











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
México posee una increíble diversidad específica y ecosistémica, que lo sitúan entre los países megadiversos del planeta; por ejemplo, ocupa el segundo lugar en el mundo en cuanto al número de especies de reptiles y figura entre los primeros 5 lugares si se consideran las especies de plantas, anfibios y mamíferos (Espinosa-Organista et al., 2008). Por otra parte, los patrones biogeográficos que exhiben los taxones de nuestro país muestran una increíble complejidad. La parte septentrional del territorio mexicano se sitúa en la región Neártica, mientras que la parte más austral del país pertenece a la región Neotropical. Entre ambas, principalmente en las sierras Madre y la Faja Volcánica Transmexicana, la mezcla de taxones neárticos y neotropicales hace que esta área constituya una zona de transición, la llamada Zona de Transición Mexicana (Halffter, 1987; Halffter y Morrone, 2017). Esta encrucijada biótica del Nuevo Mundo hace que la diversidad de México sea tan interesante, no solo en cuanto a la enorme cantidad de especies endémicas, es decir restringidas al país en su distribución, sino en cuanto a sus variadas relaciones filogenéticas y a las hipótesis que se han planteado para explicar cómo se han ensamblado en el país taxones de orígenes tan diversos. Por otra parte, el país exhibe casi todos los tipos de vegetación del planeta, con desiertos, bosques tropicales y templados, pastizales y yermos de alta montaña (Challenger y Soberón, 2008). Es decir que México representa un increíble mosaico biótico.
La regionalización biogeográfica de México ha sido objeto de numerosos análisis (e.g., Arriaga et al., 1997; Cabrera y Willlink, 1973; Morrone, 2001, 2006; Rzedowski, 1978; Smith, 1941; Stuart, 1964; West, 1964). Hace unos 15 años (Morrone, 2005), se revisó la regionalización biogeográfica del país y se propuso una regionalización de consenso basada en las propuestas previas. El objetivo ahora fue revisar nuevamente la regionalización biogeográfica de México, analizando las dimensiones fisiográfica, morfotectónica, climatológica y vegetacional, así como analizar la evolución de las regiones Neártica y Neotropical, la Zona de Transición Mexicana y las 14 provincias reconocidas. Para cada una de estas unidades, se proporciona una lista de los nombres que han recibido, una caracterización general de la vegetación, algunos ejemplos de taxones endémicos de animales y plantas, y se discuten sus relaciones bióticas. Se incluye en esta contribución el reconocimiento de 42 distritos biogeográficos dentro de las provincias, así como comentarios acerca de algunos estudios biogeográficos referidos a la regionalización y evolución biótica de dichas provincias.
Dimensiones
El factor más importante que se ha correlacionado con la diversidad biológica de México es la heterogeneidad del medio físico, la cual es resultado de la historia geológica y el clima (Espinosa-Organista et al., 2008). A partir de las condiciones geológicas y climáticas se ha desarrollado en el país una biota particular (Morrone, 2005). Las dimensiones fisiográfica, morfotectónica, climatológica y vegetacional de México ayudarán a entender de manera más acabada la regionalización biogeográfica del país.
Dimensión fisiográfica
El territorio mexicano posee una superficie de 1,964,375 km2, situados a ambos lados del Trópico de Cáncer, con su extremo septentrional situado en el paralelo de 32°42’ N y el meridional en el paralelo de 14°30’ N. La mayor parte del territorio nacional se encuentra en América del Norte, mientras que su porción suroriental forma parte de América Central. La forma del país es aproximadamente triangular, con la base situada hacia el norte (donde alcanza los 2,000 km de costa a costa) y angostándose hacia el sur (donde alcanza poco más de 200 km de largo en el istmo de Tehuantepec). México posee 2 grandes penínsulas, la de Baja California en el noroeste y la de Yucatán en el sureste.
Desde la perspectiva fisiográfica, 65% de la superficie del país es montañosa, mientras que 35% restante no supera los 500 m de altitud. Existen 8 sistemas montañosos principales (Challenger, 1988; Ferrusquía-Villafranca, 1998; Fig. 1):
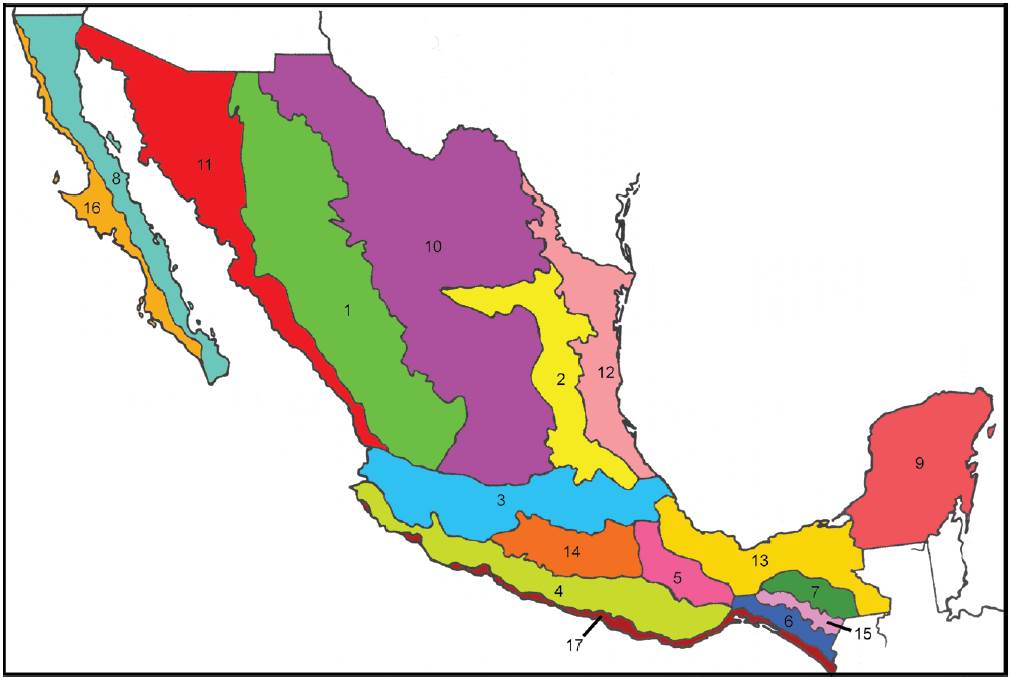
Figura 1 Provincias fisiográficas de México. 1, Sierra Madre Occidental; 2, Sierra Madre Oriental; 3, Faja Volcánica Transmexicana; 4, Sierra Madre del Sur; 5, Sierra Norte de Oaxaca; 6, Sierra Madre de Chiapas; 7, Meseta Central de Chiapas; 8, Cordillera de Baja California; 9, Plataforma de Yucatán; 10, Altiplano Mexicano; 11, Planicie Costera del Noroeste; 12, Planicie Costera del Noreste; 13, Planicie Costera del Golfo de México; 14, Cuenca del Balsas; 15, Depresión Central de Chiapas; 16, Planicie Costera de Baja California; 17, Planicie Costera del Sureste.
Cordillera de Baja California. Serranía de 1,430 km de longitud que recorre la península de Baja California. Comprende las sierras La Rumorosa, Juárez, San Pedro Mártir, La Libertad, La Giganta y San Lázaro. Posee su máximo desarrollo en la parte norte, donde la sierra de San Pedro Mártir alcanza los 2,400 m de altura. Este sistema se continúa hacia el norte en los EUA con la Sierra Nevada.
Sierra Madre Occidental. Representa el mayor sistema montañoso del país, extendiéndose paralelamente a la costa del océano Pacífico desde la frontera con los EUA hasta los estados de Nayarit y Jalisco, donde se conecta con la Faja Volcánica Transmexicana. Representa la prolongación meridional de las Montañas Rocosas de los EUA y separa la Planicie Costera Noroccidental del Altiplano Mexicano. En el este, los ríos que fluyen hacia el Pacífico han excavado barrancas profundas. Posee una extensión de 289,000 km y su altitud mayor es de poco más de 3,000 m.
Sierra Madre Oriental. Se inicia en el estado de Nuevo León y corre en dirección sur-sureste, conectándose con la Faja Volcánica Transmexicana en el centro de Puebla y Veracruz. Separa el Altiplano Mexicano de la Planicie Costera del Noreste. Posee unos 1,350 km de largo y su elevación en general es menor que la de la Sierra Madre Occidental. Posee algunas elevaciones importantes, como el cerro Potosí (3,650 m) en Nuevo León y el cerro de San Antonio Peña Nevada (3,450 m) en el límite entre Nuevo León y Tamaulipas.
Faja Volcánica Transmexicana. Sistema que cruza el país en dirección este-oeste, aproximadamente a lo largo de los paralelos de 19 y 20° N. Posee unos 1,000 km de longitud y unos 50-150 km de ancho. Separa el Altiplano Mexicano de la Cuenca del Balsas. Incluye las elevaciones mayores del territorio mexicano, constituidas por los volcanes Pico de Orizaba (5,650 m), Popocatépetl (5,450 m), Ixtaccíhuatl (5,280 m), Nevado de Toluca (4,560 m), La Malinche (4,460 m), Nevado de Colima (4,340 m), Tancítaro (4,160 m), Tláloc (4,150 m) y Cofre de Perote (4,090 m), aunque la altura media es de 2,000 m.
Sierra Madre del Sur. Corre de noroeste a sureste desde el estado de Jalisco hasta el istmo de Tehuantepec, muy cerca de la costa del océano Pacífico. Posee unos 1,100 km de longitud y su ancho promedio es de 120 km. Generalmente posee altitudes mayores a los 1,000 m y se encuentra interrumpida por los valles de numerosos ríos, como Armería, Balsas, Papagayo y Verde. Las elevaciones menores poseen unos 300-500 m de altura y las máximas llegan a los 3,400 m, en el cerro Teotepec, Guerrero.
Sierra Norte de Oaxaca. Ocupa la porción septentrional del estado de Oaxaca y áreas adyacentes de Puebla y Veracruz. En general, predominan las altitudes superiores a los 1,000 m y las elevaciones más altas se alcanzan en la Sierra de Juárez (3,400 m). Los ríos que la cruzan desembocan en las cuencas de los ríos Papaloapan y Coatzacoalcos.
Sierra Madre de Chiapas. Es una prolongación septentrional del Núcleo Centroamericano. Corre a lo largo del litoral pacífico de Chiapas y penetra hasta el istmo de Tehuantepec. Alcanza su mayor altitud en Tacaná (4,026 m), en la frontera con Guatemala.
Macizo Central de Chiapas. Es otra de las prolongaciones septentrionales del Núcleo Centroamericano. También corre paralelo a la costa del océano Pacífico, pero más alejado de ésta. Alcanza su mayor altitud en el área de San Cristóbal (2,860 m).
Además de estos sistemas montañosos, México posee 9 llanuras y depresiones principales (Fig. 1):
Planicie Costera de Baja California. Situada en la porción occidental de la península de Baja California.
Planicie Costera del Noroeste. Situada en el extremo noroccidental del país, extendiéndose desde la desembocadura del río Colorado hasta el centro de Nayarit, siendo más ancha a lo largo de la costa de Sonora.
