











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En los últimos años se han publicado diversas investigaciones que documentan la rápida reducción de poblaciones y la extinción de numerosas especies de anfibios a nivel global (Blaustein y Brancroft, 2007; Parrot et al., 2016; Picco y Collings, 2008; Pounds et al., 2008; Wake y Vredenburg, 2008). La problemática de conservación es evidente: el número de especies descritas en el mundo asciende a casi 8,000 (Frost, 2018), de las cuales 29% se encuentra en alguna categoría de riesgo (UICN, 2018). La velocidad con la que se reducen y desaparecen estas poblaciones en todo el mundo es muy acelerada; se ha calculado que su tasa actual de extinción es 211 veces mayor a la estimada a partir del registro fósil (McCallum, 2007). Los países con mayor número de especies en alguna de las categorías de vulnerabilidad son Colombia, Ecuador y México, seguidos por China, EUA, India y Madagascar (IUCN, 2018).
Una de las principales causas de la disminución de las poblaciones de especies de anfibios a nivel global, es la emergencia de una enfermedad dérmica llamada quitridiomicosis, ocasionada por el hongo Batrachochytrium dendrobatidis (Bd; Berger et al., 1998; Carey et al., 2003; Daszak et al., 1999; Kiesecker et al., 2004; La Marca et al., 2005; Lips, 1999; Longcore et al., 1999; Luja et al., 2012; Mendoza-Almeralla et al., 2015; Muñoz-Alonso, 2011). Este ha sido identificado en 56 países y 520 especies de todos los continentes en que existen poblaciones silvestres de anfibios (Global Bd mapping proyect, 2019; Skerratt et al., 2007). En el continente americano existen diversos estudios sobre este patógeno, sin embargo, la mayoría de los registros positivos provienen de EUA (2,259 registros), seguido de Puerto Rico (1,004 registros) y Venezuela (256) (Bonaccorso et al., 2003; Global Bd mapping proyect, 2019; Green y Sherman, 2001; Lips et al., 2003a, b; Ron y Merino-Viteri, 2000). En la región sureste de México, la presencia de Bd ha sido estudiada con frecuencia (Mendoza- Almeralla et al, 2015; Muñoz-Alonso, 2011); sin embargo, por la falta de estudios aún se desconoce su incidencia en los anfibios que habitan la región norte-centro del país. En el norte de México sólo se ha reportado en los estados de Sonora y Baja California (Frías-Álvarez et al., 2008; Lips et al., 2004; Luja et al., 2012; Rollins-Smith et al., 2002). Actualmente, existen registros de Bd en 50 especies de anfibios en México, desde las zonas costeras (Luja et al., 2012) hasta los ecosistemas de montaña (a 2,336 m snm; Frías-Álvarez et al., 2008). Entre las especies con mayor riesgo frente a este patógeno, destacan las que pertenecen a las familias Hylidae y Plethodontidae (Mendoza-Almeralla et al., 2015)
En México habitan 394 especies de anfibios que lo sitúan como el sexto país con mayor diversidad para este grupo (AmphibiaWeb, 2019). Sin embargo, aún es escasa la información sobre la amenaza que este patógeno representa en la región norte-centro del país donde se localiza la “Región Hidrológica Nazas-Aguanaval” (RH36), cuya hidrología ha sido fuertemente alterada por la construcción de obras de captación de las aguas en la parte media y alta de la cuenca. Durante la construcción de las presas no fue estimado el impacto que generó limitar el flujo de agua hacia los ecosistemas presentes en la parte baja de la cuenca, donde habitan organismos como los anfibios que dependen de ella para completar su ciclo vital.
