general, pero geográficamente cercanas: C. sculpturatus, registrada solo en el estado de Sonora y C. pallidiceps, con registros en Sonora y Sinaloa (Ponce-Saavedra et al., 2016), tienen coloración menos intensa en el carapacho e incluso cuando adultos, pueden ser completamente inmaculados o el pigmento se reduce a 2 manchas en la región posterior. Centruroides sculpturatus tiene dedos relativamente más largos con respecto a la mano de la nueva especie (proporción longitud dedo fijo/longitud de la mano > de 2) y un conteo de dientes pectinales claramente más alto en machos de C. sculpturatus (24-28). La mano más globosa, con carenas muy débiles y la carena media de la patela del pedipalpo curvada en C. pallidiceps, en contraste con la casi recta, completa y bien desarrollada carena media de la nueva especie. Los valores más bajos en longitud del metasoma de los machos con respecto a la longitud del carapacho (1.2-1.4), pueden ser un buen complemento para definir la nueva especie.
Descripción del macho holotipo (figs. 2a, b; 3a, c, e; 4a-k; 5a; 6a, b; 10a, d, k; medidas en la tabla 3).
Coloración. Típica de un alacrán “rayado”, dorsalmente de color ocre con metasoma y patas de color más claro, ventralmente de color amarillo muy claro (figs. 2a, b). Carapacho (fig. 3a) con el borde anterior entre las hileras de ojos laterales pigmentado de color negro, angostándose hacia los bordes de las crestas superciliares, dejando el resto del carapacho claro, con manchitas ligeramente difusas que llegan hasta el borde del surco ocular, dejando entonces 2 áreas claras delimitadas, una en la zona de los ojos laterales y la otra por debajo de ellos, separadas por una línea oblicua de gránulos pigmentados que llegan a la altura del margen anterior de los ojos medios. El surco ocular medio es muy obscuro, al igual que los surcos oculares laterales, el pigmento obscuro se extiende hasta la parte anterior del surco medio posterior. A la altura de los ojos medios inicia una mancha obscura que se ensancha oblicuamente hasta el surco medio posterior. A los lados del surco medio anterior se forman 2 áreas arriñonadas sin pigmento y 2 más entre la línea de gránulos obscuros que salen de la parte posterior de los ojos laterales y se extiende de la quilla ocular lateral hacia el borde. El surco central transverso está limitado por gránulos grandes y negros unidos en el surco posterior por el área pigmentada del mismo, dejando solo 2 pequeñas áreas con menos pigmento a cada lado del surco. La mitad distal del surco medio posterior es claro y termina ensanchándose en el borde posterior del carapacho en un amplio surco marginal posterior dando la apariencia de una T invertida. Las áreas laterales son casi completamente claras, solo se aprecia pigmento difuso en los bordes, el cual se une con unos pocos gránulos obscuros a las manchas medias y posteriores que se encuentran a los lados del surco medio. El borde posterior termina con una línea negra del mismo ancho que tendrán las bandas mesosomales formadas por las manchas en los terguitos. Los quelíceros (fig. 3a) presentan pigmentación reticulada en toda su base y son claros en el resto de su superficie, excepto por el color pardo obscuro de la punta de los dientes. Dorsalmente, el trocánter, el fémur, la patela y las manos de los pedipalpos presentan manchas obscuras, dispersas y difusas, sin llegar a tener el tono obscuro de las manchas del carapacho o los terguitos (fig. 3e). El dorso de la mano es claramente manchado, no así los dedos, completamente claros, excepto en la base del dedo fijo hacia la zona de la articulación del dedo móvil (fig. 4b). En su superficie ventral, los pedipalpos están completamente inmaculados incluyendo mano y dedos (fig. 4a). Mesosoma (fig. 3b) con terguitos de coloración similar, con el preterguito en su borde posterior intensamente pigmentado de negro y conectado por medio de pigmentación difusa y reticulada con una línea transversa negra en el borde posterior del posterguito. Terguitos I y II con la línea pigmentada del borde posterior del posterguito engrosándose hacia la parte media. En los terguitos II al VI, la mancha posterior se ensancha hacia la parte media del posterguito formando una figura trapezoidal con un espacio claro y una pigmentación reticulada que se une a la línea gruesa del preterguito. En el terguito VII (fig. 4c), este patrón se pierde y se observa pigmentación ligeramente más clara que en los terguitos anteriores. En general, no