











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Existe un proceso de pérdida de la biodiversidad a nivel global sin precedentes, causado por el impacto de las actividades humanas (Ceballos et al., 2015). Si bien se requiere de una amplia gama de iniciativas de conservación y manejo para la mitigación de este fenómeno, las áreas naturales protegidas (ANP) siguen siendo una de las estrategias más importantes para lograrlo. El fin último de las ANP es preservar la biodiversidad que albergan y los procesos ecológicos que la mantienen (Conanp, 2018). Sin embargo, alcanzar este objetivo resulta cada vez más complejo debido a los efectos acumulados y sinérgicos de factores como: la deforestación, la fragmentación del hábitat, los fuegos provocados, la minería, la cacería y el cambio climático (Ruiz-Montoya et al., 2017). Resulta entonces de primordial importancia evaluar continuamente en qué medida las ANP están cumpliendo su objetivo. Para hacer esto, se requiere contar con una línea base de información detallada que permita aplicar metodologías de monitoreo como los indicadores de Presión-EstadoRespuesta en donde se detectan los factores de presión que están poniendo en riesgo elementos biológicos específicos, se diseña una estrategia para monitorearlos y en el caso de que sea necesario, se establecen medidas de mitigación (Villaseñor y Botello, 2016). Lamentablemente, en la mayoría de los casos se carece de información detallada que permita evaluar la respuesta de poblaciones y comunidades biológicas al impacto antrópico (Ahumada et al., 2011). En el caso de la fauna de mamíferos silvestres la carencia de información detallada sobre atributos de sus poblaciones y comunidades se relaciona en gran medida con la dificultad de registrar su presencia en sus hábitats naturales. Afortunadamente, el uso cada vez más generalizado de herramientas como las fototrampas ha permitido subsanar paulatinamente esta carencia de información (Rowcliffe et al., 2008; Tobler et al., 2008). No obstante, aun cuando las fototrampas han demostrado ser un método muy eficiente para detectar la presencia de una amplia variedad de animales, existe la posibilidad de que algunas especies no sean detectadas debido a variaciones naturales en su abundancia o patrones de actividad. Por ejemplo, especies como los pecaríes de labios blancos (Tayassu pecari) realizan migraciones frecuentes por lo que en una temporada pueden registrarse en un sitio de muestreo y a la siguiente no (Fragoso, 2004). Por esta razón es muy importante contar con monitoreos de la fauna silvestre que abarquen varias temporadas e incluso años de muestreo para tener una descripción más sólida de las características de sus poblaciones y comunidades.
El impacto antrópico sobre la biodiversidad se manifiesta de distintas maneras, una de ellas es la disminución de la abundancia de las poblaciones de mamíferos silvestres que puede llegar hasta su extirpación o al extremo, hasta la extinción de especies y de los procesos ecológicos que sustentan (defaunación sensuDirzo, 2001). Si bien existe evidencia de que la defaunación está afectando un número creciente de especies con características diversas (Ceballos et al., 2015), existen especies que son particularmente sensibles debido a que combinan algunos de los siguientes atributos: requerimientos de hábitat muy específicos, áreas de acción extensas, tasas reproductivas bajas y gran atractivo para los cazadores (Ojasti, 2000). En el neotrópico, este grupo de especies incluye felinos como el jaguar (Panthera onca) y herbívoros de gran talla corporal como el tapir centroamericano (Tapirella bairdii). Por otra parte, existen especies de mamíferos que pueden responder de manera positiva a la perturbación humana porque son beneficiadas por las condiciones que se generan en los ambientes alterados (p.ej., ausencia de competidores y depredadores o incremento de alimento; Galetti et al., 2015) y porque se caracterizan por tener altas tasas reproductivas. De esta manera, la perturbación antrópica puede causar cambios en la composición de las comunidades de mamíferos silvestres que, además de reducir su diversidad, puede provocar alteraciones en procesos ecológicos importantes para el funcionamiento de los hábitats naturales (Camargo-Sanabria et al., 2015).
La Reserva de la Biosfera Selva El Ocote (REBISO) se localiza dentro del “hotspot” de biodiversidad de Mesoamérica y constituye uno de los pocos refugios existentes en el país para poblaciones de mamíferos silvestres de talla mediana y grande (Semarnat-Conanp, 2001). La REBISO juega un papel fundamental para mantener la conectividad del hábitat entre los bosques tropicales localizados en la península de Yucatán y los ubicados en Oaxaca (Mendoza et al., 2013). A pesar de esto, la región ha sido poco estudiada en términos de su diversidad de vertebrados. Se estima que, dadas sus características, la REBISO podría albergar cerca de 138 especies de mamíferos (23% del total para México; Semarnat-Conanp, 2001). El alcanzar un mejor conocimiento de la biodiversidad que alberga esta reserva es urgente debido al impacto que diversas actividades humanas tienen en la región. Por ejemplo, la REBISO ha sido fuertemente afectada por incendios forestales de distintas magnitudes, provocados en buena medida por el uso de prácticas agrícolas deficientes, siendo uno de los más importantes el ocurrido en 1998, que afectó 19,000 ha y que se asoció con una prolongada sequía debida al efecto del fenómeno de El Niño. De manera posterior a este incendio, la tasa de deforestación anual aumentó de 0.21% (periodo 1986-1995) a 2.54% (periodo 1995-2000) (Flamenco-Sandoval et al., 2007). Además, en la reserva existe cacería ilegal para subsistencia de especies como el tepezcuintle (Cuniculus paca), guaqueque (Dasyprocta mexicana) y el pecarí de collar (Dicotyles crassus). Esta serie de problemáticas se han agravado debido a la invasión por personas en porciones de la REBISO, como la zona núcleo II (Navarrete-Gutiérrez et al., 1996; Pozo-Montuy obs. pers.).