Altiplano Mexicano. Altiplano delimitado principalmente por las sierras Madre Occidental y Oriental, y la Faja Volcánica Transmexicana. Abarca los estados de Chihuahua, Coahuila, San Luis, Potosí, Zacatecas, Durango, Guanajuato, Aguascalientes y Jalisco. Es una extensa meseta surcada por serranías más o menos aisladas. La mayor parte de las tierras planas se ubican a 2,000 m hacia el sur, mientras que en el norte predominan las altitudes de 1,000-1,500 m. El Altiplano Mexicano continúa hacia el norte en las grandes llanuras del oeste de los EUA.
Planicie Costera del Noreste. Franja de tierras bajas situadas hacia el sur del río Bravo, que alcanzan el centro del estado de Veracruz. Hacia el norte se continúa con la gran llanura del sureste de los EUA. Abarca casi todo el estado de Tamaulipas y partes de Nuevo León, San Luis Potosí y Veracruz.
Planicie Costera del Sureste. Tierras bajas situadas en el sureste, desde el centro de Veracruz hasta Tabasco.
Cuenca del Balsas. Tierras bajas situadas entre la Faja Volcánica Transmexicana y la Sierra Madre del Sur, en los estados de Michoacán, Guerrero, Morelos y Puebla. Las partes más bajas se sitúan a los 300 m de altitud.
Planicie Costera del Sureste del Golfo de México. Se extiende desde el centro de Veracruz (Sierra de Naolinco) hasta Tabasco.
Plataforma de Yucatán. Planicie de la Península de Yucatán, que no sobrepasa los 200 m de altitud.
Depresión Central de Chiapas. Corredor de tierras bajas situadas entre la Meseta Central y la Sierra Madre de Chiapas. Posee 250 km de largo y un ancho promedio de 75 km. Sus altitudes menores alcanzan los 400 m.
Dimensión morfotectónica
Desde la perspectiva geológica histórica, resulta interesante analizar la evolución tectónica del territorio que hoy en día constituye México. En el país convergen 5 placas tectónicas (Ortega-Gutiérrez et al., 2000; Fig. 2): Norteamericana, Pacífico, Rivera, Cocos y Caribe. A partir de la interacción de estas placas, se han originado las sierras Madre Oriental y del Sur por plegamiento y la Sierra Madre Occidental y la Faja Volcánica Transmexicana por vulcanismo. La caracterización e historia geológica de las provincias morfotectónicas de México han sido analizadas por Ferrusquía-Villafranca (1998) y Ortega-Gutiérrez et al. (2000), quienes proporcionan una síntesis de la evolución tectónica de México, con especial referencia a los eventos ocurridos en el Fanerozoico.
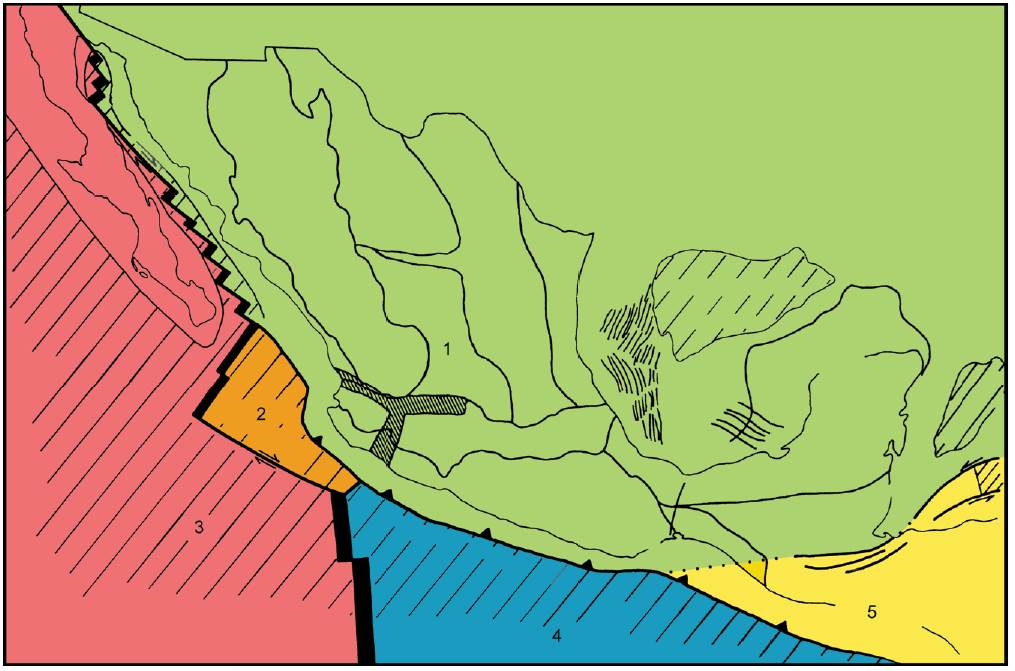
Figura 2 Placas tectónicas del territorio mexicano. 1, Norteamericana; 2, Rivera; 3, Pacífica; 4, Cocos; 5, Caribeña.
La secuencia temporal de mapas paleogeográficos de América del Norte (Fig. 3) muestra la evolución del territorio en diferentes épocas: (a) Cretácico Temprano (hace 130 ma); (b) Cretácico Temprano (hace 110 ma); (c) Cretácico Tardío (hace 87 ma); (d) Paleoceno (hace 60 ma); (e) Eoceno (hace 50 ma); (f) Oligoceno (hace 35 ma); (g) Mioceno (hace 20 ma); (h) Plioceno (hace 5 ma); (i) Reciente. Estas configuraciones diferentes son importantes para comprender las distintas conexiones bióticas de México en el pasado, en relación con América del Norte, las Antillas y América del Sur (Graham, 2018).

Figura 3 Secuencia temporal de mapas paleogeográficos de América del Norte. (a) Cretácico Temprano (hace 130 ma); (b) Cretácico Temprano (hace 110 ma); (c) Cretácico Tardío (hace 87 ma); (d) Paleoceno (hace 60 ma); (e) Eoceno (hace 50 ma); (f) Oligoceno (hace 35 ma); (g) Mioceno (hace 20 ma); (h) Plioceno (hace 5 ma); (i) Reciente.
Dimensión climatológica
Debido a su ubicación geográfica y a su diversidad fisiográfica, México posee una gran diversidad climática. A ella se suma la influencia del océano Pacífico y el Golfo de México y la alineación de las sierras Madre Occidental y Oriental, que determinan la aridez del Altiplano Mexicano. La influencia de los vientos dominantes (alisios) actúa con una asimetría clara entre la vertiente atlántica más húmeda y la pacífica más seca.
De acuerdo con el sistema de Köppen (1948), México posee los tipos climáticos A, B, C y E. Los climas de tipo A (calientes y húmedos) predominan en las tierras bajas del Pacífico, en la Península de Yucatán, Veracruz, Tabasco y partes de Chiapas. Los climas de tipo B (secos) cubren la mayor superficie del país, en particular en el Altiplano Mexicano, las Planicies Costeras Noroccidental y Nororiental, y la Península de Baja California. Los climas de tipo C (templados y húmedos) son típicos de las zonas de montaña, en las sierras Madre y la Faja Volcánica Transmexicana. Finalmente, los climas de tipo E (fríos o polares) solo se presentan en las zonas más altas de algunas montañas, siendo de distribución muy restringida.
Las temperaturas medias anuales varían entre 10 y 28 °C. Las temperaturas se correlacionan con la altitud, siendo más altas en las tierras bajas de las costas y más bajas en las cimas de las montañas. Las temperaturas más elevadas del país se registran en la Cuenca del Balsas (2830 °C), mientras que las más bajas se registran en el Pico de Orizaba (-6 °C). Las temperaturas mínimas extremas (menos de -15 °C) se registran en el norte de la Sierra Madre Occidental y áreas adyacentes del Altiplano. Las temperaturas máximas extremas (más de 45 °C) ocurren en el norte del país, a ambos lados del golfo de California, en la Planicie Costera Nororiental y el norte del Altiplano Mexicano. La oscilación anual de las temperaturas, es decir la diferencia que existe entre el mes más frío y el más cálido del año, es casi nula en las costas de Chiapas y llega hasta los 25 °C en el norte del país, donde el clima es más extremoso.
La precipitación varía entre un promedio de 50 mm anuales hasta más de 5,500 mm. Las áreas más húmedas se encuentran en el sureste, en Veracruz, Tabasco, la base de la Península de Yucatán, el norte de Chiapas y partes de Oaxaca, Puebla e Hidalgo. Las áreas más secas corresponden a la Planicie Costera de Sonora, la mayor parte de la Península de Baja California y el Altiplano Mexicano.
Dimensión vegetacional
A partir de la interacción entre flora, suelo y clima, resultan patrones de comunidades vegetales conocidos como biomas, dentro de los cuales, a su vez, es posible reconocer distintos tipos de vegetación. En México, varias contribuciones (Challenger y Soberón, 2008; González-Medrano, 2008; Miranda y Hernández, 1963; Pennington y Sarukhán, 1998; Rzedowski, 1978) han caracterizado y clasificado los tipos de vegetación de México. Una síntesis relativamente reciente (Villaseñor y Ortiz, 2013) reconoce para el país 5 biomas principales (Fig. 4), los cuales a su vez agrupan 21 tipos de vegetación:

Figura 4 Biomas principales de México reconocidos por Villaseñor y Ortiz (2013). 1, Matorral xerófilo; 2, bosque templado; 3, bosque húmedo de montaña; 4, bosque tropical estacionalmente seco; 5, Bosque tropical húmedo.
Matorral xerófilo. Reúne la vegetación arbustiva de regiones de clima seco, como es la porción neártica en sentido estricto de México. Comprende plantas espaciadas, con amplias superficies del suelo descubiertas. Este bioma habría ocupado en el pasado un tercio de la superficie del país (Sánchez-Colón et al., 2009). Altitudinalmente se sitúa entre los 3,000 m y el nivel del mar. Incluye la vegetación de chaparral, matorral submontano, matorral de desiertos fríos y matorral de desiertos cálidos. En esos tipos de vegetación abundan las plantas suculentas y semisuculentas, con predominio de familias asociadas a zonas áridas, como Cactaceae, Asparagaceae (agaves) y Crassulaceae (Challenger y Soberón, 2008).
Bosque húmedo de montaña. Comunidad vegetal con predominio de árboles, localizada en ambas vertientes del país, en zonas donde los vientos húmedos originados en el mar producen una alta concentración de humedad. Los árboles poseen 15-35 m de altura, muchos son caducifolios, pero otros conservan sus hojas todo el año. Altitudinalmente se sitúan entre los 1,000 y los 3,000 m de altitud. Incluye diferentes bosques, como los de neblina, mesófilo de montaña, de Liquidambar y templado lluvioso, entre otros. El bosque mesófilo de montaña incluye diferentes comunidades de ambientes templados y muy húmedos, situados entre los 800 y los 2,200 m. La mayor parte de sus árboles son perennifolios, aunque en terrenos escarpados de la Sierra Madre Oriental, la Sierra de Juárez y la Sierra Norte de Chiapas hay comunidades con especies que pierden sus hojas en invierno, como Carpinus tropicalis, Ulmus mexicana, Liquidambar styraciflua y Fagus mexicana (Challenger y Soberón, 2008). En el pasado, los bosques mesófilos de montaña ocuparon una mayor extensión, pero en la actualidad abarcan 1% de la superficie del país (Challenger, 2007).