se aprecia ningún patrón de color en esternitos, excepto en el VII (fig. 4d) que tiene ligera pigmentación en las carenas submedias y laterales. Las patas (fig. 4e), ventralmente de coloración muy clara e inmaculadas distalmente, se obscurecen hacia el fémur, dorsal y dorsolateralmente jaspeadas, con pigmento muy aparente en la cara dorsolateral del trocánter, fémur, patela, tibia y basitarso en todas las patas, excepto en pata I, la cual tiene pigmentación más tenue. Metasoma dorsalmente inmaculado en todos sus segmentos, pero las carenas dorsolaterales, desde el segmento I hasta el III, están pigmentadas, siendo más fuertes en el segmento I (fig. 4f-i) y se desvanecen hacia el IV. Las áreas dorsolaterales y laterales están difusamente pigmentadas dando apariencia de estar manchadas en todos los segmentos. Segmento metasomal V (fig. 4j) más obscuro que los anteriores y que la vesícula, la cual es de tonalidad más clara (fig. 4k), muy ligeramente manchada sobre el lado ventral cerca del tubérculo subaculear y la porción distal del aguijón es obscura. Ventralmente, los segmentos I-IV pigmentados en la región media entre las carenas submedianas ventrales formando una banda continua hasta el segmento IV, ya que el V es de coloración uniforme (fig. 4f-i). Las carenas ventrolaterales también tienen ligera pigmentación en su porción interna dando la apariencia, en los segmentos I-III, de que el animal tuviera 2 líneas muy delgadas flanqueando una de posición medial, claramente más gruesa y pigmentada.
Carapacho. Ojos redondos, ampliamente separados (0.30 mm); borde anterior del carapacho recurvado, ligeramente crenulado, con gránulos muy fuertes sobre el surco medio anterior que llegan a las crestas superciliares, las cuales son gruesas, granulares y se prolongan hacia el borde anterior con gránulos grandes e irregulares (fig. 3a). Hay 3 ojos laterales. Los bordes laterales del carapacho en la parte más anterior claramente crenulados tornándose granulares hasta que en la parte posterior es claramente granuloso, formado por gránulos grandes separados por gránulos pequeños; en la zona del surco medio posterior se forma un grupo de gránulos irregulares inmersos en una granulación muy fina que se extiende anteriormente en el área del surco medio posterior, el cual también es finamente granulado. Las superficies intercarenales y áreas sin pigmento, se aprecian casi lisas o muy finamente granulosas, sobre todo en la parte media y anterior del carapacho. Las carenas posteriores laterales son muy fuertes, formadas por unos cuantos gránulos grandes (fig. 5a).
Pedipalpos. Trocánter con 4-5 setas basales y 2 distales, no se aprecian microsetas. El fémur, con la superficie dorsal cubierta por gránulos finos y dispersos, cara externa con la carena insinuada y superficie irregular sin llegar a ser granulosa. Patela (fig. 3e) con la carena media muy bien desarrollada, las dorsales externas crenuladas y ventroexternas ligeramente aserradas, ambas bien desarrolladas; las carenas internas con 5 gránulos grandes, la dorsointerna con una seta muy grande y un diente basal muy grande, que distalmente termina en pequeños dientecillos agudos; superficies intercarenales granulosas (fig. 3e). Mano (fig. 4b) con todas las carenas bien desarrolladas y granulares, digital completa, la dorsal secundaria también, pero es más débil hacia la base de la mano, la externa secundaria muy bien desarrollada y ligeramente crenulada a granular, la dorsal marginal formada por dientecillos separados, distribuidos uniformemente, los cuales se van haciendo más pequeños hacia la base del dedo fijo, la ventroexterna y la media interna con menor desarrollo. Dedos hirsutos, con las puntas densamente pilosas, con 8 hileras de dientecillos, la basal formada por 2 subhileras, una proximal identificable por 6 gránulos accesorios internos y 6 externos, y una distal identificable por 4 gránulos externos e internos de diferentes tamaños, la hilera terminal formada por 3 dientecillos. Ventralmente, los pedipalpos presentan una superficie finamente granulosa (fig. 4a). Coxas de pedipalpos, patas I-II y laminillas maxilares lisas al igual que todos los esternitos. Coxas de pedipalpos con una seta grande, coxas de patas I-II con 2 setas cerca del borde anterior y 2 hacia el borde posterior localizadas cerca de la articulación con el trocánter, coxas de patas III con 4 setas, coxas de patas IV con solo 1 par de setas al nivel del borde anterior de las placas genitales.