En este estudio nos centramos, como una primera aproximación, en los impactos que la actividad antropogénica tiene sobre la composición de la comunidad de mamíferos medianos y grandes (> 500 g) de la REBISO y su área de influencia. En particular los objetivos del estudio fueron: a) realizar un muestreo multianual (2013-2016) de la fauna de mamíferos medianos y grandes presentes en la REBISO y su área de influencia mediante fototrampeo; b) comparar la riqueza y composición de la comunidad de mamíferos obtenida a partir del muestreo multianual vs. el conjunto de especies cuya presencia era reportada para la REBISO en 1996, con base en inventarios faunísticos, y c) analizar las diferencias en la composición y frecuencia de captura de las especies presentes en la zona núcleo, la zona de amortiguamiento y el área de influencia de la REBISO. Nuestra expectativa es que si la REBISO está cumpliendo con su función de preservar la biodiversidad local deberían de evidenciarse pocos cambios en la riqueza y composición de especies de mamíferos medianos y grandes registradas en el muestreo multianual vs. lo reportado hasta 1996 para la reserva. Por otra parte, se esperaría encontrar diferencias entre la riqueza y la composición de la fauna entre la zona núcleo y las zonas de amortiguamiento e influencia, con la zona núcleo mostrando una mayor presencia de especies sensibles a la perturbación (p.ej., jaguar, tapir y temazate) y las zonas de amortiguamiento e influencia mostrando una mayor presencia de especies generalistas (p.ej., coaties, zorrillos, etc.).
Materiales y métodos
La Reserva de la Biosfera Selva El Ocote se localiza en el sureste de México, en la porción occidental de Chiapas, entre las coordenadas 16°45’42” y 17°09’00” N, -93°54’19” y -93°21’20” O). La REBISO tiene una superficie total de 101,288 ha distribuidas entre los municipios de Ocozocoautla de Espinosa, Cintalapa de Figueroa, Tecpatán de Mezcalapa y Jiquipilas (Semarnat-Conanp, 2001). Cuenta con 2 zonas núcleo y una zona de amortiguamiento. La zona núcleo I tiene una extensión de 30,648 ha, la zona núcleo II 9,783 ha y la zona de amortiguamiento 60,856 ha. Además, existe un área de influencia en donde se establece una estrecha interacción social, económica y ecológica entre la REBISO y las comunidades locales vecinas. El área de influencia tiene una superficie de 124,531.44 ha, formando un polígono alrededor de la REBISO, el cual se definió utilizando como límites las microcuencas de los ríos: El Tigre, Cacahuano, El Encajonado, Zona Kárstica, Nezahualcoyotl, Selva Zoque, Negro, Cárdenas, La Venta, Las Láminas, El Francés, Ocuilapa y La Lucha. Además de las microcuencas, se tomaron en cuenta los tipos de tenencia de la tierra para hacer la delimitación (Fig. 1; Semarnat-Conanp, 2001; Promaanp, 2012). Estas 3 zonas (núcleo, amortiguamiento y área de influencia) difieren en la composición y estado de conservación de su vegetación y grado de colonización (Tabla 1; Promaanp, 2012). Siendo la zona núcleo la más conservada respecto a la zona de amortiguamiento, y el área de influencia la más perturbada con una mayor población humana y número de asentamientos.
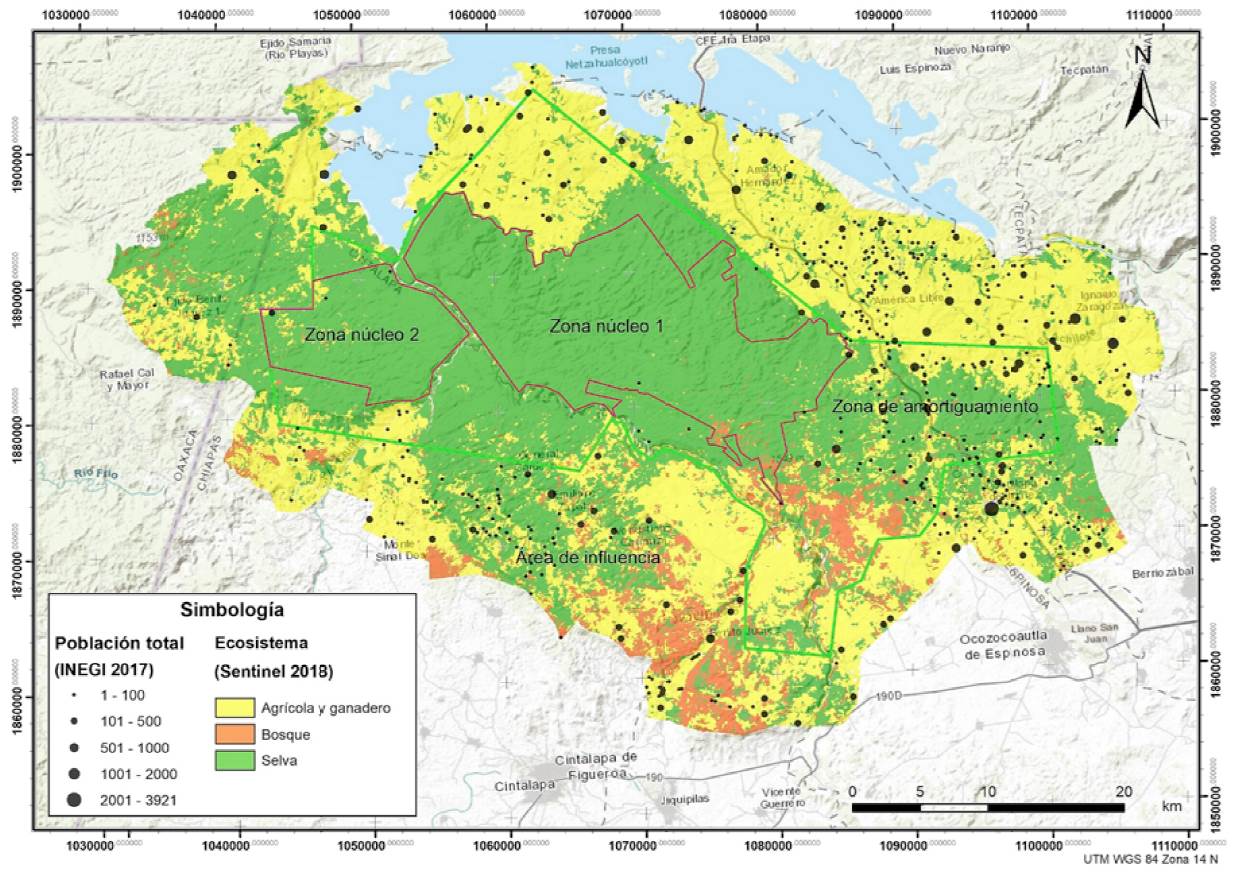
Figura 1 Mapa de ubicación y zonificación de la Reserva de la Biosfera Selva El Ocote en donde se muestra la extensión de diferentes usos de suelo como selvas, bosques y zonas agrícolas y ganaderas, así como la distribución de asentamientos humanos como indicadores del grado de conservación/perturbación. Elaborado por Irving Castañeda Guerrero y Oscar Godínez-Gómez.