Bosque templado. Comunidad dominada por árboles que se encuentra en zonas templadas y semihúmedas de montaña. Pueden ser perennifolios o totalmente caducifolios. Su altura varía ente 2 y 30 m. Altitudinalmente se sitúan entre el nivel del mar y los 3,500 m. Junto con el bosque húmedo de montaña caracterizan la Zona de Transición Mexicana. Incluye los bosques de Abies, encino, pino y pino-encino. Los bosques de pino son los de mayor extensión y se encuentran en todas las áreas montanas del país, formando bosques de altura, siendo su densidad variable y su diversidad escasa, pues usualmente predomina una especie (Challenger y Soberón, 2008). Los bosques de pino-encino poseen una diversidad mayor, albergando un gran número de especies animales y vegetales (Challenger, 2007).
Bosque tropical estacionalmente seco. Comunidad vegetal dominada por árboles, en regiones cálidas y con una larga temporada de sequía. Posee árboles relativamente bajos (5-15 m), la mayor parte de los cuales pierde sus hojas en la temporada seca. Es más abundante en la vertiente pacífica del país. Altitudinalmente se encuentra desde el nivel del mar hasta los 1,900 m. Estos bosques, en general, se consideran menos complejos estructuralmente y con menor diversidad que los bosques húmedos (Trejo, 2010). Incluye el bosque tropical caducifolio y el bosque espinoso.
Bosque tropical húmedo. Comunidad dominada por árboles, en zonas con precipitación abundante y sin heladas. Los árboles son comúnmente perennifolios y miden entre 35-40 m de altura. Altitudinalmente se sitúa entre el nivel del mar y los 1,000 m. Junto con el bosque tropical estacionalmente seco, caracterizan la porción neotropical en sentido estricto de México. Incluye los bosques tropicales lluvioso, perennifolio y siempreverde. De estos tipos de vegetación, el bosque tropical perennifolio es el más exuberante, con árboles de más de 30 m de altura y con cierta frecuencia algunas especies, como Guatteria anomala, Swietenia macrophylla y Terminalia amazonia, alcanzan los 75 m (Pennington y Sarukhán, 1998). Este bosque es más abundante en las planicies del Golfo de México, el sur y el este de la Península de Yucatán y el este de Chiapas (Challenger y Soberón, 2008). El bosque tropical húmedo se desarrolla en el trópico húmedo y subhúmedo en la costa del Pacífico, cañones de la vertiente del Pacífico de la Sierra Madre Occidental, planicies de la Península de Yucatán y sur de Veracruz y Tabasco, en áreas en que el suelo se anega completamente en la temporada de lluvias (Challenger y Soberón, 2008).
De acuerdo con Villaseñor y Ortiz (2013), el bosque templado es el bioma más rico en especies de plantas y el bosque tropical húmedo el de menor riqueza. En cuanto a sus especies endémicas, el bosque templado es el que posee la riqueza mayor, si bien todos los demás biomas poseen una endemicidad pronunciada. Cabe señalar la relevancia del bosque húmedo de montaña, pues si bien es tercero en cuanto a su riqueza específica (después del bosque templado y el matorral xerófilo), ocupa una porción significativamente menor que los demás y es el bioma más amenazado de México.
Además de los biomas y tipos de vegetación antes descritos, vale la pena mencionar la existencia de otros 2 tipos de vegetación: pastizales y humedales. Los pastizales son comunidades donde abundan las gramíneas e incluyen los zacatonales, páramos de altura y sabanas (Challenger y Soberón, 2008). Los humedales son comunidades muy variadas, que crecen en áreas saturadas o cubiertas de agua, como la vegetación marina litoral, manglares, popales, tulares, carrizales, vegetación sumergida y bosques de galería. Destacan los manglares, particularmente adaptados a la inundación y mareas, con mangle rojo (Rhizophora mangle), mangle blanco (Laguncularia racemosa), mangle negro (Avicennia germinans) y botoncillo (Conocarpus erecta), por lo general, comportándose alguna de estas especies como dominante.
Regionalización biogeográfica de México
Durante el siglo XX se publicaron varias regionalizaciones biogeográficas de México (Espinosa-Organista et al., 2008; Luna-Vega, 2008; Morrone, 2005, 2014a). Estas regionalizaciones se basan en distintos taxones de animales, plantas o una combinación de ellos. Aquí comento brevemente algunas de ellas.
Smith (1941)
Uno de los primeros esquemas zoogeográficos de México fue propuesto por Smith (1941). Este autor consideró que las especies de lagartijas del género Sceloporus constituían un grupo ideal para analizar la biogeografía de México debido a su gran diversidad, su intervalo altitudinal (desde el nivel del mar hasta 3,800 m), su presencia en prácticamente todos los hábitats terrestres y lo poco que su distribución geográfica había sido afectada por factores ambientales y los seres humanos. Reconoció 23 provincias biogeográficas (Fig. 5), las que clasificó en 2 regiones y 3 subregiones.
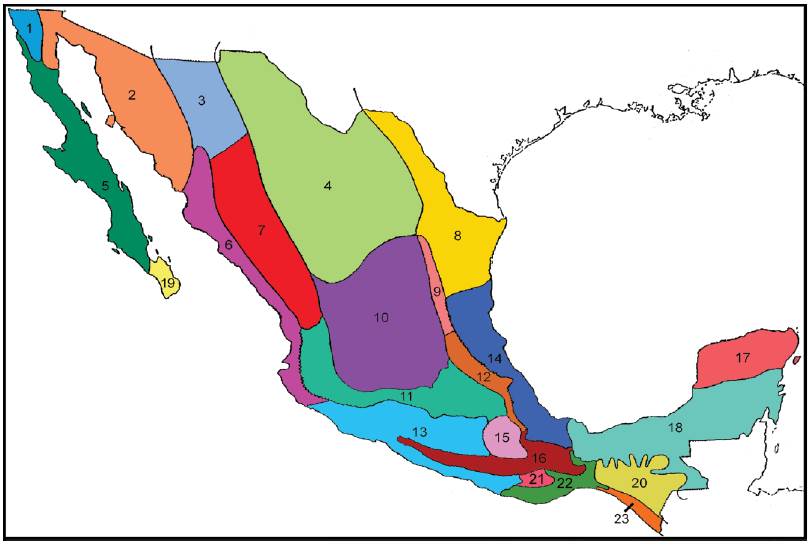
Figura 5 Provincias reconocidas en el esquema biogeográfico de Smith (1941). 1, Sandieguina; 2, Arizoniana; 3, Apache 4, Chihuahuense; 5, Baja California; 6, Sinaloense; 7, Durangueña; 8, Tamaulipeca; 9, Austro-Oriental; 10, Guerrerense; 11, AustroCentral; 12, Hidalguense; 13, Balsas Inferior; 14, Veracruzana; 15, Balsas Superior; 16, Austro-Occidental; 17, Yucateca; 18, del Petén; 19, del Cabo de Baja California; 20, de la Altiplanicie Chiapaneca; 21, de la Altiplanicie Oaxaqueña; 22, Tehuana; 23, Tapachulteca.
La región Neártica comprende la subregión de las Montañas Rocosas (con 15 provincias) y la subregión Californiana (con la provincia Sandieguina). Las provincias neárticas son las siguientes:
Provincia de la Altiplanicie Oaxaqueña. Porción elevada y pequeña, constituida mayoritariamente por planicies con pastizales, que se sitúa en la parte superior del río Balsas y se extiende hacia el sur hasta el río Tehuantepec.
Provincia Guerrerense. Porción sur de la altiplanicie al sur del río Balsas, en los estados de Guerrero y Oaxaca, que se extiende hacia el norte por el lado oriental hasta El Mirador (Veracruz) y hacia el sur hasta el istmo de Tehuantepec. Dentro de ésta se reconocen 2 secciones: Oriental y Occidental.
Provincia del Balsas Superior. Área desértica o semiárida, que limita con el extremo austral de la Altiplanicie Mexicana, ocupando la parte superior de la Cuenca del río Balsas. De acuerdo con el autor, esta provincia conecta las regiones Neártica y Neotropical.
Provincia Austro-Central. Área amplia y uniforme, situada en el centro de la Altiplanicie Mexicana, rodeada por el este, sur y oeste por cadenas montañosas. En el noroeste, esta provincia se fusiona con la provincia Chihuahuense.
Provincia Austro-Occidental. Área montañosa situada en el borde suroeste del Altiplano Mexicano, que se extiende desde el norte de Nayarit hasta Jalisco, el sur de la Ciudad de México y el norte de Puebla. Dentro de esta provincia se reconocen 3 secciones: Occidental, Central y Oriental.
Provincia Austro-Oriental. Norte de la región montañosa que forma el ángulo oriental del Altiplano Mexicano.
Provincia Hidalguense. Sección sur de la región montañosa del borde oriental del Altiplano Mexicano.
Provincia Chihuahuense. Área central al norte de la provincia Austro-Central, al este de la Sierra Madre Occidental y al oeste de la provincia Tamaulipeca. Dentro de esta provincia se reconocen 3 secciones: Occidental, Central y Oriental.
Provincia Sinaloense. Región costera desde el sur de Sonora hasta el sur de Mazatlán.
Provincia Tamaulipeca. Región costera ancha que llega hasta el Trópico de Cáncer.
Provincia Arizoniana. Parte occidental del estado de Sonora y el noroeste de Baja California.
Provincia Apache. Ángulo occidental montañoso de la meseta que se extiende desde el centro de Chihuahua hasta el sureste de Arizona.
Provincia Durangueña. Montañas de la Sierra Madre Occidental desde el centro de Chihuahua hasta el norte de Nayarit.
Provincia de Baja California. La mayor parte de la península de Baja California, con excepción del extremo sur, que corresponde a la provincia del Cabo, y el extremo norte, que corresponde a las provincias Sandieguina y Arizoniana.
Provincia del Cabo de Baja California. Extremo sur de la península de Baja California, al sur y este de la isla de Espíritu Santo.
Provincia Sandieguina. Centro y norte de la península de Baja California al norte del paralelo de 30°.
Para la región Neotropical, Smith (1941) reconoció la subregión Mexicana, con 7 provincias biogeográficas:
Provincia de la Altiplanicie Chiapaneca. Región elevada que se extiende al oeste hasta el istmo de Tehuantepec y al este hacia el interior de Guatemala.
Provincia Tapachulteca. Región costera al sur de la provincia anterior, que se extiende desde la frontera occidental de Chiapas hasta Guatemala. Dentro de esta provincia se reconocen 2 secciones: Norte y Sur.
Provincia del Petén. Área septentrional de la Altiplanicie Chiapaneca, incluyendo la vertiente del Golfo de México en el istmo de Tehuantepec. Hacia el norte se extiende hasta el paralelo de 20° de latitud norte y al este hasta Guatemala y Belice.
Provincia Yucateca. Península de Yucatán al norte del paralelo de 20° de latitud norte.
Provincia Veracruzana. Área costera al norte del río Papaloapan hasta el Trópico de Cáncer.