Patas. Las I-II sin características distintas al resto de las especies del género; III-IV con espuelas tarsales asimétricas, pero bien desarrolladas (fig. 4e), en IV hay una tercera espuela dorsointerna débilmente desarrollada.
Mesosoma. Terguitos con los preterguitos angostos. Preterguitos I-V (fig. 3c) con el borde anterior con granulación densa y muy fina, pero en todos los casos apreciable, el borde posterior con gránulos mucho más gruesos que en el anterior y ampliamente separados con un arreglo parecido al borde posterior del carapacho de 2 gránulos grandes separados por uno pequeño, excepto en el área sin pigmento que corresponde a la zona de la carena media. Las manchas posteriores del posterguito, todas con gránulos dispersos grandes y pequeños, aparentemente sin ningún arreglo, pero delimitando la mancha obscura (todos los gránulos se localizan sobre la zona con pigmentación obscura). Las áreas laterales a la carena media van incrementando su granulación a partir del terguito IV, siendo en V y VI muy evidente y también la carena media es más fuerte, condición que se pierde en el terguito VII, en donde solo se mantiene una fina granulación y la quilla no se aprecia bien desarrollada, aunque las carenas medias laterales son muy fuertes y completas; los espacios intercarenales son claramente granulares. Esta condición granulosa también se aprecia en las áreas sin pigmento en las zonas laterales de los terguitos. Placa basal de los peines rectangular, con una muesca muy marcada, borde anterior cóncavo y el posterior recto, conteo de dientes pectinales 24-24; placas genitales subtriangulares, con 3 setas en el borde posterior; esternón profundo, con 4 setas, 2 anteriores y 2 mediobasales (fig. 6). Esternito III con 1 seta anterior seguida por 5 pares de setas arregladas formando un triángulo, con un parche blanquecino en el borde medio posterior (fig. 6p); esternito IV con 6 setas, 2 medias, 2 medias laterales y 2 laterales posteriores, mismo número de setas en V y VI solo que más alineadas, de tal manera que en VI ya es muy corta la distancia entre pares de setas; VII con 2 setas medias y 2 medias laterales; las carenas medianas y laterales fuertes y crenuladas (fig. 4d).
Metasoma. Segmento I con las carenas dorsolaterales claramente aserradas, con dientes triangulares muy bien desarrollados al igual que las carenas laterales supramedianas y lateroventrales, la base de la carena inicia con 3 dientes romos grandes, carenas ventrales submedianas granulares en I, formadas por gránulos grandes y regulares, ocupando toda la longitud del segmento; segmento II con carenas menos fuertes, con sólo 2 dientes al inicio de las dorsolaterales las cuales son claramente aserradas, con dentículos más finos pero también completas, el resto similar al segmento I; segmento III con las carenas dorsolaterales fuertemente crenuladas en las que solamente se aprecian dentículos triangulares en la parte distal cerca de la articulación, las laterales supramedianas muy reducidas, moderadamente crenuladas; segmento IV con carenas dorsolaterales crenuladas, completas y con 2 dentículos triangulares distales, laterales supramedianas ligeramente granuladas, en su mayoría lisas, con gránulos separados e irregularmente distribuidos sobre la carena, ventrolaterales moderadamente crenuladas, bien desarrolladas y completas, ventrales submedianas ligeramente aserradas, más débiles que en el segmento anterior; segmento V más largo que el IV, de forma redondeada, rebordeado dorsalmente en la base y ligeramente acanalado en toda su longitud, se aprecia la quilla dorsolateral vestigial en la mayor parte del segmento y aparente en el tercio posterior del mismo, quilla media ventral distinta, muy débilmente aserrada pero completa (fig.4j). Los espacios intercarenales lisos a muy finamente granulosos. En el segmento I hay un patrón de 6 setas en el lado externo de las carenas ventrales submedianas, 2 en posición anterior, 2 en posición media posterior y otras 2 en el borde posterior del segmento al final de la banda de pigmento en medio de las carenas ventrales submedianas, en las ventrolaterales también externas a las carenas hay un par de setas en posición anterior y otro par en el cuarto posterior del segmento. Este patrón se mantiene en II, III y IV. En V también hay 4 setas, pero mucho más débiles que en los anteriores.