Considerando esta problemática de conservación y el escenario de vulnerabilidad de los anfibios en la región, es importante generar información sobre su estado de riesgo ante la peor enfermedad infecciosa registrada hasta ahora entre los vertebrados (Daszak et al., 2004; Garner et al., 2006). Por lo tanto, en el presente estudio se plantearon objetivos que permitieran conocer la presencia y prevalencia de Bd en anuros desde la región subhúmeda, hasta la zona árida y semiárida del área de estudio. Tomado en cuenta que las condiciones de temperatura y humedad son esencialmente importantes para el establecimiento de este quitridio (Daszak et al., 2003; Drew et al., 2006; Hossack et al., 2009; Pounds et al., 2006), se discuten sus posibles implicaciones en las prevalencias encontradas, las cuales pueden ser de importancia para predecir su dinámica.
Materiales y métodos
La RH36 se localiza en la parte árida y semiárida del norte de México, entre 22°40’ y 26°35’ N, 101°30’ y 106°20’ O, con una superficie de 92,000 km² (Fig. 1). Presenta un régimen de escurrimiento endorreico y está dividida por una zona alta de escurrimiento y una zona baja de acumulación de agua. La RH36 está estructurada por el sistema de sus 2 ríos más importantes, Nazas y Aguanaval, que fluyen de oeste a este y de sur a norte, con un gradiente altitudinal de 3,310 y 2,900 m snm a 1,100 m snm, respectivamente. La desembocadura natural de ambos ríos en la parte baja de la cuenca tenía lugar en las lagunas de Mayrán y Viesca. Sin embargo, su caudal natural ha sido sometido a las obras de aprovechamiento de las aguas y actualmente las aportaciones que reciben las lagunas se limitan a la escasa precipitación y algunos escurrimientos (Descroix et al., 1993). Ubicada en las mesetas continentales del norte de México, la RH36 tiene clima semiárido en la mayoría de su superficie, pero en su parte occidental se apoya en la sierra Madre, que se caracteriza por un clima subhúmedo de montaña. La pluviometría tiene gran variabilidad espacial y disminuye de 900 mm en la parte alta de la cuenca del Nazas, en el parteaguas del Pacífico, a menos de 200 mm en las antiguas lagunas de Viesca y Mayrán. Los tipos de vegetación presentes pueden agruparse en 4 formaciones: bosque, chaparral, matorral xerófilo y pastizal (Tarin-Torres, 1993). La ocupación del suelo por la vegetación natural en toda la RH36 es de 80,024 km2, el resto son áreas dedicadas a las actividades agrícolas (10,182 km2) y superficies sin vegetación aparente (689 km2) (Tarin-Torres, 1993).
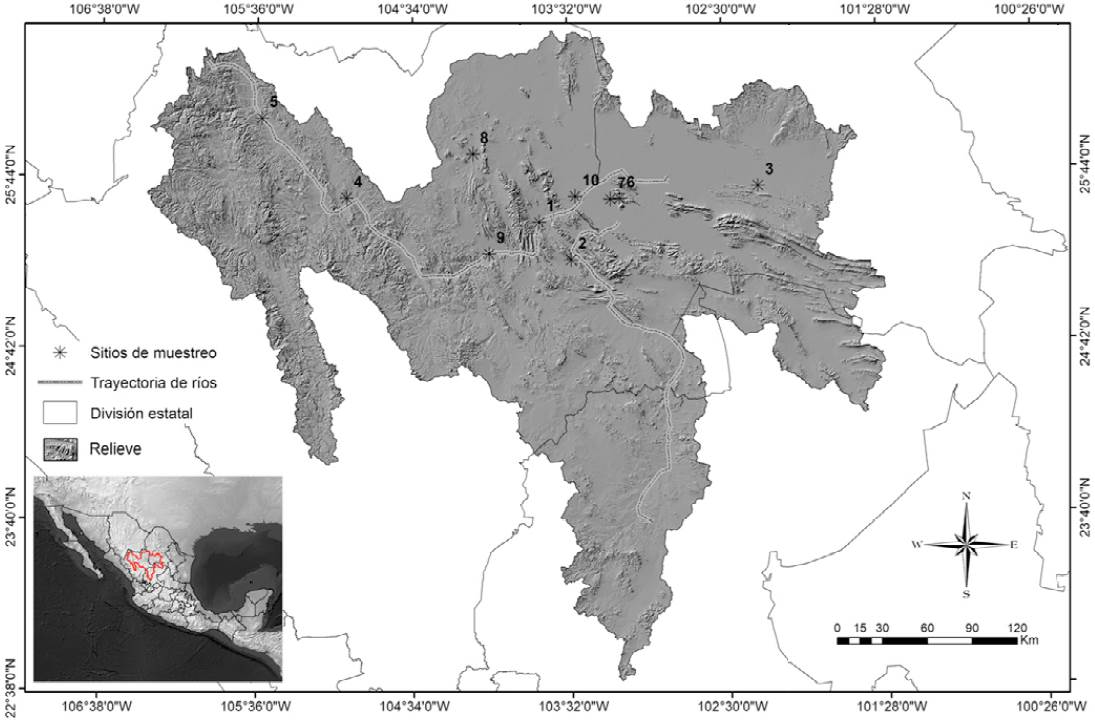
Figura 1 Área de estudio y sitios de muestreo. 1: Parque Estatal Cañón de Fernández, 2: REM SC Jimulco, 3: Parras, 4: presa Lázaro Cárdenas, 5: río Sextín, 6: ejido Vizcaya, 7: La Partida, 8: arroyo La Cadena, 9: Nazas y 10: San Alberto.