Provincia Tehuana. Región semiárida del estado de Oaxaca, que se extiende desde la Altiplanicie Oaxaqueña y la provincia Guerrerense al sur hacia la costa, al este hasta los límites de Chiapas y al oeste aproximadamente hasta el río Verde.
Provincia del Balsas Inferior. Área extendida a lo largo de las costas del Pacífico desde el río Verde en Oaxaca hasta el sur de Mazatlán en Sinaloa, extendiéndose por el río Balsas hasta el norte de Oaxaca y el suroeste de Puebla.
El esquema zoogeográfico de Smith (1941) es bastante detallado, aunque dado que se restringe a México, resulta difícil evaluar cómo el autor relaciona las provincias mexicanas con otras situadas fuera del territorio nacional. Sin embargo, la asignación de 16 provincias a la región Neártica y 7 a la Neotropical, plantea la existencia del límite entre ambas regiones dentro del territorio mexicano, tal como ha sido reconocido por otros autores.
Stuart (1964)
Este autor realizó un análisis detallado de la fauna de vertebrados de México y América Central. En el mapa que resume su contribución (Fig. 6), Stuart (1964) reconoció 16 provincias para México: California, Sanlucasense, Sonoriana, Sierra Madre Occidental, Chihuahua-Zacatecas, Tamaulipense, Sinaloense, Sierra Madre Oriental, Volcánica Transversa, Nayarit-Guerrero, Veracruzana, Sierra Madre del Sur, Tehuantepec, Tierras Altas de Chiapas-Guatemala, Yucatán y Petén. El esquema de West (1964), publicado en la misma obra que el anterior, propuso 13 provincias que guardan cierta semejanza con las anteriores.
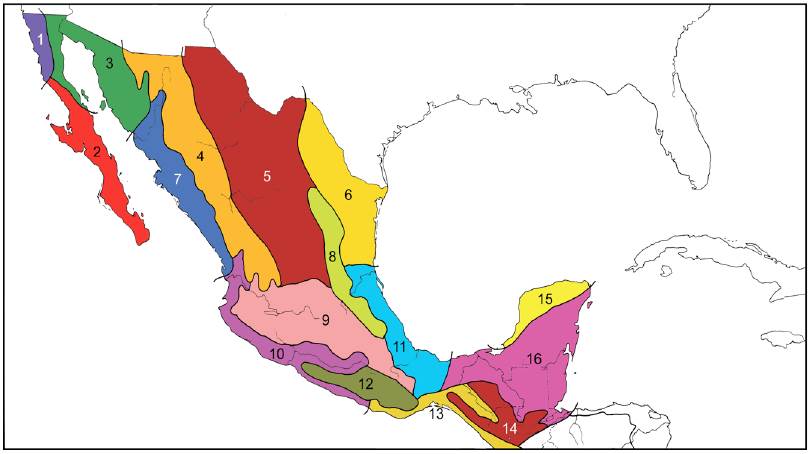
Figura 6 Provincias reconocidas en el esquema biogeográfico de Stuart (1964). 1, California; 2, Sanlucasense; 3, Sonoriana; 4, Sierra Madre Occidental; 5, Chihuahua-Zacatecas; 6, Tamaulipense; 7, Sinaloense; 8, Sierra Madre Oriental; 9, Volcánica Transversa; 10, Nayarit-Guerrero; 11, Veracruzana; 12, Sierra Madre del Sur; 13, Tehuantepec; 14, Tierras Altas de Chiapas-Guatemala; 15, Yucatán; 16, Petén.
Cabrera y Willink (1973)
Estos autores presentaron un esquema biogeográfico de regiones, dominios, provincias y, en algunos casos, distritos de América Latina y el Caribe, detallando los taxones animales y vegetales de cada área. Para México, reconocieron 4 provincias (Fig. 7), las cuales clasificaron en 2 regiones y 3 dominios.

Figura 7 Provincias reconocidas en el esquema biogeográfico de Cabrera y Willink (1973). 1, Bosque Montano; 2, Mesoamericana de Montaña; 3, Xerófila Mexicana; 4, Caribe; 5, Guajira; 6, Pacífica.
La región Holártica comprende el dominio Norteamericano Pacífico, con una sola provincia:
Provincia del Bosque Montano. Abarca desde el suroeste de los EUA, penetrando en México en el norte de la península de Baja California, sobre las montañas de San Pedro Mártir.
La región Neotropical dentro del territorio mexicano comprende el dominio Caribe (con las provincias Mesoamericana de Montaña y Xerófila Mexicana) y el dominio Amazónico (con la provincia Pacífica). Las provincias mexicanas incluidas en la región Neotropical son las siguientes:
Provincia Mesoamericana de Montaña. Ocupa las “tierras frías” de México y América Central, sobre las sierras Madre Occidental y Oriental y las montañas del sur de México, Guatemala y Panamá. Su altitud varía de acuerdo con la orientación y la latitud, pero usualmente se halla entre 1,000 y poco más de 4,000 m.
Provincia Xerófila Mexicana. Ocupa la mayor parte del territorio mexicano, abarcando áreas xéricas con lluvias inferiores a 750 mm anuales. Dentro de esta provincia, Cabrera y Willink (1973) reconocieron 3 distritos: el Sonoriano, que abarca el estado de Sonora, el norte de Sinaloa y la península de Baja California; el Chihuahuense, que ocupa el Altiplano Mexicano entre 1,000 y 2,000 m de altitud; y el Árido del Sur de México, que se encuentra en la porción austral de la provincia.
Provincia Pacífica. Abarca desde las laderas occidentales bajas de los Andes ecuatorianos y colombianos hasta las regiones bajas y las costas de América Central y el sur de México.
Cuando se lo compara con otros esquemas biogeográficos, el tratamiento de México parece demasiado simplificado, pues Cabrera y Willink (1973) solo reconocen 4 provincias. Sin embargo, estas provincias se asignan a 2 regiones y 3 dominios diferentes, destacando así la naturaleza compleja de la regionalización biogeográfica del país.
Rzedowski (1978)
El libro “Vegetación de México” (Rzedowski, 1978) constituye un hito en la fitogeografía de México (para un análisis de las contribuciones de Jerzy Rzedowski, ver Luna-Vega, 2008). Rzedowski (1978) discute las bases fisiográficas y climáticas, la influencia del hombre, las relaciones geográficas y los posibles orígenes de la flora mexicana, las formas y espectros biológicos, y los tipos de vegetación de nuestro país. En el capítulo dedicado a las provincias florísticas de México, presenta un esquema de 17 provincias (Fig. 8), las cuales clasifica en 2 reinos y 4 regiones.

Figura 8 Provincias reconocidas en el esquema biogeográfico de Rzedowski (1978). 1, California; 2, Isla Guadalupe; 3, Sierra Madre Occidental; 4, Sierra Madre Oriental; 5, Serranías Meridionales; 6, Serranías Transístmicas; 7, Baja California; 8, Planicie Costera del Noroeste; 9, Altiplanicie; 10, Planicie Costera del Noreste; 11, Valle de Tehuacán-Cuicatlán; 12, Costa Pacífica; 13, Islas Revillagigedo; 14, Depresión del Balsas; 15, Soconusco; 16, Costa del Golfo de México; 17, Península de Yucatán.
Dentro del reino Holártico, existen 2 regiones: Pacífica Norteamericana y Mesoamericana de Montaña. La región Pacífica Norteamericana incluye 2 provincias fitogeográficas:
Provincia de California. Abarca la mayor parte del estado de California, extendiéndose a algunas partes vecinas, como el sector septentrional de la península de Baja California, que incluye las sierras de Juárez y San Pedro Mártir y la planicie costera adyacente del lado del océano Pacífico.
Provincia de la Isla Guadalupe. Isla situada a 200 km de la costa de Baja California.
A su vez, la región Mesoamericana de Montaña, que Rzedowski (1978) consideró intermedia entre los 2 reinos, posee 4 provincias fitogeográficas:
Provincia de la Sierra Madre Occidental. Se extiende desde Sonora y Chihuahua en el norte hasta Nayarit, Zacatecas y norte de Jalisco en el sur.
Provincia de la Sierra Madre Oriental. Incluye partes de Coahuila, Nuevo León, Tamaulipas, San Luis Potosí, Querétaro, Hidalgo, Veracruz y Puebla. Su límite austral es difícil de delimitar, pues se une sin solución de continuidad con la Faja Volcánica Transmexicana.
Provincia de las Serranías Meridionales. Incluye la Faja Volcánica Transmexicana, la Sierra Madre del Sur y el complejo montañoso del norte de Oaxaca.
Provincia de las Serranías Transístmicas. Abarca las montañas de Chiapas, continuando más allá de las fronteras de México hacia América Central.
Dentro del reino Neotropical, Rzedowski (1978) incluyó 2 regiones: Xerofítica Mexicana y Caribea. La región Xerofítica Mexicana comprende 5 provincias:
Provincia de Baja California. Área peninsular de Baja California.
Provincia de la Planicie Costera del Noroeste. Ocupa la mayor parte del estado de Sonora y se extiende por la costa de Sinaloa en forma de franja angosta.
Provincia de la Altiplanicie. Se extiende desde Chihuahua y Coahuila en el norte hasta Michoacán, Estado de México, Tlaxcala y Puebla en el sur, siendo la provincia más extensa del país.
Provincia de la Planicie Costera del Noreste. Comprende la zona fisiográfica homónima, en los estados de Tamaulipas, Nuevo León, Coahuila, San Luis Potosí y el norte de Veracruz, e incluye también una porción adyacente del estado de Texas en los EUA.
Provincia del Valle de Tehuacán-Cuicatlán. Corresponde a un área pequeña del sureste del estado de Puebla y porciones adyacentes de Oaxaca y Veracruz.
La región Caribea posee 6 provincias fitogeográficas:
Provincia de la Costa Pacífica. Se extiende en forma de franja angosta e ininterrumpida desde el este de Sonora y suroeste de Chihuahua en el norte hasta Chiapas en el sur, prolongándose a lo largo de la misma vertiente hasta América Central.
Provincia de las Islas Revillagigedo. Incluye las islas Socorro, San Benedicto y Clarión, situadas en el océano Pacífico, a más de 350 km del continente.
Provincia de la Depresión del Balsas. Se intercala entre la Faja Volcánica Transmexicana y la Sierra Madre del Sur, incluyendo partes de Jalisco, Michoacán, Estado de México, Guerrero, Morelos, Puebla y Oaxaca.
Provincia del Soconusco. Corresponde a una franja estrecha en las estribaciones inferiores de la Sierra Madre de Chiapas, prolongándose hacia Guatemala.
Provincia de la Costa del Golfo de México. Se extiende en forma de franja continua a lo largo de las partes bajas de Veracruz y Tabasco, además de algunas porciones adyacentes de Tamaulipas, San Luis Potosí, Hidalgo, Puebla, Oaxaca, Chiapas y probablemente de Campeche.
Provincia de la Península de Yucatán. Comprende la Península de Yucatán, una parte de Belice y el departamento de El Petén en Guatemala.
El sistema de Rzedowski (1978) ha sido el más empleado en los análisis fitogeográficos de las últimas décadas en México.