Telson. Vesícula rebordeada dorsalmente, acanalada, con una carena media ventral muy fuerte que llega hasta la base del aguijón, tubérculo subaculear cónico muy débil acompañado por al menos 8 macrosetas ventrales, 2 de ellas cerca del tubérculo además de varias microsetas dispersas en toda la superficie (fig. 4k). Ventral y ventrolateralmente con 2 setas en la base, 2 a un lado de las quillas que confluyen en la carena media formando una depresión antes del aguijón totalizando 8 setas grandes, incluyendo el par de setas que se encuentran por debajo del tubérculo subaculear.
Dimorfismo sexual. Las hembras presentan las diferencias típicas del género al ser más robustas y con los segmentos metasomales más cortos y gruesos. Adicionalmente se puede apreciar el carapacho de la hembra paratipo 1 ligeramente más moreno, con los espacios intercarenales en la mitad anterior un poco más obscuros y granulosos, con gránulos grandes irregulares y dispersos (fig. 3b); pedipalpos ligeramente más jaspeados (fig. 3f), con el fémur, patela y manos con todas las carenas más fuertes y granulares, con los espacios entre carenas finamente granulosos y con gránulos dispersos, grandes e irregulares; la placa basal de los peines es más cuadrada (proporción L/A entre 0.68 y 0.88; media = 0.76 ± 0.08); n = 9), con el borde anterior ligeramente cóncavo y el posterior claramente redondeado (fig. 7); mientras que en los machos son rectangulares y claramente más anchos que largos (proporción L/A entre 0.48 y 0.62; media = 0.54 ± 0.05 n = 9). El conteo de dientes pectíneos es más bajo (21-24, moda = 22, n = 17) por 23-25 (moda = 24, n = 17) de los machos. El terguito VII tiene carenas medianas dorsales menos desarrolladas y son muy débiles en las partes anterior y posterior, aparentando estar incompletas (figs. 2c, 3d). Esternito VII también con carenas medias ligeramente más débiles (fig. 2d). Los segmentos metasomales dorsalmente son profundamente cóncavos, fuertemente acanalados y con todas las carenas más fuertes que en los machos (fig. 2c); el segmento metasomal V es de apariencia recta, más largo que el IV, con carenas distintas y granulosas en la mitad basal y más débiles distalmente, aunque la superficie se mantiene finamente granulosa, dorsalmente con un surco profundo en toda su longitud, lo que le da una apariencia lobulada. La vesícula es ligeramente más redondeada (proporción L/A = 1.73 ± 0.48) y más alta (1.68 ± 0.09) que la de los machos en los que la proporción L/A (1.87 ± 0.20) es mayor y la altura menor (1.44 ± 0.09) (figs. 2c, d; tabla 3).
Variación. Carapacho con patrón de color bastante constante en todos los ejemplares machos (n = 10) y hembras (n = 9), variando el color de fondo de amarillo a ocre tenuemente rojizo. El tamaño de hembras y machos es similar y varía de 35 a 46 mm de longitud.
Resumen taxonómico
Material tipo. México: Sonora: Tepache, municipio de Tepache (fig. 1): holotipo ♂ (CNAN-T01260), localidad, 29°32.19’ N, 109°33.0’ O, 606 m snm, 6/Octubre/2012, Colector T. Van Devender. Paratipos: 7 ♂, 7 ♀ (CNAN-T01261), 2 ♂♂, 2 ♀♀ (CAFBUM -S0100) (tabla 4).