Se identificaron 10 sitios con presencia de anfibios, donde se llevaron a cabo muestreos diurnos y nocturnos (Fig. 1). Los anfibios fueron capturados directamente con la mano utilizando guantes de látex, o con redes de golpeo. El método se eligió considerando las características de cada cuerpo de agua (longitud, profundidad y presencia de vegetación en la orilla). Se tomó una muestra dérmica a todos los anfibios capturados siguiendo la metodología no invasiva descrita por Boyle et al. (2004). El hisopo fue deslizado 30 veces en las siguientes partes del cuerpo de los adultos: la superficie inferior o vientre, ingles, patas y membranas interdigitales (telas entre los dedos de los miembros inferiores). El hisopo se colocó en un tubo de rosca estéril y se mantuvo refrigerado hasta su traslado al Laboratorio de Medicina de la Conservación de la Facultad de Ciencias Biológicas de la Universidad Juárez del Estado de Durango, y posteriormente al laboratorio del Grupo de Ecofisiología, Comportamiento y Herpetología (GECOH) de la Universidad de los Andes, en Bogotá, Colombia, donde fueron analizadas.
El análisis molecular de las muestras se realizó bajo el protocolo descrito por Annis et al. (2004) con algunas modificaciones. Para realizar la extracción de ADN cada hisopo se lavó con 500µl de etanol a 96% y se aplicó vórtex a cada muestra por 40 segundos cambiando constantemente el tubo de posición. Con la ayuda de unas pinzas, se escurrió el hisopo contra la pared del tubo. El hisopo fue desechado y el etanol se transfirió a un tubo de 0.6 ml. Las pinzas fueron esterilizadas cada vez que se cambió de muestra para evitar contaminación. Las muestras se centrifugaron por 10 minutos a 12,000 rpm hasta observar un pellet en el fondo del tubo. Después de centrifugar la muestra, el sobrenadante fue descartado y se colocó por 40 minutos en un Speedvac a 30 ºC para evaporar el etanol restante y así evitar inhibición de la reacción de PCR. Posteriormente, se agregaron 10 µl de agua libre de nucleasas y se llevó de nuevo al vórtex para resuspender el pellet. Para concluir la extracción de ADN, la muestra fue transferida a un nuevo tubo de 0.2 ml, donde se añadieron 12 µl de GeneReleaser® (BioVentures) y posteriormente fue llevada al termociclador con el protocolo sugerido por el proveedor. El control positivo se preparó agregando 10 µl de agua libre de nucleasas y una muestra tomada de un cultivo puro de Bd aislado en agar TGH (Tryptone Gelatin Hydrolysate). Para el control negativo se utilizaron solo 10 µl de agua libre de nucleasas. Al finalizar el protocolo las muestras fueron centrifugadas durante 3 minutos a 12,000 rpm. El sobrenadante (la fase transparente) fue transferido a un nuevo tubo de 0.2 ml etiquetado con el número de colecta. La extracción se almacenó a -20 ºC para luego ser utilizada en la PCR.