Tabla 1 Indicadores del estado de conservación de las zonas núcleo, de amortiguamiento y de influencia de la Reserva de la Biosfera Selva El Ocote. Elaboración a partir de datos cartográficos de Promaanp (2012).
| Indicador | Zona núcleo |
Zona de amortiguamiento |
Área de influencia |
|---|---|---|---|
| Porcentaje de superficie cubierta por selvas medianas y altas | 97.5 | 55.9 | 29.8 |
| Porcentaje de superficie cubierta por zonas agrícolas y ganaderas | 1.7 | 37.4 | 61.9 |
| Porcentaje de superficie cubierta por bosques | 0.7 | 5.7 | 6.7 |
| Población humana (número de habitantes) | 396 | 7,621 | 33,558 |
| Número de asentamientos humanos | 5 | 133 | 391 |
La REBISO tiene una variación altitudinal que va de los 180 m hasta los 1,500 m, presentando mesetas kársticas y laderas con pendientes abruptas. El clima dominante de la reserva es cálido húmedo Am(f) con una temperatura media anual de 22 °C y una temperatura del mes más frío de 18 °C; se presentan abundantes lluvias en verano teniendo una precipitación anual que varía entre los 1,500 y 2,500 mm (García, 1988).
Para establecer la composición histórica de la fauna de mamíferos medianos y grandes (> 500 g) de la REBISO, se recopiló información de inventarios faunísticos entre 1991 y 1995. La información que conforma esta línea base, publicada en 1996, fue generada por observaciones directas, recolecta de ejemplares y registros de huellas y restos (Navarrete-Gutiérrez et al., 1996). Del total del listado de especies, se consideraron solo aquellas que fueran factibles de ser detectadas mediante fototrampeo (i.e., terrestres o con actividad principalmente en el piso de la selva). A partir de esta revisión, se obtuvo un listado de 24 especies de mamíferos medianos y grandes reportadas para la REBISO, entre las que se incluyen 5 de las 6 especies de felinos existentes en el país y el tapir centroamericano (Tabla 2). Este listado se utilizó para hacer la comparación contra los resultados del muestreo multianual. Los nombres científicos de las especies se actualizaron basándose en la taxonomía propuesta por Ramírez-Pulido et al. (2014).
Tabla 2 Especies de mamíferos medianos y grandes reportadas para la Reserva de la Biosfera Selva El Ocote por Navarrete-Gutiérrez et al. (1996) y especies registradas durante este estudio. Se incluye información de los gremios alimentarios y el rango de peso de individuos adultos de cada especie de acuerdo a la Conabio (2018). Taxonomía basada en Ramírez-Pulido et al. (2014).
| Familia/Especie | Clave | Gremio alimentario |
Reportada originalmente |
Este estudio |
Rangos de peso (kg) |
|---|---|---|---|---|---|
| Procyonidae Bassariscus sumichrasti | Basu | Omnívoro | 1 | 0 | 0.6-1.6 |
| Nasua narica | Nana | Omnívoro | 1 | 1 | 4.1-5.4 |
| Procyon lotor | Prolo | Omnívoro | 1 | 0 | 5.5 -15 |
| Mephitidae Conepatus leuconotus* | Cole | Insectívoro | 1 | 0 | 1.5-2.7 |
| Conepatus semistriatus | Cose | Insectívoro | 0 | 1 | 1.4-3.5 |
| Cuniculidae Cuniculus paca | Cupa | Frugívoro | 1 | 1 | 7-10 |
| Agoutidae Dasyprocta mexicana | Dame | Frugívoro | 1 | 1 | 1.3-2.5 |
| Dasypodidae Dasypus novemcinctus | Dano | Insectívoro | 1 | 1 | 4-8 |
| Didelphidae Didelphis marsupialis | Dima | Omnívoro | 1 | 1 | 2.0-4 |
| Didelphis virginiana | Divi | Omnívoro | 1 | 1 | 1.9- 2.1 |
| Philander opossum | Phop | Omnívoro | 1 | 0 | 0.7-1.4 |
| Mustelidae Eira barbara | Eiba | Carnívoro | 1 | 1 | 2.7- 7 |
| Galictis vittata** | Gavi | Carnívoro | 1 | 1 | 1.5-3.2 |
| Felidae Leopardus pardalis | Lepa | Carnívoro | 1 | 1 | 7.8-14 |
| Leopardus wiedii | Lewi | Carnívoro | 1 | 1 | 2.6-5 |
| Panthera onca | Paon | Carnívoro | 1 | 1 | 45.0- 114 |
| Puma concolor | Puco | Carnívoro | 1 | 1 | 34-80 |
| Herpailurus yagouaroundi | Heya | Carnívoro | 1 | 0 | 3.5-6.5 |
| Cervidae Mazama temama | Mate | Herbívoro | 1 | 1 | 12-32 |
| Odocoileus virginianus | Odvi | Herbívoro | 1 | 1 | 60-160 |
| Tayassuidae Dicotyles crassus | Dicra | Herbívoro | 1 | 1 | 17.0-21.6 |
| Leporidae Sylvilagus floridanus | Syflo | Herbívoro | 1 | 0 | 1- 1.2 |
| Mirmecophagidae Tamandua mexicana | Tame | Insectívoro | 1 | 1 | 3.8- 8.5 |
| Tapiridae Tapirella bairdii | Taba | Herbívoro | 1 | 1 | 180- 300 |
| Canidae Urocyon cinereoargenteus | Urci | Omnívoro | 1 | 1 | 1.8- 3.5 |
| Total | 24 | 19 |
*Navarrete et al. (1996) lo reportan como C. mesoleucus, **sinonimia de G. allamandi
Los muestreos se llevaron a cabo en los siguientes periodos: septiembre de 2013 a marzo de 2014, septiembre de 2014 a marzo de 2015, julio de 2015 a febrero de 2016 y septiembre de 2016 a enero de 2017. En el primer periodo se establecieron 9 fototrampas en la zona núcleo y una en la zona de amortiguamiento. En 2014- 2015 se colocó una fototrampa en el área de influencia y 2 en la zona de amortiguamiento. En el tercer periodo (2015-2016) se aumentó el número de estaciones de fototrampeo a 14, que fueron repartidas así: una en el área de influencia, una en la zona de amortiguamiento y 8 en la zona núcleo. Finalmente, en el periodo 2016-2017 se instalaron 10 fototrampas: una en el área de influencia, una en la zona de amortiguamiento y 8 en la zona núcleo (Tabla 3). En algunas ocasiones, las fototrampas fueron colocadas en los mismos sitios que el periodo previo.