Arriaga et al. (1997)
En 1997 se llevó a cabo un taller sobre regionalización en la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio), en el que participaron 20 expertos, con el objetivo de generar un esquema biogeográfico de consenso. Inicialmente, los participantes del taller analizaron mapas temáticos con características de relieve, clima, temperatura media anual, régimen de lluvias y vegetación potencial, y los compararon con 4 sistemas biogeográficos compilados en 1990 en el "Atlas Nacional de México".
El sistema de Ferrusquía-Villafranca (1990), basado en rasgos morfotectónicos, posee 32 provincias, las cuales se clasifican en 2 dominios, 2 regiones y varias zonas de transición. El dominio Holártico (región Neártica) comprende las provincias Baja Californiense, Californiense, Sonorense, Sierra Madre Occidentalense,
Chihuahuense, Coahuilense, Tamaulipense, Sierra Madre Orientalense, Altiplanense, Neovolcanense, 8 zonas de extensión regional y áreas adyacentes con asociaciones bióticas complejas. Son transicionales las provincias San Lucasense, Sinaloense y Veracruzense. El dominio Neotropical (región Mesoamericana) comprende las provincias Nayaritense, Jalisqueño-Guerrerense, Oaxaquense, Pacifiquense, Campechano-Petenense, Yucatense, Chiapasense, 4 zonas de extensión regional y áreas adyacentes con asociaciones bióticas complejas.
El sistema de Rzedowski y Reyna-Trujillo (1990) incluye 16 provincias florísticas o fitogeográficas, clasificadas en 2 reinos y 4 regiones. El reino Holártico (región Pacífica Norteamericana) comprende las provincias de California y la Isla Guadalupe. El reino Neotropical comprende 3 regiones: la región Mesoamericana de Montaña, con las provincias de la Sierra Madre Occidental, Sierra Madre Oriental, Serranías Meridionales y Serranías Transístmicas; la región Xerofítica Mexicana con las provincias de Baja California, Planicie Costera del Noroeste, Altiplanicie,
Planicie Costera del Noreste y del Valle de TehuacánCuicatlán; y la región Caribeña con las provincias de la Costa Pacífica, Depresión del Balsas, Soconusco, Costa del Golfo de México y Península de Yucatán.
El sistema de Casas-Andreu y Reyna-Trujillo (1990), basado en especies de anfibios y reptiles, consta de 15 provincias herpetofaunísticas: Californiana, Desierto Colorado-Sonorense, Peninsular, del Cabo, Sierra Madre Occidental, Desierto Chihuahuense, Tamaulipeca, Eje Neovolcánico, Sierra Madre Oriental, Sierra Madre del Sur, Veracruzana, Petén, Mexicana del Oeste, Sierra Madre de Chiapas y Yucateca.
El sistema de Ramírez-Pulido y Castro-Campillo (1990), basado en especies de mamíferos, incluye 20 provincias mastofaunísticas clasificadas en 2 regiones. La región Neártica comprende las provincias de Baja California, Californiana, Sonorense, Sierra Madre Occidental, Chihuahuense, Sierra Madre Oriental, Tamaulipeca, Zacatecana y Volcánico-Transversal. La región Neotropical comprende las provincias de la Sierra Madre del Sur, Oaxaca-Tehuacanense, del Cabo, Sinaloense, Nayarita, del Balsas, Guerrerense, Chiapaneca, Yucateca y del Golfo.
Los 4 mapas correspondientes a los distintos sistemas fueron digitalizados y superpuestos con el objeto de determinar cuáles provincias eran compatibles. Las provincias resultantes se superpusieron con los mapas temáticos para ajustar sus límites. El sistema de consenso obtenido por Arriaga et al. (1997) incluye 19 provincias (Fig. 9), clasificadas en 3 medios. El medio árido subtropical incluye las provincias de California, Baja California, Cabo, Sonorense, Tamaulipeca, Altiplano Norte (Chihuahuense) y Altiplano Sur (Zacatecano-Potosino). El medio tropical húmedo y subhúmedo incluye las provincias de la Costa del Pacífico, Depresión del Balsas, Golfo de México, Yucatán y Petén. El medio montano incluye las provincias de la Sierra Madre Occidental, Sierra Madre Oriental, Eje Volcánico Transversal, Sierra Madre del Sur, Oaxaca, Altos de Chiapas y Soconusco. El esquema biogeográfico de Arriaga et al. (1997) resulta muy interesante, pues se basa en un consenso entre diferentes sistemas. Por ello, las provincias reconocidas generalmente se encuentran bien sustentadas, aunque algunas de ellas deberían poseer estatus de distrito (Morrone et al., 2002).

Figura 9 Provincias biogeográficas de México reconocidas por Arriaga et al. (1997). 1, California; 2, Baja California; 3, del Cabo; 4, Sonora; 5, Altiplano Norte; 6, Tamaulipeca; 7, Sierra Madre Occidental; 8, Sierra Madre Oriental; 9, Altiplano Sur; 10, Golfo de México; 11, Costa del Pacífico; 12, Eje Volcánico; 13, Depresión del Balsas; 14, Sierra Madre del Sur; 15, Yucatán; 16, El Petén; 17, Los Altos de Chiapas; 18, Soconusco; 19, Oaxaca.
Espinosa-Organista et al. (2008) revisaron el esquema anterior, proponiendo 20 provincias y clasificándolas de acuerdo con la jerarquización de Rzedowski (1978). La región Neártica comprende la provincia de California y la Zona de Transición Mexicana de Montaña, que a su vez incluye las provincias de la Sierra Madre Occidental, Sierra Madre Oriental, Eje Neovolcánico, Sierra Madre del Sur, Soconusco, Los Altos de Chiapas y Oaxaca. Dentro de la región Neotropical, Espinosa-Organista et al. (2008) reconocieron el Neotrópico árido del norte, con las provincias de Baja California, Cabo, Revillagigedo, Altiplano Norte (Chihuahuense), Altiplano Sur (Zacatecano-Potosino) y Sonorense, y el Neotrópico subhúmedo y húmedo de Mesoamérica, con las provincias del Pacífico, Depresión del Balsas, Tamaulipas, Golfo de México, Petén y Yucatán.
Morrone (2005, 2006)
En un esquema biogeográfico de América Latina y el Caribe, basado en distintos análisis de taxones de animales y plantas, reconocí 14 provincias biogeográficas para México, clasificadas en 2 regiones y una zona de transición (Morrone, 2005, 2006). La región Neártica comprende las provincias Californiana, de Baja California, de Sonora, del Desierto Chihuahuense y de Tamaulipas. La Zona de Transición Mexicana comprende las provincias de la Sierra Madre Occidental, Sierra Madre Oriental, Faja Volcánica Transmexicana, Sierra Madre del Sur y de las Tierras Altas de Chiapas. La región Neotropical comprende las provincias de las Tierras Bajas del Pacífico, Cuenca del Balsas y Veracruzana. Contribuciones posteriores han reconocido distritos dentro de algunas de estas provincias (Escalante et al., 2007; Torres-Miranda y Luna, 2007; Espinosa-Organista et al., 2008; Ramírez-Barahona et al., 2009; Duno-de Stefano et al., 2012; Gámez et al., 2012; Morrone, 2014a, 2017a, b).
Morrone et al. (2017) proporcionaron un mapa actualizado de las provincias biogeográficas mexicanas (Fig. 10). El delineado de este mapa y los shapefiles correspondientes fueron elaborados a partir del mapa de ecorregiones de Challenger y Soberón (2008), por lo que las provincias corresponden a áreas de endemismo a la vez que poseen identidad ecológica y fisiográfica. El esquema biogeográfico completo del país, hasta el nivel de distrito, que se presenta en esta contribución se resume en la Tabla 1.

Figura 10 Provincias biogeográficas de México (Morrone et al., 2017). 1, Californiana; 2, Baja California; 3, Sonora; 4, Desierto Chihuahuense; 5, Tamaulipas; 6, Península de Yucatán; 7, Sierra Madre Occidental; 8, Sierra Madre Oriental; 9, Faja Volcánica Transmexicana; 10, Cuenca del Balsas; 11, Sierra Madre del Sur; 12, Tierras bajas del Pacífico; 13, Veracruzana; 14, Tierras Altas de Chiapas.
Tabla 1 Esquema biogeográfico de México.
| Regiones | Provincias | Subprovincias | Distritos |
|---|---|---|---|
| Región Neártica | Californiana | ||
| Baja California | Desierto del Vizcaíno | ||
| Isla Guadalupe | |||
| Llanos de Magdalena | |||
| Santo Domingo | |||
| Sierra de La Laguna | |||
| El Cabo de Baja California | |||
| Sonora | |||
| Desierto Chihuahuense | Coahuilense | ||
| Zacatecano | |||
| Tamaulipas | |||
| Zona de Transición Mexicana | Sierra Madre Occidental | Apache | |
| Durangueño | |||
| Sierra Madre Oriental | Austro-Oriental | Saltillo-Parras | |
| Potosí | |||
| Austro-Oriental | Sierra Gorda | ||
| Zacualtipán | |||
| Faja Volcánica Transmexicana | Oeste | Otomí | |
| Tarasco | |||
| Este | Azteca | ||
| Orizaba-Zempoaltepec | |||
| Sierra Madre del Sur | Sierra Madre del Sur Occidental | Jalisciense | |
| Jalisciense-Manantlán | |||
| Sierra Madre del Sur Central | Michoacán | ||
| Sierra Madre del Sur Oriental | Guerrerense | ||
| Altiplanicie Oaxaqueña | |||
| Tierras Altas de Chiapas | Sierramadrense | ||
| Comitanense | |||
| Lacandón | |||
| Soconusco | |||
| Región Neotropical | Tierras Bajas del Pacífico | Sinaloense | |
| Islas Revillagigedo | |||
| Nayarit-Guerrero | |||
| Islas Tres Marías | |||
| Tehuano | |||
| Cuenca del Balsas | Balsas Inferior | ||
| Balsas Superior | |||
| Veracruzana | Bosque Deciduo del Norte de Veracruz | ||
| Los Tuxtlas | |||
| Valle de Chiapas | |||
| Bosque Lluvioso del Sur de Veracruz-Tabasco | |||
| Península de Yucatán | Norte de Yucatán | ||
| Rooeño | |||
| Petén |
Región Neártica
La región Neártica comprende las áreas templado-frías y áridas subtropicales de América del Norte, que abarcan Canadá, Groenlandia, los EUA y el norte de México (Morain, 1984; Morrone, 2006; Müller, 1986). Pertenece al reino Holártico, el cual incluye también a la región Paleártica, incluida en las áreas templado-frías del Viejo Mundo (Europa, Asia al norte del Himalaya y África al norte del Sahara). Desde una perspectiva paleogeográfica, el reino Holártico corresponde al paleocontinente de Laurasia.
La región Neártica se caracteriza por la presencia de varios taxones de plantas y animales (Espinosa-Organista et al., 2008; Müller, 1986). Las coníferas de los géneros Pinus, Abies y Juniperus son taxones endémicos, junto con las angiospermas del género Quercus. Entre los animales, destacan varias familias de mamíferos endémicos, como Aplodontiidae (castores de montaña), Geomyidae (tuzas o ratas de abazones), Heteromyidae (ratas canguro) y Antilocapridae (berrendos). Entre los reptiles se encuentran las Helodermatidae (monstruos de Gila) y las lagartijas del género Abronia; y entre los anfibios, las Ambystomidae (axolotes), Amphiumidae (salamandras) y Sirenidae (salamandras acuáticas).