Tabla 4 Datos de los ejemplares tipo de Centruroides lauriadnae sp. n. depositados en Colección Nacional de Arácnidos (CNAN) del Instituto de Biología, Universidad Autónoma de México y en la Colección Aracnológica de la Facultad de Biología de la Universidad Michoacana de San Nicolás de Hidalgo (CAFBUM). Todos los ejemplares fueron colectados por T. R. Van Devender.
| Ejemplar | Datos de localidad | Coordenadas geográficas |
| ♂ Holotipo | Tepache, municipio de Tepache, Sonora. 270.3. En lava. 6/octubre/2012 | 29°32’11.1” N, 109°32’59.59” O |
| ♂Paratipo 1 | Divisadero, municipio de Tepache, Sonora. 317.1. En lava. 1/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♂Paratipo 2 | Tepache, municipio de Tepache, Sonora. 270.3. En lava. 7/octubre/2012 | 29°32’11.10” N 109°32’59.59” O |
| ♂Paratipo 3 | Tepache, municipio de Tepache, Sonora. 270.3. En lava. 6/octubre/2012 | 29°32’11.10” N, 109°32’59.59” O |
| ♂Paratipo 4 | Tepache, municipio de Tepache, Sonora. 270.3. En lava. 6/octubre/2012 | 29°32’11.10” N, 109°32’59.59” O |
| ♂Paratipo 5 | Tepache, municipio de Tepache, Sonora. 270.3. En lava. 6/octubre/2012 | 29°32’11.10” N, 109°32’59.59” O |
| ♂Paratipo 6 | Divisadero, municipio de Tepache, Sonora. 317.1. En lava. 7/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♂Paratipo 7 | Divisadero, municipio de Tepache, Sonora. 317.1. En lava. 7/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♂Paratipo 8 * | 337.5. En lava. 7/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♂Paratipo 9 * | Divisadero, municipio de Tepache, Sonora. 317.1. En lava. 7/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♀ Paratipo 1 | Tepache, municipio de Tepache, Sonora. 270.3. En lava. 6/octubre/2012 | 29°32’11.10” N, 109°32’59.59” O |
| ♀ Paratipo 2 | Divisadero, municipio de Tepache, Sonora. 317.1. En lava. 1/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♀ Paratipo 3 | Divisadero, municipio de Tepache, Sonora. 316.7. En lava. 7/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♀ Paratipo 4 | Divisadero, municipio de Tepache, Sonora. 316.7. En lava. 7/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♀ Paratipo 5 | Divisadero, municipio de Tepache, Sonora. 316.7. En lava. 7/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♀ Paratipo 6 | Divisadero, municipio de Tepache, Sonora. 317.1. En lava. 1/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♀ Paratipo 7 | Divisadero, municipio de Tepache, Sonora. 317.1. En lava. 1/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♀ Paratipo 8 * | 337.5. En lava. 7/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
| ♀ Paratipo 9 * | Divisadero, municipio de Tepache, Sonora. 317.1. En lava. 1/octubre/2012 | 29°36’43.14” N, 109°27’58.33” O |
* Paratipos depositados en CAFBUM
Etimología. El epíteto específico es un nombre en genitivo formado a partir de los nombres Laura y Ariadna, hijas del primer autor, a quienes está dedicada esta especie.
Aspectos ecológicos. Esta especie fue colectada en un hábitat con lava en un pequeño valle de la sierra Madre Occidental, al oeste de Sonora, a una altitud de ± 600 m snm con muy escasa vegetación de tipo matorral subtropical y un clima seco cálido, con una temperatura media máxima mensual de 31.2 °C y una temperatura media mínima mensual de 13.7 °C. La temperatura media anual es de 22.4 °C; la época de lluvias se presenta en verano en los meses de julio y agosto con una precipitación media anual de 520.8 mm (Servicio Meteorológico Nacional, 2016). El hábitat y el habitus son consistentes con el ecomorfotipo lapidícola (Prendini, 2001).