Para la PCR se usaron reacciones de 12 µl; encadatubo se agregaron 6 µl de GoTaq Green Master Mix (Promega), 0.5 µl de cada primer (Bd1a 5’CAGTGTGCCATATGTCACG3’y Bd2a 3’CATGGTTCATATCTGTCCAG5’; Annis et al., 2004), 3 µl de agua libre de nucleasas y 2 µl de la muestra de ADN. Se utilizó el mismo método para los controles positivo y negativo. Después de servir todas las muestras, se aplicó vórtex y se colocaron en la centrifuga (spin) durante unos segundos para precipitar el total de la muestra. La reacción se llevó a cabo en un termociclador BioRad® C1000 con placas de 60 pocillos, utilizando el programa de amplificación descrito por Annis et al. (2004). Los productos de PCR fueron separados por electroforesis en un gel de agarosa al 1% en una cámara Mini-Sub GT de BioRad® a 40 voltios y 110 MA durante 90 minutos. El revelado de los geles se llevó a cabo en un transiluminador Gel Doc XR+ de BioRad©. Se buscó una banda cercana a los 300 pb de longitud en las muestras positivas para Bd.
La prevalencia por especie y por localidad se calculó en R 3.2.0 (R Core Team, 2017) como una proporción de individuos con resultados positivos con respecto al número de muestras por localidad y por especie. Se calcularon intervalos de confianza (95%) con base en una distribución binomial para todas las estimaciones de prevalencia a través del intervalo de Wilson (Paquete binom; Dorai-Raj, 2009; Hughey et al., 2014).
Resultados
Se realizaron en total 12 muestreos en 10 localidades, donde se recolectaron 169 muestras dérmicas de 7 especies de anfibios en estado adulto. Se obtuvieron muestras de Anaxyrus punctatus, A. debilis, A. cognatus, Scaphiopus couchii, Gastrophryne olivacea, Lithobates berlandieri y L. catesbeianus. Como resultado de los análisis de PCR, se obtuvieron 52 bandas positivas para Bd, lo cual equivale a 30% del total de muestras recolectadas. Entre paréntesis, acompañando a los porcentajes obtenidos, se presenta el intervalo de confianza. La localidad con mayor prevalencia fue el Ejido Vizcaya con 80% (62-91), seguido del Parque Estatal Cañón de Fernández, con 40% (23-61). La Presa Lázaro Cárdenas presentó resultados positivos, con 37% (18-61) de las muestras, seguida por la Reserva Ecológica Municipal Sierra y Cañón de Jimulco (REM SC Jimulco) con 21% (13-33). La localidad de Río Sextín presentó una prevalencia de 40% (11-76), con únicamente 5 muestras. En algunas localidades el número de muestras fue reducido, por lo que los resultados negativos para Bd no son concluyentes.
A nivel de especie, la mayor prevalencia fue observada en A. punctatus con 80% (62-91). Las muestras de esta especie fueron recolectadas en la localidad de Ejido Vizcaya. En segundo lugar, se encontró a A. cognatus en la REM SC Jimulco y en tercer lugar, a L. berlandieri en la Presa Lázaro Cárdenas con prevalencias de 57 (36-75) y 42% (21-67), respectivamente. Lithobates catesbeianus, con únicamente 4 muestras, presentó una prevalencia de 75% (30-95) en la localidad del Parque Estatal Cañón de Fernández (Tabla 1).