Tabla 3 Esfuerzo de muestreo (días cámara-trampa) y número de cámaras (indicado entre paréntesis) colocadas en cada periodo de muestreo y zona de la Reserva de la Biosfera Selva El Ocote.
| Zona | Área de influencia | Zona de amortiguamiento | Zona núcleo | Esfuerzo de muestreo |
|---|---|---|---|---|
| 2013-2014 | 0 | 48 (1) | 745 (9) | 793 (10) |
| 2014-2015 | 197 (1) | 254 (2) | 0 | 451 (3) |
| 2015-2016 | 200 (3) | 405 (4) | 454 (7) | 1,059 (14) |
| 2016-2017 | 30 (1) | 80 (1) | 422 (8) | 532 (10) |
| Total | 427 (5) | 787 (8) | 1,621 (24) | 2,835 |
Las fototrampas se colocaron en sitios de paso y senderos de la fauna, siempre manteniendo una separación mínima entre ellas de 1 km. Se utilizaron cámaras Bushnell Trophy Camp HD y Cuddeback Black Flash E3 de 20 mega pixeles de resolución. Estos equipos se programaron para tomar una serie de 3 fotografías seguida por un video de 10 s. Su sensor se ajustó a nivel medio y se establecieron 3 minutos de inactividad entre detecciones. Estas cámaras operaron de manera continua las 24 h y se colocaron a una altura que varió de 60 a 80 cm, dependiendo de la pendiente del terreno. Las cámaras fueron revisadas mensualmente para verificar la carga de baterías y descargar la información de las memorias. Cada una de las imágenes registradas se asoció con los datos de la hora, fecha, temperatura y fase lunar correspondientes.
Se organizaron las imágenes en una base de datos con los siguientes campos: nombre científico de la especie registrada, número de estación de muestreo, fecha y hora de recolecta, tipo de vegetación y zona donde fue registrada (núcleo, amortiguamiento y de influencia). Para evitar contar cada imagen de una misma especie como un registro independiente, se agruparon las imágenes que fueron obtenidas en la misma estación y que fueron de la misma especie dentro de intervalos de 24 h (de aquí en adelante evento). Se calculó la frecuencia de captura (FC) para cada especie usando la siguiente fórmula: número de eventos/esfuerzo de muestreo × 100.
Para comparar la riqueza de especies registrada en los distintos periodos de muestreo, se construyeron curvas de rarefacción basadas en muestras con intervalos de confianza de 95%, usando el paquete iNEXT (Hsieh et al., 2014). Estas curvas de rarefacción se construyeron con base en el índice de diversidad verdadera de orden q = 0 ( 0 Dγ), que es equivalente a la riqueza de especies (Chao et al., 2014). Las comparaciones se hicieron con base en el esfuerzo de muestreo más pequeño pero también extrapolando al esfuerzo de muestreo más grande. Se aplicó este mismo enfoque de análisis para comparar la diversidad de la comunidad de mamíferos entre la zona núcleo de la REBISO, su zona de amortiguamiento y el área de influencia en el periodo 2015-2016, que fue el que tuvo un esfuerzo de muestreo más equitativo entre zonas (200 días-cámara en el área de influencia, 405 días-cámara en la zona de amortiguamiento y 454 días-cámara en la zona núcleo). Asimismo, se comparó la composición de la comunidad de mamíferos entre estas 3 zonas mediante curvas de rango-abundancia realizadas con base en la frecuencia de captura de cada especie y mediante un escalamiento multidimensional no-métrico y un análisis de similitud (Anosim), este último usando la función metaMSD del paquete Vegan (Oksanen et al., 2013). El escalamiento calcula un valor de estrés, escalado entre 0 y 1, que mide qué tan lejanas son las distancias en el espacio multidimensional a las distancias originales en la matriz de asociación (Borcard et al., 2011).
Finalmente, para tener una visión general de la variación en su frecuencia de captura, las especies registradas se ubicaron en una gráfica que tuvo como ejes: (i) la proporción de periodos de muestreo en los que la especie fue registrada (p.ej., 1 de 4 = 0.25 a 4 de 4 = 1) y (ii) la proporción promedio de estaciones en las que la especie fue registrada durante el estudio. Todos los análisis se hicieron con el programa R (R Core Team, 2018).
Resultados
Con un esfuerzo de muestreo acumulado de 2,835 días-cámara, se obtuvieron 1,111 eventos de fototrampeo que incluyeron 19 especies de mamíferos terrestres pertenecientes a 13 familias y 7 órdenes. Las familias con mayor número de especies fueron Felidae (4 especies) seguida de Procyonidae y Didelphidae, con 3 especies cada una (Tabla 2). Por otra parte, se obtuvo el registro de una especie que no había sido previamente reportada (Conepatus semistriatus) para la reserva. De manera adicional, se obtuvieron registros de 2 especies de primates (Alouatta palliata y Ateles geoffroyi). Se registraron entre 11 y 18 especies por periodo de muestreo. La riqueza registrada en este estudio corresponde a 79% de las especies de mamíferos terrestres medianos y grandes que se esperaría encontrar con el fototrampeo en la REBISO de acuerdo con los datos reportados en 1996. El gremio alimentario con mayor número de especies fue el de los carnívoros con 6 especies, seguido de los herbívoros y los insectívoros, cada uno con 3 especies.