Distribución. La especie se conoce solo de la localidad tipo (fig. 1).
Comentarios taxonómicos
Se hizo una comparación con las descripciones y los ejemplares de las 2 especies del género hasta ahora registradas para Sonora, C. sculpturatus Ewing y C. pallidiceps Pocock; además de C. suffusus, reportada en Durango y Sinaloa.
La especie de alacrán más común en Sonora y cuya distribución en México, a la fecha, se restringe a localidades de esa entidad es C. sculpturatus (fig. 8), especie de tamaño mediano entre 48 a 55 mm de longitud total (Stahnke, 1971). Es un alacrán de importancia médica y su distribución alcanza el estado de Arizona en el sur de los EUA.
Centruroides lauriadnae sp. n. es un alacrán de tamaño mediano (35 a 46 mm) y apariencia general distinta, pudiendo resaltarse las siguientes diferencias: la coloración del carapacho es del tipo “infamatus” (figs. 3a, 10a), más notoria en el carapacho de las hembras (fig. 3b); mientras que en C. sculpturatus, la mayor parte del carapacho es clara y solo se observa pigmento obscuro en la parte media anterior y sobre los ojos laterales, así como unos cuantos gránulos ubicados en línea oblicua por debajo del ojo lateral más externo, los ojos medios y 2 líneas que ocupan la mitad posterior del carapacho llegando hasta el borde (fig. 8a, c); en ejemplares conservados en alcohol, puede observarse la pigmentación castaña en la parte anterior y al menos sobre el borde posterior y gránulos adyacentes (fig. 10b). Los terguitos de C. lauriadnae sp. n. tienen las manchas de pigmento claramente más anchas y obscuras, dejando la banda mesal, sin pigmento, mucho más delgada, quillas gruesas muy notorias y con granos grandes e irregulares (fig. 3c, d). En C. sculpturatus, las bandas de pigmento lucen más delgadas y por ende la banda mesal es más ancha (fig. 8a, c); las quillas son débiles a moderadas y la granulación es mucho más fina. En machos, la mano del pedipalpo de C. lauriadnae sp. n. es un poco más globosa y con los dedos proporcionalmente menos largos que la mano con respecto a la proporción en C. sculpturatus, cuyos dedos son relativamente más largos con respecto a la mano (proporción longitud dedo móvil/ longitud de la mano > 2), la cual es ligeramente más esbelta (fig. 10d, e). La parte anterior del carapacho de la especie nueva tiene abundantes gránulos grandes, gruesos e irregulares tanto en el margen anterior como a los lados del surco medio anterior, sobre las carenas medias anteriores, quillas superciliares, toda el área entre el surco medio posterior y las quillas centrales laterales dejando sin granulación solo el espacio intercarenal por debajo de los ojos laterales (fig. 10a); mientras que en C. sculpturatus el carapacho tiene granulación fina y en general las quillas son mucho menos fuertes (fig. 10b). Los márgenes y superficie de los terguitos de C. lauriadnae sp.n. están cubiertos por granos grandes y gruesos, mientras que en C. sculpturatus, la granulación es fina. El segmento metasomal V en la nueva especie es 1.3 a 1.5 veces más largo que el carapacho, dorsalmente solo ligeramente acanalado, carenas dorsolaterales vestigiales casi en toda su longitud y solo la parte posterior es aparente mientras que la quilla media ventral es ligeramente aserrada y distinta en toda la longitud del segmento (fig. 10k); en C. sculpturatus es 1.4 o menos veces más largo que el carapacho; con la acanaladura dorsal más profunda, con carenas dorsolaterales similares pero las ventrolaterales y la media ventral son débiles, distintas y finamente granulares (fig. 10j). La vesícula de la nueva especie es casi 2 veces más larga que ancha, mientras que en C. sculpturatus la relación longitud/ancho es cercana a 1.6. Tubérculo subaculear poco desarrollado, ubicado relativamente alejado de la base del aguijón en comparación del tubérculo cónico relativamente grande y cercano a la base del aguijón que se observa en C. lauriadnae sp. n. El conteo de dientes pectinales en la nueva especie es más bajo con 23-25 en machos y 21-24 en hembras por 24-28 y 22-24 respectivamente en C. sculpturatus.