Tabla 1 Prevalencia de Bd por especie y localidad de muestreo. La prevalencia está dada como la proporción de individuos infectados en relación con el tamaño de la muestra. Entre paréntesis se muestra el intervalo de confianza de Wilson (95%). Bd+: Número de resultados positivos para Bd; Bd+ (%): porcentaje de resultados positivos con respecto al número de muestras.
| Localidad | Ubicación | Elevación (m snm) |
Especie | n | Bd+ | Bd+ (%) |
Prevalencia por especie |
Prevalencia por localidad |
|
|---|---|---|---|---|---|---|---|---|---|
| X | Y | ||||||||
| Parras | -102.27201 | 25.6597 | 1,141 | S. couchii | 1 | 0 | 0 | 0 (0-80) | 0 (0-20) |
| G. olivacea | 14 | 0 | 0 | 0 (0-21) | |||||
| Parque Estatal Cañón de Fernández |
-103.72957 | 25.455 | 1,165 | L. berlandieri | 17 | 6 | 35 | 35 (17-58) | 40 (23-61) |
| L. catesbeianus | 4 | 3 | 75 | 75 (30-95) | |||||
| G. olivacea | 1 | 0 | 0 | 0 (0-80) | |||||
| San Alberto | -103.48905 | 25.6085 | 1,125 | S. couchii | 10 | 0 | 0 | 0 (0-27) | 0 (0-27) |
| REM SC Jimulco | -103.51951 | 25.228 | 1,231 | A. cognatus | 21 | 12 | 57 | 57 (36-75) | 21 (13-33) |
| A. debilis | 20 | 1 | 5 | 5 (0-23) | |||||
| G. olivacea | 21 | 1 | 4 | 4 (0-22) | |||||
| L. berlandieri | 2 | 0 | 0 | 0 (0-65) | |||||
| Ejido Vizcaya | -103.19434 | 25.5929 | 1,113 | A. punctatus | 26 | 21 | 80 | 80 (62-91) | 80 (62-91) |
| La Partida | -103.25417 | 25.5895 | 1,111 | L. berlandieri | 8 | 0 | 0 | 0 (0-32) | 0 (0-32) |
| Nazas | -104.06102 | 25.2652 | 1,229 | A. cognatus | 1 | 0 | 0 | 0 (0-80) | 0 (0-65) |
| L. catesbeianus | 1 | 0 | 0 | 0 (0-80) | |||||
| Presa Lázaro Cárdenas | -105.00805 | 25.6066 | 1,445 | L. berlandieri | 14 | 6 | 42 | 42 (21-67) | 37 (18-61) |
| A. punctatus | 2 | 0 | 0 | 0 (0-65) | |||||
| Río Sextín | -105.57091 | 26.0886 | 1,655 | L. berlandieri | 5 | 2 | 40 | 40 (11-76) | 40 (11-76) |
| Arroyo La Cadena | -104.16634 | 25.8689 | 1,518 | L. berlandieri | 1 | 0 | 0 | 0 (0-80) | 0 (0-80) |
Discusión
Las prevalencias de Bd registradas en las especies A. cognatus, A. debilis, A. punctatus y G. olivacea, constituyen los primeros registros positivos de esta infección reportados en la literatura científica. Estudios anteriores como el de Ouellet et al. (2005) reporta prevalencia de cero en ejemplares de A. cognatus y A. punctatus recolectados en EUA y México, respectivamente. En todas las muestras recolectadas de Scaphiopus couchii, se obtuvieron resultados negativos para Bd, de forma similar que en un estudio previo desarrollado por Goka et al. (2009). Respecto a G. olivacea y A. debilis, no se hallaron estudios sobre esta enfermedad que precedan a este estudio.