La vegetación actual de esta región consiste en bosques templados en el este y oeste, tundra en el extremo norte, grandes praderas en el centro y matorrales xerófilos en el suroeste. La historia de la vegetación de la región Neártica ha sido moldeada por 2 eventos principales: las glaciaciones del Pleistoceno y la aridez del Holoceno (Morain, 1984). Durante el Pleistoceno, varios eventos de glaciación y los periodos interglaciales llevaron al avance y retroceso de los hielos hacia el sur, afectando la distribución de los bosques templados que permanecieron en refugios durante el avance de los hielos. El aumento de la aridez que se registró en el Holoceno llevó a una modificación de la parte más austral de la región Neártica, donde se desarrollaron los desiertos de Sonora, Chihuahua y la Península de Baja California.
Regionalización biogeográfica
Las 5 provincias biogeográficas más septentrionales de México -Californiana, de Baja California, de Sonora, del Desierto Chihuahuense y de Tamaulipas- se asignan a la región Neártica. Con excepción de la provincia de Baja California, que es exclusiva de nuestro país, las demás se extienden hacia el norte, abarcando parte de los EUA. Al analizar las relaciones bióticas de las provincias de Baja California, Sonora y del Desierto Chihuahuense, que poseen vegetación xerofítica, Rzedowski (1973) destacó sus afinidades con la vegetación tropical de otras áreas del mundo, en particular con la provincia del Monte, situada en el centro de la Argentina, que resultan incluso más marcadas que con las de las áreas áridas de los EUA. Por ello, Rzedowski (1978) consideró a estas provincias como parte de la región Xerofítica Mexicana, la cual asignó a la región Neotropical. Al sur de estas provincias, se encuentran las provincias montanas de la Sierra Madre Occidental, Sierra Madre Oriental, Faja Volcánica Transmexicana, Sierra Madre del Sur y Tierras Altas de Chiapas, las cuales pertenecen simultáneamente a la región Neotropical, por lo que se asignan a la llamada Zona de Transición Mexicana.
Escalante et al. (2018) realizaron un análisis de trazos de las especies de mamíferos de la región Neártica, con el objeto de identificar patrones de distribución en la región e investigar la existencia de áreas complejas. El análisis se basó en el "Atlas Biogeográfico de los Mamíferos Terrestres de América del Norte" (Escalante, 2013). Los autores construyeron los trazos individuales de 574 especies presentes exclusivamente en América del Norte. A partir de estos trazos individuales, construyeron una matriz de presencia-ausencia a partir de 323 cuadrantes ocupados por al menos una especie. El análisis de parsimonia de endemismos se llevó a cabo usando pesos implicados y la estrategia conocida como "eliminación progresiva de caracteres". El primer análisis de parsimonia produjo un cladograma totalmente resuelto, con 4 clados que fueron identificados como trazos generalizados por estar sustentados por la presencia de al menos 2 especies: Baja California, Altiplano Mexicano, Neotrópico y la costa noroeste de los EUA. Luego de la eliminación de las especies sustentando estos clados, se obtuvo un segundo cladograma, que permitió identificar otros trazos generalizados en el Altiplano de Columbia, Mesoamérica y el sur de las Montañas Rocosas. Un tercer análisis no permitió identificar más trazos generalizados. En las áreas donde los trazos generalizados obtenidos se superpusieron, los autores identificaron nodos o áreas complejas (Fig. 11). De acuerdo con Escalante et al. (2018), los resultados de este análisis corroboran la complejidad biogeográfica de la región Neártica. Al igual que otros análisis publicados por autores previos, es clara la diferenciación de patrones en el este y el oeste de la región. Los nodos encontrados, a su vez, coinciden con áreas geológicamente complejas, como la Sierra Madre de Chiapas, que pertenece a la Zona de Transición Mexicana.
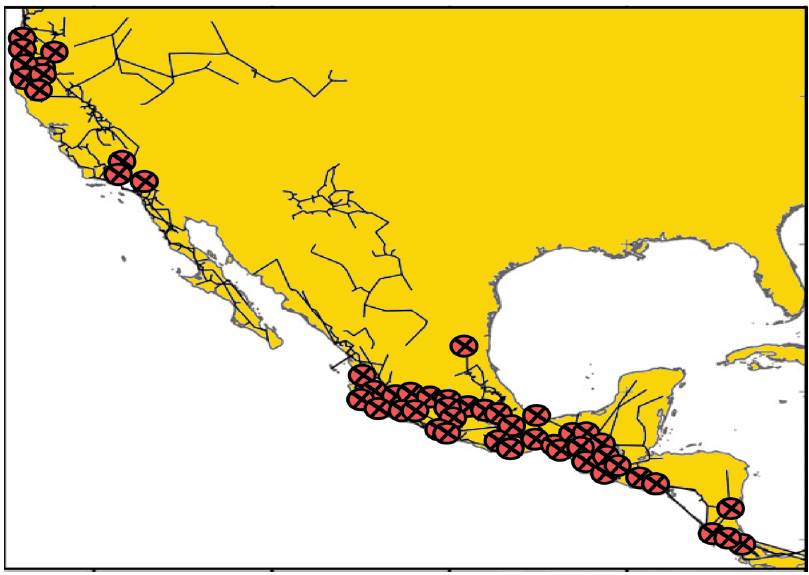
Figura 11 Trazos generalizados y nodos obtenidos por Escalante et al. (2018) para las regiones Neártica y Neotropical y la Zona de Transición Mexicana.
Evolución biótica
Numerosos análisis biogeográficos han analizado las relaciones de las regiones Neártica y Paleártica, que constituyen el reino Holártico. Sanmartín et al. (2001) llevaron a cabo un análisis biogeográfico cladístico, basado en la comparación de las filogenias de 57 taxones animales. Estos autores detectaron una separación básica entre las regiones Neártica y Paleártica, la cual se habría debido a vicarianza. Asimismo, encontraron que dentro de cada una de las regiones posteriormente ocurrieron eventos vicariantes entre sus porciones este y oeste. Al evaluar los posibles eventos de dispersión (Fig. 12), estos autores determinaron que las dispersiones continentales (entre oeste y este de la región Neártica o entre oeste y este de la región Paleártica) fueron significativamente más frecuentes que las dispersiones paleocontinentales (entre oeste de la región Neártica y este de la región Paleártica, y entre oeste de la región Paleártica o este de la región Neártica) y las disyuntas (entre este de la región Neártica y este de la región Paleártica, y entre oeste de la región Paleártica y oeste de la región Neártica). Esto implicaría que en la evolución biótica de la región Neártica existirían por un lado elementos endémicos que la caracterizarían como una unidad biótica natural y, por otro lado, elementos característicos del este y del oeste de la región, los cuales exhiben conexiones bióticas con distintas partes de la región Paleártica.

Provincia Californiana
Provincia Sandieguina Smith, 1941: 109.
Provincia de California Goldman y Moore, 1945: 350; Stuart, 1964: 350; Casas-Andreu y Reyna-Trujillo, 1990: mapa; Ramírez-Pulido y Castro-Campillo, 1990: mapa; Rzedowski y Reyna-Trujillo, 1990: mapa; Arriaga et al., 1997: 59; Brown et al., 1998: 30; Morrone, 2001: 17, 2005: 230; Espinosa-Organista et al., 2008: 49.
Provincia del Bosque Montano Cabrera y Willink, 1973: 30.
Provincia Californiana Rzedowski, 1978: 101; Morrone et al., 2017: 278.
Provincia Californiense Ferrusquía-Villafranca, 1990: mapa.
Ecorregión del Chaparral Costero de Salvia Californiano Dinerstein et al., 1995: 108.
Provincia de Baja California, N Escalante et al., 1998: 284.
Ecorregión de California Mediterránea Challenger y Soberón, 2008: 103.
Región Mediterránea González-Abraham et al., 2010: 75.
Esta provincia se ubica en el extremo septentrional de la península de Baja California, abarcando la mayor parte del estado de Baja California y extendiéndose a lo largo de la Sierra Nevada hasta el sudoeste de los EUA (Morrone, 2001, 2006). En esta provincia el sistema montañoso de Baja California alcanza su desarrollo máximo, con 2 sierras principales: San Pedro Mártir y Juárez, que alcanzan poco más de 3,400 m de altitud. Su clima varía de húmedo a semiseco y de frío a semicálido. En general el clima de las sierra es más templado y húmedo, y en la vertiente pacífica es más cálido y árido.
La vegetación característica de la provincia es el chaparral perennifolio, con especies de Adenostoma, Ceanothus, Quercus, Rhus y otros arbustos. También hay bosques de coníferas con Pinus lambertiana, Pseudotsuga macrocarpa, Abies concolor y Calocedrus recurrens; así como matorrales de Artemisia y bosques de encinos (Dinerstein et al., 1995; González-Abraham et al., 2010; Rzedowski, 1978).
Se considera que esta provincia es una de las que poseen un mayor número de plantas endémicas de la región Neártica (Morain, 1984). Algunos taxones endémicos son la selaginela azulada (Selaginella asprella), varias especies de encinos (Quercus agrifolia, Q. chrysolepis, Q. dumosa y Q. engelmanni), la rana arborícola californiana (Pseudacris cadaverina), una ardilla (Tamiasciurus mearnsi), el ratón californiano (Peromyscus californicus), el pájaro carpintero de Lewis (Melanerpes lewisi), el cuitlacoche californiano (Toxostoma redivivum) y el ratón californiano (Peromyscus californicus).
Evolución biótica. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999) y a un análisis filogeográfico comparativo basado en especies de aves (Zink, 2002), la provincia Californiana se relacionaría estrechamente con la provincia de Baja California. Otros análisis, en cambio, tratan a la provincia de Baja California como más relacionada con las provincias de Sonora y Desierto Chihuahuense, dejando así a la provincia Californiana como las más aislada de las provincias neárticas de México (Grismer, 1994; Riddle y Hafner, 2006). Su afinidad biótica con el resto de la región Neártica es evidente.