Centruroides pallidiceps es una especie reportada para Sinaloa y Sonora (Ponce-Saavedra et al., 2016; Ponce-Saavedra y Moreno-Barajas, 2005), cuya diferencia es muy clara ya que los adultos tienen muy escasa pigmentación tanto en carapacho como en terguitos (Hoffmann, 1932; Pocock, 1902) y el carapacho con escasas granulaciones gruesas, tanto en hembras como en machos, a diferencia de C. lauriadnae sp. n. cuya pigmentación es muy definida en juveniles y adultos y su carapacho está cubierto por una granulación densa, gruesa en quillas y en espacios intercarenales. Los ejemplares revisados de C. pallidiceps se encuentran decolorados y tienen tonalidad parda cobriza y no se aprecia un patrón de pigmentación; sin embargo, se pueden mencionar otras diferencias morfológicas importantes como: carapacho con el borde anterior ligeramente más cóncavo y en general, mucho menos granuloso, tanto en quillas como en espacios intercarenales, gránulos grandes solo en la parte anterior de las quillas superciliares y centrales laterales (fig. 10c); mano globosa, con carenas muy débiles excepto la externa secundaria que está bien desarrollada y fuerte, pero es completamente lisa (fig. 10f); la carena media de la patela del pedipalpo se aprecia curvada, lisa en su base y posteriormente bien granulosa formada por gránulos gruesos definidos que llegan casi hasta el borde distal donde se une con la quilla dorsointerna; mientras que en la nueva especie es casi recta, completa y bien desarrollada, uniéndose a la dorsointerna antes de llegar al borde distal (fig. 10g, h); en general las quillas metasomales están menos desarrolladas que en la especie nueva; el segmento metasomal V es de tamaño similar al IV, más alto en la región media basal, lo que le da una apariencia algo inflada, es completamente liso con vestigios de granulación en su cara ventral y entre 1.2 y 1.4 veces la longitud del cefalotórax; acanalamiento dorsal ligeramente más profundo que en la nueva especie (fig. 10i). La vesícula es completamente lisa y más globosa que en C. lauriadnae sp. n. al ser más alta, el tubérculo subaculear reducido, espiniforme y comparativamente más alejado de la base del aguijón que en la especie nueva (fig. 11). La base de peines en C. pallidiceps tiene el borde anterior con la muesca media más profunda y evidente que en la especie nueva y un conteo de dientes pectinales de 22-23.

Figura 11. Comparación de vesículas y aguijón en machos de a) Centruroides lauriadnae sp. n., b) C. sculpturatus, c) C. pallidiceps, d) C. suffusus.
Centruroides suffusus, el “alacrán de Durango”, es una especie con distribución conocida en Nayarit, Sinaloa y Zacatecas, además de Durango, cuya coloración general es similar a la nueva especie; sin embargo, hay diferencias notables, como la longitud del metasoma del macho que es 1.8 o más veces más largo que el resto del cuerpo en C. suffusus y de menos de 1.6 veces en la nueva especie. También hay diferencias en ornamentación, ya que las quillas de las manos en C. suffusus son débiles, lisas a ligeramente granulosas con gránulos pequeños y dispersos, a diferencia de las quillas fuertes, bien definidas, con granulación uniforme formada por gránulos bien delimitados y regulares de la nueva especie; los terguitos también presentan diferencias en granulación siendo más fuerte y definida la de C. lauriadnae sp. n. Centruroides suffusus presenta los esternitos completamente lisos, incluyendo las quillas del esternito VII, que son muy débiles, mientras que la nueva especie tiene la superficie de lisa a finamente granulosa y las quillas del esternito VII están bien desarrolladas y crenuladas. La vesícula de C. suffusus es similar a la de la nueva especie, pero difiere en el tubérculo subaculear, el cual en C. lauriadnae sp. n. es cónico y se encuentra cerca de la base del aguijón, mientras que en C. suffusus es reducido y se ubica relativamente alejado de la base del aguijón (fig. 11).