Para Lithobates berlandieri, solo se encontraron 2 reportes de quitridiomicosis publicados: Muñoz-Alonso (2011), que menciona 13 individuos de esta especie infectados con un promedio de 57 esporas por individuo y Cabrera-Hernández (2012) que señala una prevalencia de 50% con únicamente 2 muestras (ambos reportes realizados para poblaciones del estado de Chiapas, México). Durante el presente estudio se recolectaron 47 muestras de ejemplares adultos de L. berlandieri, correspondientes a 6 localidades. La prevalencia más alta se encontró en la presa Lázaro Cárdenas con 42% (21-67). Las muestras recolectadas en la localidad de La Partida fueron de ejemplares sin vida, sin embargo, estas muestras resultaron negativas para Bd, sugiriendo que la muerte de estos ejemplares se dio por otros factores.
La mayor prevalencia (80% [62-91]) se observó en la población de A. punctatus presente en el ejido Vizcaya. Los individuos de esta población se encontraron con apariencia saludable, lo cual podría explicarse por la hipótesis que plantean las variables climáticas externas como limitantes para el desarrollo de la infección (Hughey et al., 2014), junto con otras defensas naturales de los anfibios contra Bd. Entre estas variables se ha mencionado la presencia de bacterias antagonistas capaces de inhibir el crecimiento de este hongo (Harris et al., 2006, 2009; McKenzie et al., 2012). Asimismo, existen reportes que demuestran la coexistencia de Bd con poblaciones de Bufo boreas sin causar declinaciones rápidas aparentes (Pilliod et al., 2009), o su presencia como especie enzoótica en poblaciones sanas del este de Norteamérica (Ouellet et al., 2005). Estas hipótesis respecto a la resistencia hacia Bd que pudiera existir en algunas especies de anfibios, generan interés sobre los resultados obtenidos en este estudio para A. punctatus. Asimismo, destaca el interés por evaluar a detalle el estado de riesgo de otras especies como A. debilis, G. olivacea y L. berlandieri, sujetas a protección especial (Pr) de acuerdo con la Norma Oficial Mexicana 059 (Semarnat, 2010). En este aspecto, los resultados obtenidos en esta investigación pueden ser de utilidad en el establecimiento de prioridades, considerando que los descensos poblacionales debidos a esta enfermedad una vez en marcha, son difíciles de detener (Bielby et al., 2013).
La ubicación de los registros positivos encontrados denota importancia en las 3 áreas naturales protegidas de la región, como la REM SC de Jimulco y el Parque Estatal Cañón de Fernández (ubicados en la parte media de la cuenca), donde se concentran los esfuerzos de conservación y donde el riesgo epidemiológico de los anfibios debido a esta enfermedad es poco conocido.
Se desconoce si las condiciones climáticas de la región están limitando el potencial patogénico de este hongo, o si otras variables importantes para su desarrollo como el pH del agua (Piotrowski et al., 2004) o el microhábitat utilizado por los anfibios (Ron, 2005) juegan un papel importante. Por lo tanto, ampliar los estudios en la zona resulta primordial para identificar las poblaciones sensibles y sus factores de riesgo, así como para evaluar la necesidad de implementar estrategias encaminadas hacia la conservación de la biodiversidad de anfibios de la región y los ecosistemas donde habitan, o al monitoreo de este patógeno y su dinámica inter o intraespecífica.