En general, se encontró un patrón similar de las estimaciones de riqueza de especies obtenidas en los distintos periodos tanto al hacer la comparación con el esfuerzo mínimo de muestreo, como al extrapolar al esfuerzo máximo (Fig. 2a). Solo la riqueza de especies registrada durante el primer periodo de muestreo (2013-2014) fue significativamente menor que el resto (no hay traslape en los intervalos de confianza de 95%). Las especies más frecuentes en los muestreos fueron consistentemente C. paca y D. mexicana, seguidas de Urocyon cinereoargenteus, solo en el primer periodo (Fig. 2b).
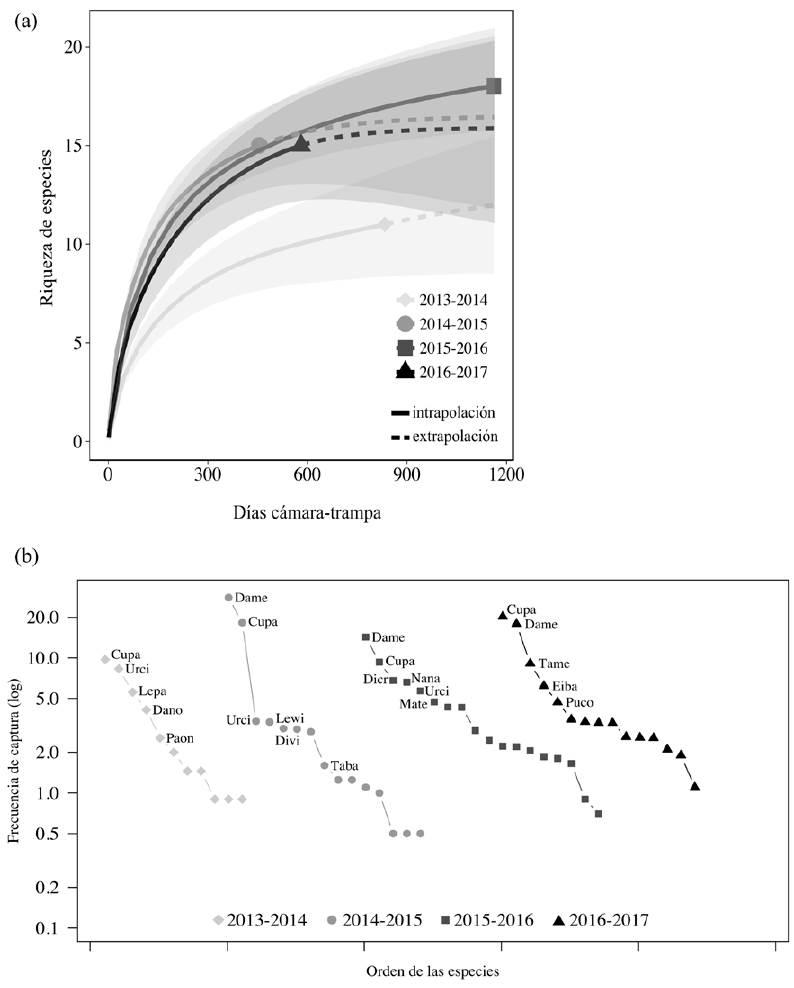
Figura 2 Comparación de la riqueza de especies de mamíferos (a) y su frecuencia de captura (b) en 4 periodos (2013-2016) de muestreo realizado en la Reserva de la Biosfera Selva El Ocote, Chiapas.
Se registraron 18 especies en la zona núcleo de la reserva, 16 en el área de influencia y 14 en la zona de amortiguamiento, en el periodo 2015-2016. La comparación de la riqueza de especies entre zonas, usando el esfuerzo de muestreo mínimo común en las curvas de rarefacción de especies, mostró que la riqueza que se alcanza en la zona núcleo y el área de influencia es muy similar (intervalos de confianza de 95% se traslapan completamente). En contraste, la riqueza de especies en la zona de amortiguamiento es claramente menor (no hay traslape en los intervalos de confianza de 95%). Este contraste disminuye cuando se usan los valores de riqueza correspondientes al esfuerzo de muestreo extrapolado. Aun así, la curva de rarefacción de especies para la zona de amortiguamiento se mantiene por debajo de las otras 2 curvas (Fig. 3a). Las especies más frecuentes en la zona núcleo fueron C. paca y D. crassus; en la zona de amortiguamiento D. mexicana y C. paca y en el área de influencia, D. mexicana, N. narica y C. paca (Fig. 3b).