La provincia Californiana posee una flora muy interesante, que alberga 2 elementos bien caracterizados, asimilables a cenocrones. Por un lado, existen linajes antiguos o paleoendémicos, como Sequoia (Cupressaceae) y otros taxones relictuales, y por otro, linajes neoendémicos, como Arctostaphylos (Ericaceae) y Caenothus (Rhamnaceae), que han evolucionado en respuesta a eventos geológicos más recientes (Hauser et al., 2017). En relación con el desarrollo geológico del área, ha sido particularmente importante el levantamiento de la Sierra Nevada, en los últimos 10-20 ma, y el consecuente desarrollo de un clima mediterráneo. Con el objeto de contribuir a entender los patrones de diversificación de la flora de la provincia Californiana, Hauser et al. (2017) llevaron a cabo el análisis de las especies del género Quercus de la serie Agrifoliae. Los autores coleccionaron ejemplares de las 4 especies de la serie Agrifoliae, Quercus agrifolia, Q. kelloggii, Q. parvula y Q. wislizeni, además de algunas especies asignadas a otras series. Llevaron a cabo análisis morfométricos y extrajeron ADN para llevar a cabo un análisis filogenético molecular, utilizando el enfoque bayesiano. Adicionalmente, estimaron los tiempos de divergencia de los linajes analizados. Los análisis morfométricos de varios caracteres confirmaron la identidad de las especies estudiadas. El análisis filogenético confirmó la monofilia de la serie Agrifoliae y sus especies, a la vez que permitió confirmar la presencia de híbridos interespecíficos. El análisis de los tiempos de divergencia (Fig. 13) condujo a estimar la separación de Quercus kelloggii de las restantes especies en unos 20-25 ma, la de Q. agrifolia y Q. parvula-Q. wislizeni en 12-18 ma, y la de Q. parvula y Q. wislizeni en 8-14 ma. Los autores concluyeron que la serie Agrifoliae representa uno de los taxones más basales de encinos rojos, que comprende las 4 especies reconocidas y que hay evidencia de hibridación entre algunas de ellas. La diversificación de este taxón en el Mioceno Medio se relacionaría con la elevación de la Sierra Nevada, luego de la cual se habría producido una contracción de la distribución debido al enfriamiento y aridez progresiva del área. Esta modificación climática permitiría explicar la evolución de algunas de las especies analizadas.
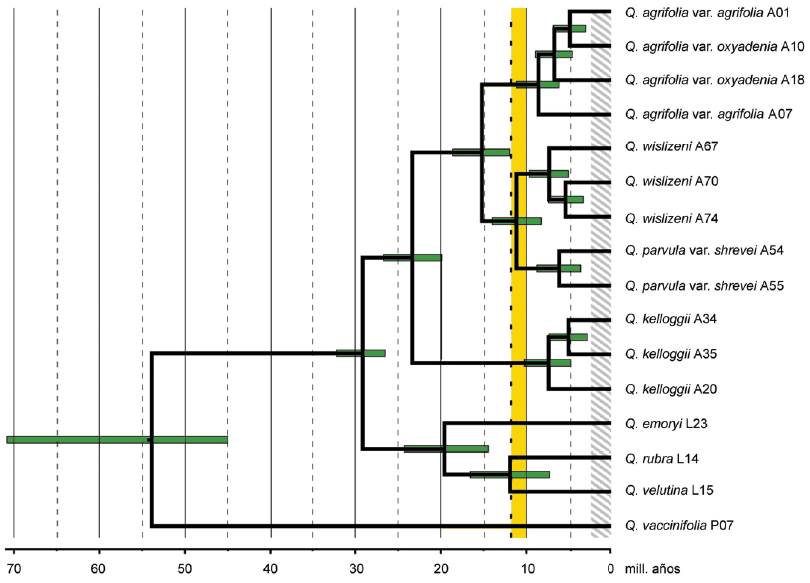
Figura 13 Análisis de los tiempos de divergencia de los encinos de la serie Agrifoliae (Hauser et al., 2017).
Conservación. La provincia Californiana se halla amenazada por el uso intensivo con fines turísticos, las presiones de los asentamientos humanos, la conversión de hábitats naturales para la agricultura y el sobrepastoreo (Dinerstein et al., 1995).
Provincia de Baja California
Provincia de Baja California Smith, 1941: 109; Goldman y Moore, 1945: 350; Rzedowski, 1978: 105; Ramírez-Pulido y Castro-Campillo, 1990: mapa; Rzedowski y Reyna-Trujillo, 1990: mapa; Arriaga et al., 1997: 60; Morrone, 2001: 18, 2005: 231; Espinosa-Organista et al., 2008: 59; Morrone et al., 2017: 278.
Provincia Sanlucasense (en parte) Stuart, 1964: 350.
Región de Baja California West, 1964: 368.
Distrito Sonoriano (en parte) Cabrera y Willink, 1973: 35.
Provincia Peninsular Casas-Andreu y Reyna-Trujillo, 1990: mapa.
Provincia Baja-Californiense Ferrusquía-Villafranca, 1990: mapa.
Provincia Baja Californiana Ramírez-Pulido y Castro-Campillo, 1990: mapa.
Provincia de San Pedro Mártir Álvarez-Castañeda et al., 1995: 80.
Ecorregión de los Bosques Secos de Baja California Dinerstein et al., 1995: 99.
Ecorregión del Matorral Xerófilo de Baja California Dinerstein et al., 1995: 109.
Provincia Sonorense (en parte) Brown et al., 1998: 32.
Provincia de Baja California, M Escalante et al., 1998: 284.
Ecorregión de los Desiertos de América del Norte, Desiertos Cálidos (en parte) Challenger y Soberón, 2008: 103.
Región Desértica González-Abraham et al., 2010: 75.
Esta provincia ocupa la mayor parte de la península de Baja California, al sur de la provincia Californiana. Posee una historia geológica compleja, ya que durante los últimos 5 ma se ha ido separando gradualmente de la costa de México debido a los movimientos diferenciales de las placas Norteamericana y Pacífica, a lo largo de la falla de San Andrés. Esta separación ocurrió en distintas etapas, con eventos de levantamiento, inmersión y fragmentación, que han tenido efectos importantes en la evolución biótica de la provincia (Grismer, 2000). Se halla recorrida por el sistema montañoso de Baja California, el cual continúa hacia el norte, atravesando la provincia Californiana, para unirse con las montañas de la Alta California, en particular con la Sierra Nevada (Morrone, 2001). El centenar de islas e islotes del Golfo de California en su mayoría se asignan a la provincia de la Península de Baja California. Case et al. (2002) compendian el conocimiento geológico y biológico de estas islas con varios capítulos referidos a sus plantas, insectos y vertebrados. De acuerdo con el momento de su formación, se distinguen 3 grupos de islas (Carreño y Helenes, 2002). El grupo septentrional incluye las islas más jóvenes, las cuales se formaron por sedimentación del delta del río Colorado o por actividad volcánica durante los últimos 2 ma. El grupo central incluye las islas que se comenzaron a formar a partir de sedimentos marinos en el Mioceno Medio (hace 14-12 ma) y emergieron entre el Pleistoceno y el Holoceno (hace 5-2 ma). El grupo austral incluye las islas graníticas o volcánicas, que emergieron cuando se abrió el extremo sur del Golfo de California (hace menos de 4 ma).
La vegetación de la provincia de Baja California incluye bosques secos, bosques de pino-encino y matorrales (Dinerstein et al., 1995; González-Abraham et al., 2010). Entre los géneros dominantes de plantas se encuentran Agave, Ambrosia, Coulterella, Gongylocarpus, Pachycormus, Pelucha y Yucca (Cabrera y Willink, 1973; Rzedowski, 1978).
Algunos taxones endémicos de esta provincia son: un helecho (Cheilanthes brandeeei), el ocotillo (Fouquieria diguetii), un mesquite (Prosopis palmeri), un encino (Quercus peninsularis), la serpiente de cascabel de Baja California (Crotalus enyo), el colín o codorniz californiana (Callipepla californica) y la rata canguro de Dulzura (Dipodomys simulans). Álvarez-Castañeda y Lorenzo (2017) reconocieron 3 subespecies de la liebre de California o de cola negra (Lepus californicus) endémicas de la Península de Baja California: L. californicus xanti al sur del Istmo de la Paz, L. californicus magdalenae entre el Istmo de la Paz y el sur del desierto del Vizcaíno, y L. californicus martirensis del desierto del Vizcaíno y el norte de la península.
Regionalización biogeográfica. Algunos autores han separado el área del Cabo, situada en el extremo austral de la península, como una provincia independiente de la de Baja California (Arriaga et al., 1997; Espinosa-Organista et al., 2008), mientras que otros la han considerado como parte de una misma unidad biogeográfica (ÁlvarezCastañeda et al., 1995; Morrone, 2006; Morrone et al., 2002). Rojas-Soto et al. (2003) mostraron, a partir de un análisis de parsimonia de endemismos basado en especies de aves, que a pesar de que esta área constituye una unidad natural, se halla anidada completamente dentro de las otras áreas de la Península de Baja California, por lo cual no se justificaría separarla como una provincia independiente. A partir del análisis de Álvarez-Castañeda et al. (1995), que consideró características climáticas, fisiográficas, de vegetación y la distribución de especies de mamíferos, es posible reconocer 6 distritos: Desierto del Vizcaíno, Isla Guadalupe, Llanos de Magdalena, Santo Domingo, Sierra de La Laguna y del Cabo de Baja California.
Distrito del Desierto del Vizcaíno, stat. nov.
Provincia del Desierto del Vizcaíno Goldman y Moore, 1945: 351; Álvarez-Castañeda et al., 1995: 82.
Subprovincia Vizcainoana Ferrusquía-Villafranca, 1990: mapa.
Ecorregión del Desierto de San Felipe o del Bajo Colorado González-Abraham et al., 2010: 77.
Ecorregión del Desierto del Vizcaíno González-Abraham et al., 2010: 79.
Corresponde al desierto situado en el norte de la provincia. Es el distrito más árido de la provincia.
Distrito de la Isla Guadalupe, stat. nov.
Provincia de la Isla Guadalupe Goldman y Moore, 1945: 350; Rzedowski, 1978: 101; Rzedowski y ReynaTrujillo, 1990: mapa; Escalante et al., 1998: 284.
Isla situada a 200 km de la costa. Puede considerarse como un distrito tomando en consideración la endemicidad de especies de plantas señalada por Rzedowski (1978). En esta isla se distribuye la única especie de encino restringida a islas, Quercus tomentella (Flores-Villela y Gerez, 1994), y la palma endémica Brahea edulis (Klimova et al., 2017).
Distrito de los Llanos de Magdalena, stat. nov.
Subprovincia Gigantana Ferrusquía-Villafranca, 1990: mapa.
Provincia de los Llanos de Magdalena Álvarez-Castañeda et al. 1995: 81.
Ecorregión del Desierto Central González-Abraham et al., 2010: 78.
Ecorregión de la Sierra de la Giganta González-Abraham et al., 2010: 78.
Ecorregión de los Llanos de Magdalena González-Abraham et al., 2010: 79.
Se extiende desde el sur del Desierto del Vizcaíno hasta la parte montañosa del área del Cabo. Incluye la Sierra de La Giganta. La vegetación es matorral sarcocaule.
Distrito de Santo Domingo, stat. nov.
Subprovincia San-Pedro-Martiriana Ferrusquía-Villafranca, 1990: mapa.
Provincia de Santo Domingo Álvarez-Castañeda et al. 1995: 82.
Provincia de San Pedro Mártir Álvarez-Castañeda et al., 1995: 80.
Ecorregión de la Costa Central del Golfo González-Abraham et al., 2010: 78.
Corresponde a la costa del Golfo de Cortés. La vegetación es matorral sarcocaule.
Distrito Sierra de La Laguna, stat. nov.
Provincia Sierra de La Laguna Álvarez-Castañeda et al. 1995: 81.
Ecorregión del Bosque de la Sierra de La Laguna González-Abraham et al., 2010: 80.
Área del Cabo que corresponde a la Sierra de La Laguna, por encima de los 1,200 m. Incluye una serie de cañones, cerros y valles, con vegetación predominante de pinos y encinos.