Algunos estudios sugieren que las limitaciones fisiológicas de Bd pueden condicionar su distribución en los hábitats más áridos o con temperaturas no óptimas para su crecimiento (Forrest y Schlaepfer, 2011; Lannoo et al., 2011). Por lo tanto, estos sitios a menudo se consideran de menor riesgo para los anfibios. Sin embargo, en los resultados de esta investigación, la prevalencia más elevada se observó en una población de A. punctatus que habita en la localidad ejido Vizcaya, ubicada en la parte árida de la cuenca con una precipitación anual promedio de 204 mm y un rango de temperatura anual entre 4 y 36 °C (Descroix et al., 1993). Esta localidad podría considerarse no apta para el establecimiento del quitridio tomando en cuenta los datos de Piotrowski et al. (2004), que sugieren su crecimiento en un rango de temperatura de 4-25 °C, y su muerte luego de una exposición prolongada a más de 29 °C. La presencia del quitridio en esta zona podría explicarse bajo la hipótesis de otros autores que asocian la incidencia de esta enfermedad con la variación estacional de la temperatura que puede favorecer su crecimiento en ciertas temporadas del año, aunque en invierno esté por debajo de su óptimo y en verano por encima de él. También se ha encontrado una relación entre estas variaciones estacionales y la prevalencia de la enfermedad (Bielby et al., 2013; Hughey et al., 2014; Sapsford et al., 2013).
Asimismo, Ron (2005) menciona que el rechazo activo a las altas temperaturas por parte del hospedero, que puede refugiarse en sitios más frescos, favorecería el crecimiento de Bd. Dicho comportamiento se observa comúnmente en los anfibios de la región estudiada, sobre todo en los del género Anaxyrus que habitan los sitios más áridos. Otros datos sugieren que las poblaciones de anfibios de zonas montañosas (frías y elevadas) tienen mayor riesgo de sufrir declinaciones, debido a que en estas condiciones su sistema inmune decrece y el potencial patogénico de Bd es mayor (Berger et al., 2004; Bielby et al., 2013; Bradley et al., 2002; Puschendorf et al., 2009; Rollins-Smith et al., 2002; Sapsford et al., 2013).
Lamentablemente, los registros limitados obtenidos en este trabajo para zonas altas (5 localidades que van de los 1,000 a 1,700 m snm), con respecto a la extensión total del área de estudio, no permiten analizar la posible influencia de las variables climáticas en la prevalencia de este patógeno. No obstante, considerando la hipótesis de Sapsford (2013), se puede sugerir que la presencia de este patógeno en las partes más altas pudo favorecer su dispersión hacia las zonas bajas, debido a las conexiones hidrológicas. Este mismo autor señala que los reservorios pueden ser importantes en la persistencia de esta enfermedad en zonas bajas, concepto interesante al considerar la presencia de la rana toro norteamericana (L. catesbeianus) en el área de estudio, descrita como un portador asintomático de esta enfermedad (Daszak et al., 2004). Durante este estudio, la prevalencia en la rana toro fue de 75% (30-95), en la localidad de Parque Estatal Cañón de Fernández. Esta prevalencia es similar a la reportada por otros autores como Sánchez et al. (2008): 79.9% (73.4-86.3). Debido al número limitado de muestras recolectadas y a la ausencia de estudios poblacionales, aún no es posible conocer el alcance de esta especie exótica como vector o reservorio del quitridio en el área de estudio. Si bien, durante la toma de muestras la rana toro fue observada cerca de otras especies nativas, como L. berlandieri y G. olivacea, no existen estudios que permitan entender el nivel de traslape entre éstas, así como el papel que pudiera desempeñar la rana toro en la transmisión de esta enfermedad.
El desconocimiento general de la dinámica poblacional de los anuros regionales y las fuertes alteraciones que se han realizado en los últimos años sobre el régimen hidrológico de la cuenca (principalmente por la construcción de presas río arriba para la captación de agua con interés agrícola), genera preocupación sobre el riesgo de estos organismos frente a Bd. Por lo tanto, es necesario llevar a cabo estudios tanto inter como intraespecíficos que permitan tener una idea del estado de conservación actual en sus poblaciones, así como evaluar, en escala más fina, el estado en que se encuentran los ecosistemas donde habitan. La perspectiva hidrológica presentada en este trabajo, puede ser de utilidad para estudios posteriores sobre la ecología de este patógeno en la región y su relación con variables bioclimáticas.