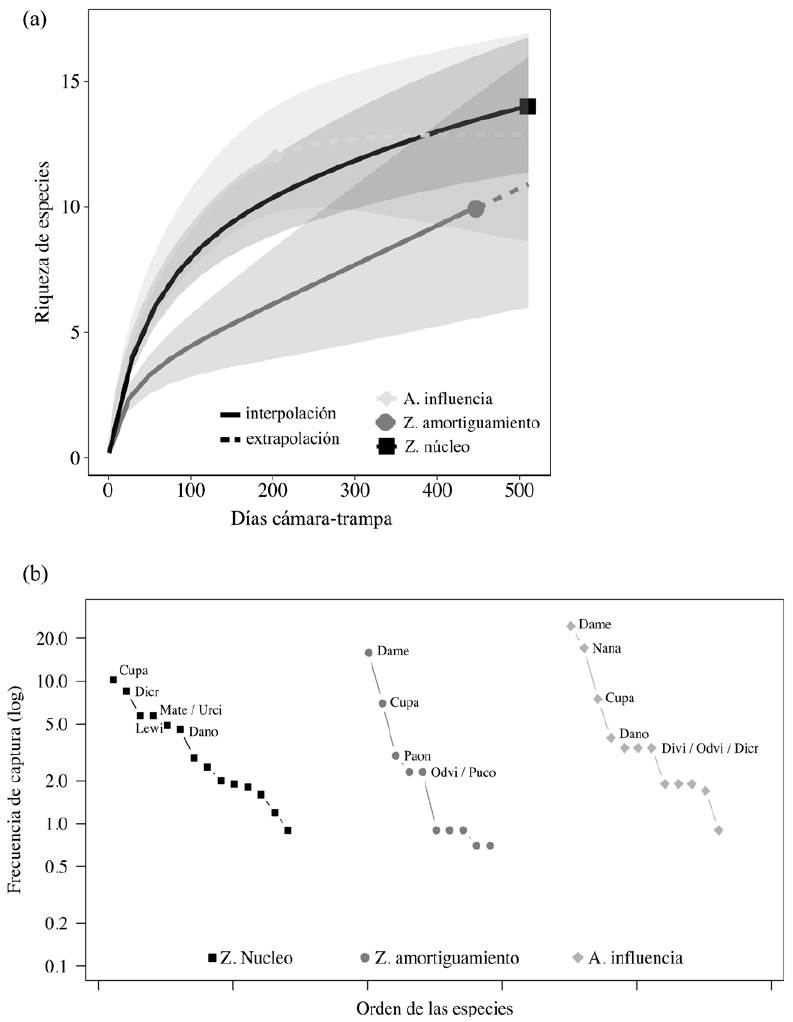
Figuras 3 Comparación de la riqueza de especies de mamíferos (a) y su frecuencia relativa (b) entre las zonas núcleo, de amortiguamiento y área de influencia de la Reserva de la Biosfera Selva El Ocote, Chiapas, para el periodo 2015-2016.
La composición de especies de mamíferos difirió entre zonas de la REBISO para el periodo 2015-2016 (Anosim R = 0.64, p < 0.05; Fig. 4). Solo 5 especies estuvieron presentes en las 3 zonas (C. paca, D. mexicana, N. narica, Odocoileus virginianus y Puma concolor). Las diferencias entre zonas parecen explicarse con base en la presencia única de ciertas especies (por ejemplo, C. semistriatus y Tamandua mexicana solo se registraron en la zona de amortiguamiento y Eira barbara y Galictis vittata solo estuvieron presentes en la zona núcleo), pero también con base en la magnitud de las diferencias en las frecuencias de captura entre zonas. Por ejemplo, la frecuencia de captura de los felinos Leopardus pardalis y L. wiedii fue entre 2 y 4 veces mayor en la zona núcleo que en la de amortiguamiento y área de influencia, respectivamente (Tabla 4). Por su parte, C. paca y P. concolor tuvieron frecuencias similares entre las 3 zonas. Algunas especies generalistas sólo se registraron o tuvieron una frecuencia más alta en el área de influencia. Por ejemplo, N. narica tuvo una frecuencia de 17.1 eventos en el área de influencia y de solo 0.9 y 1.8 en la zona de amortiguamiento y zona núcleo, respectivamente (Tabla 4).
Tabla 4 Frecuencia de captura de las especies de mamíferos terrestres registradas en el periodo 2015-2016 en las 3 zonas de la Reserva de la Biosfera Selva El Ocote.
| Especie / zona | Área de influencia | Zona de amortiguamiento | Zona núcleo |
|---|---|---|---|
| Conepatus semistriatus | -- | 0.7 | -- |
| Cuniculus paca | 7.5 | 7.0 | 10.2 |
| Dasyprocta mexicana | 24.4 | 15.8 | 1.9 |
| Dasypus novemcinctus | 4.0 | -- | 4.6 |
| Dicotyles crassus | 3.4 | -- | 8.5 |
| Didelphis marsupialis | 1.7 | -- | 2.0 |
| Didelphis virginiana | 3.4 | 0.7 | -- |
| Eira barbara | -- | -- | 1.7 |
| Galictis vittata | -- | -- | 2.9 |
| Leopardus pardalis | -- | 0.9 | 2.6 |
| Leopardus wiedii | 1.9 | -- | 4.9 |
| Mazama temama | 1.9 | -- | 5.7 |
| Nasua narica | 17.1 | 0.9 | 1.8 |
| Odocoileus virginianus | 3.4 | 2.3 | 0.9 |
| Panthera onca | 0.9 | 3.0 | -- |
| Puma concolor | 1.9 | 2.3 | 1.2 |
| Tamandua mexicana | -- | 0.9 | -- |
| Urocyon cinereoargenteus | -- | -- | 5.7 |
| Total de especies | 12 | 10 | 14 |
| Esfuerzo de muestreo (días cámara-trampa) | 200 | 443 | 502 |
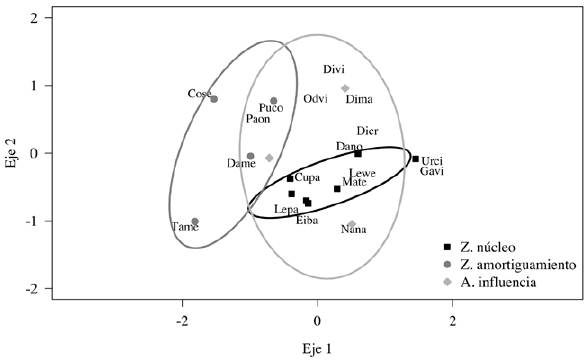
Figura 4 Ordenación de las estaciones de fototrampeo ubicadas en la zona núcleo, de amortiguamiento y área de influencia de la Reserva de la Biosfera Selva El Ocote, Chiapas, con base en las frecuencias de captura obtenidas en el periodo 2015-2016. Estrés = 0.11. Para la clave de las especies, ver tabla 2. Prueba Anosim, R = 0.64, p = 0.001.
Al combinar los datos sobre la proporción de las estaciones de fototrampeo en las que las especies fueron registradas a lo largo del estudio y la proporción de periodos en los que se observaron, fue posible distinguir algunos patrones. En primera instancia, destaca que C. paca fue la especie más común, ya que apareció en todos los muestreos y en más de 80% de las cámaras, en promedio (Fig. 5, cuadrante I). Por otra parte, destaca un contingente de especies encabezado por N. narica con una alta proporción de apariciones en los muestreos (> 60%) pero con registros en una proporción relativamente baja (< 50%) de las cámaras de cada muestreo. Dentro de este contingente, se incluyeron los depredadores tope P. onca y P. concolor, junto con mesodepredadores como: L. pardalis, L. wiedii, E. barbara y U. cinereoargenteus. También se incluyeron Didelphis virginiana y D. marsupialis, Dasypus novemcinctus, D. crassus así como D. mexicana y T. mexicana (cuadrante II). Finalmente, fue posible distinguir un grupo de especies raras que incluye al carnívoro G. vitatta, el insectívoro C. semistriatus y 3 herbívoros de gran tamaño: T. bairdii, Mazama temama y O. virginianus que fueron registradas en menos de 20% de los periodos y de las estaciones de fototrampeo (cuadrante III). No hubo especies que se ubicaran en el cuadrante IV, es decir, que fueran registradas en un solo periodo pero en una alta proporción de las estaciones de muestreo (i.e., > 0.5; Fig. 4).

Discusión
De las 24 especies de mamíferos medianos y grandes que se esperaba encontrar por fototrampeo en la REBISO, se lograron registrar 19 (79.2%). Entre las especies registradas se incluyó el zorrillo C. semistriatus, que no aparece en el listado de mamíferos de la REBISO realizado en 1996. Además, se obtuvo el primer registro videográfico del tapir centroamericano (https://youtu.be/PXNM7CYV0J0), una especie para la que no se contaba con evidencia formal de presencia en la REBISO desde 1977 (Álvarez-del Toro, 1977). Asimismo, es de destacar las altas frecuencias de captura de felinos como L. pardalis, P. onca, L. wiedii y P. concolor. Por el contrario, existen otras especies que no fueron registradas pero que están enlistadas en el plan de manejo de la REBISO, incluyendo Bassariscus sumichrasti, Conepatus leuconotus, Herpailurus yagouaroundi y Sylvilagus floridanus. Esto pudo deberse a que en el muestreo no se incluyeron hábitats como pastizales inducidos o vegetación altamente fragmentada en donde suelen registrarse algunas de estas especies. Es posible también, que estas especies tengan ya abundancias muy bajas en el área de estudio y que para detectarlas sea necesario un esfuerzo de muestreo mucho mayor o que por sus hábitos (p.ej., especies que también se mueven en la copa de los árboles) se requiera de métodos complementarios como cámaras trampas en el dosel, estaciones olfativas, búsqueda de huellas y rastros, y entrevistas a pobladores locales.
La riqueza de mamíferos terrestres medianos y grandes (> 500 g, n = 19) registrada en este estudio, es comparable a la encontrada en otros muestreos realizados en áreas protegidas en bosques tropicales del sur de México, en donde se han utilizado metodologías de muestreo similares a la empleada para este trabajo. Por ejemplo, en la zona núcleo I de la Reserva de la Biosfera el Triunfo, se han reportado 16 especies (Cornejo-Solchaga, 2015) y en los Petenes en Campeche, 13 especies (Hernández-Pérez et al., 2015). Asimismo, en la Reserva Montes Azules, en la región de la Lacandona en Chiapas, en los Chimalapas y en el istmo de Tehuantepec, Oaxaca, que son regiones identificadas como de alta diversidad mastofaunística, se ha registrado una riqueza de especies similar a la registrada en la REBISO (20, 19 y 20 especies, respectivamente; Azuara, 2005; Cortés-Maciel y Briones-Salas, 2014; Lira-Torres y Briones-Salas, 2012).
La riqueza de mamíferos medianos y grandes registradas en la REBISO es a su vez comparable, e incluso mayor, a la registrada en estudios realizados en otros bosques tropicales de Centro y Sudamérica (basados también en el uso de fototrampeo y con duraciones de por lo menos 1 año de muestreo). Por ejemplo, en la Reserva de la Biosfera Río Plátano en Honduras, Gunthier y Castañeda (2013) registraron 14 especies de mamíferos medianos y grandes; en los bosques de San Lorenzo y Pagaibamba en el norte de Perú, Jiménez et al. (2010) registraron 8 especies y De Souza-Martins et al. (2006) registraron 16 en el Bosque Nacional Caxiuana en Brasil. Solo las selvas de tierras bajas, al oeste del Ecuador, superan la riqueza registrada en la REBISO con 23 especies de mamíferos grandes y medianos (Blake y Loiselle, 2018). Por otra parte, los muestreos estandarizados de fototrampeo, que el grupo TEAM (Tropical Ecology Assessment and Monitoring Network) ha realizado en comunidades de mamíferos tropicales en Uganda, Tanzania, Indonesia, Laos, Surinam, Brasil y Costa Rica, han registrado una riqueza de entre 13 y 28 especies, con un promedio de 21 (Ahumada et al., 2011). Tomando estos datos como referencia, se confirma que la REBISO puede ser considerada como un sitio de alta importancia para la conservación de la diversidad de mamíferos medianos y grandes a nivel mundial.
En este estudio, se encontró que las especies más comunes fueron: C. paca, D. mexicana y N. narica. Este resultado concuerda con lo hallado en otras regiones tropicales de México y del neotrópico (Blake y Loiselle, 2018; Cornejo-Solchaga, 2015; Lira-Torres y BrionesSalas, 2012). C. paca y D. mexicana son especies que pueden ser abundantes a pesar de que son altamente cazadas (Gallina et al., 2012; Pérez-Irineo y SantosMoreno, 2013). Por ejemplo, se han calculado densidades entre 0.2 a 1 ind./ha y tasas de extracción de 488 ind./ año en el Parque Estatal de la Sierra en Tabasco (Pozo-Montuy et al., 2018). Por su parte, N. narica es una especie gregaria que forma grupos numerosos (Valenzuela, 2005), lo que en combinación con sus hábitos de alimentación omnívora, el tamaño grande de sus camadas y su alta capacidad reproductiva, le puede conferir la capacidad de tolerar el impacto antrópico (Yupanqui-Castañeda, 2005). Por el contrario, en la REBISO hay especies raras como T. bairdii, la cual se registró en un solo periodo y posee una frecuencia de captura muy baja. Esto contrasta con el hecho de que en otros bosques tropicales de la región (p. ej., la Reserva de la Biosfera Montes Azules, los Chimalapas y la Reserva de la Biosfera El Triunfo), el tapir es una especie con una frecuencia de captura relativamente alta (Azuara, 2005; Carbajal-Borges et al., 2014; Cornejo-Solchaga, 2015; Pérez-Irineo y Santos Moreno, 2016). Esta baja frecuencia podría deberse a los incendios de 1998, que afectaron más de 19,000 ha en la reserva y a los eventos de cacería que se registran en el corredor existente entre la región de los Chimalapas en Oaxaca y la REBISO.
Por otra parte, encontramos que la frecuencia de captura de especies sensibles a los cambios de uso de suelo como M. temama, L. pardalis y L. wiedii fue menor en la zona de amortiguamiento y área de influencia que en la zona núcleo, mientras que especies generalistas como D. virginiana y N. narica, fueron más frecuentes en las zonas de amortiguamiento y de influencia. De manera similar, Ahumada et al. (2011) observaron una disminución de la riqueza, diversidad y ocupación de especies de mamíferos en paisajes con mayor grado de fragmentación del bosque tropical.
Estudios multianuales, como el presentado aquí, son importantes para contar con una línea base a partir de la cual entender las tendencias de cambio de las comunidades de vertebrados en bosques tropicales (Pitman et al., 2011). Aunque hay ciertos avances en la evaluación de poblaciones de aves (Blake y Loiselle, 2015; Blake et al., 2017), los estudios multianuales con mamíferos son raros (Blake y Loiselle, 2018). Sin embargo, debido al constante impacto de las perturbaciones antrópicas (e.g., caza, actividades extractivas, desarrollo de carreteras y cambio en el uso del suelo), surge la necesidad de contar con estudios que evalúen indicadores de la biodiversidad en escalas temporales y espaciales amplias y usando métodos estandarizados (Brodie et al., 2015). Este trabajo es el primer estudio multianual para mamíferos medianos y grandes de la REBISO, aplicando la metodología de fototrampeo. En este sentido, es un esfuerzo que sienta las bases para contar con indicadores de la efectividad de la REBISO a largo plazo. El monitoreo a largo plazo podría ampliarse para evaluar otros elementos de la diversidad, por ejemplo, incluir la colocación de fototrampas en el dosel para el registro de mamíferos arborícolas (Méndez-Carvajal, 2014). Asimismo, se podría documentar la tasa de remoción de frutos por mamíferos frugívoros tanto en el piso de la selva como en el dosel (Camargo-Sanabria y Mendoza, 2016) para tener un indicador del rol funcional del gremio de los frugívoros y con esto, dar seguimiento a procesos ecológicos característicos de los bosques tropicales. Por ejemplo, algunos estudios realizados en la cercana región de la selva Lacandona en el estado de Chiapas, muestran que especies como T. bairdii y T. pecari (registradas con muy baja frecuencia en este estudio o totalmente ausentes) están entre los consumidores más importantes de los frutos que se acumulan en el piso del bosque en el caso de especies como la palma Attalea butyraceae y el árbol Pouteria sapota (Camargo-Sanabria y Mendoza, 2016; Martínez-Delgado, 2017). De esta manera, existe la posibilidad que aún en las porciones de bosque en la REBISO que presentan un dosel en buen estado de conservación, se presenten procesos de cambio en su potencial de regeneración de la vegetación, asociado a la extinción ecológica de algunas especies animales como el tapir. Esta es una línea de investigación que requiere atención y estudios específicos en la REBISO como se ha hecho en otros sitios (Camargo-Sanabria et al., 2015).
Las diferencias observadas en la composición de la comunidad de mamíferos en las distintas zonas dentro de la REBISO y en su área de influencia, pueden tener implicaciones profundas para la conservación de la biodiversidad. Es por ello, que es muy importante establecer y manejar el área de influencia con el fin de reducir o mitigar los impactos humanos hacia dentro de las reservas tomando en cuenta la heterogeneidad del paisaje que les rodea (Seiferling et al., 2011), las funciones y procesos ecológicos, así como la gestión del uso humano a nivel regional y su efecto de borde (Alexandre et al., 2010; Hansen y Defries, 2007). Nuestros resultados apuntan a que hay una relación entre el grado de actividad humana y la composición de especies de mamíferos. La presencia de las especies más frecuentes no mostró mayor variación entre periodos de muestreo, pero sí las especies exclusivas de las zonas núcleo. Esto indica que si las zonas núcleo son afectadas, se podría perder un componente muy importante de la comunidad de mamíferos.
Respecto a implicaciones de manejo, la presencia en la REBISO de especies de mamíferos que han sido extirpadas en otras áreas del país justifica la aplicación de estrategias de conservación como el de las Unidades de Manejo para la Conservación de la Vida Silvestre (UMAS). Esta estrategia es posible emplearla con especies relativamente abundantes como C. paca, D. mexicana y D. crassus con fines de aprovechamiento sustentable y para especies bandera como P. onca con fines de conservación, lo que ayudaría a proteger su hábitat y a impedir su desaparición y la de otras especies (Pozo-Montuy et al., 2017). Es urgente de igual manera, contar con un mayor apoyo por parte de las autoridades locales y federales para frenar los eventos de invasión por comunidades humanas a las zonas núcleo de la reserva y, en la medida de lo posible, revertirlas.
Finalmente, los análisis más recientes sobre escenarios de cambio climático para la región donde se encuentra la REBISO predicen que en el año 2039 será más cálida y seca de lo que es actualmente, con un aumento mínimo de 0.19 a 0.87 °C en la temperatura máxima promedio anual, de 0.41 a 1.03 °C en la temperatura media promedio anual y de 0.58 a 1.16 °C para la temperatura mínima promedio anual. De manera asociada, Manzanilla-Quiñones y Aguirre-Calderón (2017) estiman que en 31% de la superficie de la REBISO podría incrementarse la incidencia de incendios. Estas condiciones seguramente aumentarán la presión existente sobre las poblaciones de mamíferos silvestres, haciendo aún más importante contar con datos sólidos sobre los cambios en la riqueza y composición de sus comunidades.