Distrito del Cabo de Baja California, stat. nov.
Provincia del Cabo de Baja California Smith, 1941: 109.
Provincia Sanlucasense (en parte) Stuart, 1964: 350.
Provincia del Cabo Casas-Andreu y Reyna-Trujillo, 1990: mapa; Ramírez-Pulido y Castro-Campillo, 1990: mapa; Arriaga et al., 1997: 60.
Provincia San-Lucasense Ferrusquía-Villafranca, 1990: mapa; Brown et al., 1998: 32.
Provincia de las Tierras Bajas de Los Cabos Álvarez-Castañeda et al., 1995: 81.
Ecorregión de los Bosques de Pino y Roble de San Lucas Dinerstein et al., 1995: 102.
Ecorregión del Mezquital de San Lucas Dinerstein et al., 1995: 109.
Provincia de Baja California, S Escalante et al., 1998: 284.
Ecorregión de las Selvas Cálido-Secas, Sierras y Planicies del Cabo Challenger y Soberón, 2008: 103.
Provincia del Cabo Espinosa-Organista et al., 2008: 59.
Región Tropical o del Cabo González-Abraham et al., 2010: 79.
Ecorregión del Matorral Tropical del Cabo González-Abraham et al., 2010: 79.
Ecorregión de la Selva Baja del Cabo González-Abraham et al., 2010: 79.
Se ubica en el extremo sur de la provincia y está delimitado por la Sierra de la Laguna, que se extiende desde el Cabo San Lucas hasta la Bahía de la Paz. Abundan los matorrales xerófilos y las selvas bajas caducifolias, existiendo pequeñas extensiones de bosques de pino-encino (Espinosa-Organista et al., 2008). Ejemplos de especies endémicas son: un pino piñonero (Pinus cembroides var. lagunae), un copal (Bursera ceracifolia), 2 especies de encinos (Quercus brandegeei y Q. devia), una serpiente de cascabel (Crotalus ruber lucasensis) y un colibrí (Hylocharis xantusii).
Evolución biótica. Las primeras teorías propuestas para explicar la evolución biótica de la península de Baja California, formuladas durante la primera mitad del siglo XX, asumieron eventos de dispersión a partir de áreas situadas al norte de la península. A partir de la década de 1970, los análisis comenzaron a incorporar la información de modelos geológicos, y así sugerir que la vicarianza tendría también un papel importante. Grismer (1994) llevó a cabo un análisis biogeográfico de la herpetofauna de la península de Baja California. Tomando en consideración la información filogenética disponible de numerosos taxones de lagartijas, geckos, serpientes y tortugas, junto con un modelo tectónico que explica la evolución geológica del área, Grismer (1994) postuló un modelo que contempla tanto dispersión como vicarianza. De acuerdo con éste, hace unos 4-4.5 ma, Baja California se comenzó a separar del oeste de México, formándose el Golfo de California y produciendo vicarianza entre las poblaciones situadas a ambos lados de éste. Asimismo, una inundación marina a lo largo del Istmo de La Paz, separó la región del Cabo del resto de la península, promoviendo también la vicarianza. En el Plioceno Medio, hace unos 3 ma, el Golfo de California estaba completamente formado, llegando en la parte norte a constituir una barrera efectiva para especies del noroeste de México y del sudeste de los EUA. En el Plio-Pleistoceno, hace 1-1.6 ma, comenzaron a elevarse montañas en la costa de California, con lo que el Golfo de California se redujo a su extensión actual y una tendencia al clima árido en la península llevó a la expansión de las áreas desérticas, a la dispersión de especies desde el desierto de Sonora y a la fragmentación de las especies peninsulares no adaptadas a condiciones de aridez. Finalmente, la región del Cabo se reconectó con las áreas del norte. Por otra parte, el desarrollo de los desiertos de Sonora, Mojave y Great Basin coadyuvó a la fragmentación de las especies de la Península de Baja California de las del suroeste de EUA y el noroeste de México.
Un análisis biogeográfico de los mamíferos asociados con las áreas desérticas de la Península de Baja California (Hafner y Riddle, 1997) concluyó que su biota indicaba una antigua historia evolutiva separada de la del desierto de Sonora. De acuerdo con estos autores, algunos linajes de mamíferos peninsulares habrían comenzado a evolucionar en el Mioceno Medio al separarse la península del continente. A lo largo del Mioceno Tardío y el Plioceno Temprano se habrían formado semidesiertos, aislados de otros semidesiertos continentales por ingresiones marinas. Durante el Pleistoceno Tardío las especies peninsulares se habrían aislado en refugios. La alternancia de periodos de aislamiento y de contacto secundario por dispersión explicaría la complejidad biótica de la Península de Baja California.
Otro análisis basado en especies de mamíferos, aves, reptiles, anfibios y cactos (Riddle y Hafner, 2006) permitió postular un escenario biogeográfico complejo. Estos autores llevaron a cabo su análisis en 5 etapas. La primera etapa consistió en delinear la biota de interés e identificar los taxones a analizar. Estudios filogeográficos previos determinaron que los eventos de diversificación de los desiertos cálidos norteamericanos (Chihuahuense, Sonora, Mojave y de la Península de Baja California) preceden al Pleistoceno Tardío, correspondiendo en la mayor parte de los casos al Plioceno y Mioceno. De acuerdo con esto, los autores seleccionaron varios taxones de estos desiertos para los cuales había hipótesis filogenéticas robustas basadas en caracteres moleculares. La segunda etapa del análisis consistió en determinar las áreas de endemismo que constituirían las unidades del análisis. Para ello llevaron a cabo un análisis de parsimonia de endemismos basado en 74 taxones (31 mamíferos, 27 aves, 11 reptiles, 3 anfibios y 2 plantas). Como resultado identificaron 4 áreas de endemismo (Fig. 14a): Continental este (Trans-Pecos, Coahuila y Zacatecas), Continental oeste (Sonora y Colorado), Peninsular sur (Magdalena y San Lucas) y Peninsular norte (Cirios). En la siguiente etapa, determinaron la estructura general de la divergencia entre las áreas, a partir de un análisis de parsimonia de Brooks primario, utilizando las filogenias disponibles para los taxones de la etapa anterior. El cladograma general de áreas resultante (Fig. 14b) mostró que las 2 áreas continentales y las 2 áreas peninsulares eran grupos hermanos, respectivamente. Cabe destacar que las áreas Continental oeste, Continental este y Peninsular norte resultaron con un elevado número de homoplasias, es decir, que las mismas especies aparecen en 2 áreas no relacionadas bióticamente entre sí. Con el objeto de resolver las diferencias entre los resultados de las divergencias observadas en la etapa anterior, los autores procedieron a un análisis de parsimonia de Brooks secundario. En éste, a partir de duplicar las áreas Continental oeste, Continental este y Peninsular norte, se obtuvo un cladograma general de áreas (Fig. 14c), que muestra una primera disyunción entre las áreas Continental oeste 1 y Continental este 1. Una segunda disyunción separa las áreas Peninsular norte 2 y Peninsular sur de las áreas Peninsular norte 1, Continental oeste 2 y Continental este 2. Cada una de estas disyunciones puede ser asociada con un evento geológico (Fig. 14d):
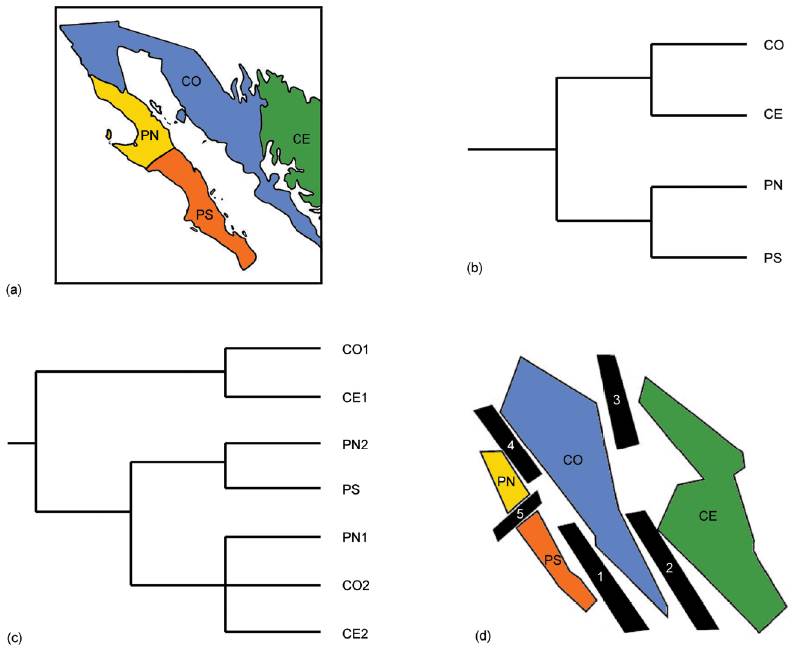
Figura 14 Análisis biogeográfico evolutivo de la Península de Yucatán y áreas adyacentes llevado a cabo por Riddle y Hafner (2006). (a) Áreas de endemismo; (b) cladograma general de áreas resultante del análisis de parsimonia de Brooks primario; (c) cladograma general de áreas resultante del análisis de parsimonia de Brooks secundario; (d) eventos geológicos que habrían inducido la vicarianza. CE= Continental Este; CO= Continental Oeste; PN= Peninsular Norte; PS= Peninsular Sur; 1, apertura del Golfo Trans-Golfo; 2, formación del Altiplano Mexicano; 3, lecantamiento de la Sierra Madre Occidental; 4= embahiamientos Trans-Golfos; 5, pasaje marino del Vizcaíno.
Disyunción basal. Apertura del Golfo Trans-Golfo, formación de la planicie mexicana y embahiamientos Trans-Golfos.
Disyunción Continental oeste 1 y Continental este 1. Levantamiento de la Sierra Madre Occidental.
Disyunción Peninsular norte 2-Peninsular sur y Peninsular norte 1. Pasaje marino del Vizcaíno.
El levantamiento de la Sierra Madre Occidental, la formación del Altiplano Mexicano, la apertura del Golfo de California y el paisaje marino del Vizcaíno habrían inducido diferentes eventos vicariantes sucesivos combinados con dispersión. Los eventos vicariantes implicarían una separación inicial entre los desiertos Chihuahuense y Sonorense, luego una separación entre los desiertos continentales y el norte de la Península de Baja California y una separación más reciente entre el norte y el sur de la Península de Baja California. Los eventos de dispersión implicarían movimientos desde el sur al norte de la península. En la última etapa del análisis los autores contrastaron hipótesis de taxones particulares a partir de los resultados anteriores. Algunos taxones mostraron eventos de dispersión a lo largo de la Península de Baja California (Polioptila californica y Lophocereus schotii), mientras que otros fueron compatibles con eventos vicariantes (Dipodomys merriami, Auriparus flavipes y Campylorhynchus brunneicapillus). En conclusión, las afinidades más evidentes de la provincia de Baja California son con otras provincias de la región Neártica, especialmente las que poseen vegetación xérica (de Sonora y del Desierto Chihuahuense). Otras conexiones con áreas neotropicales, sin embargo, no se pueden descartar, sobretodo en su extremo más austral (distrito de El Cabo).
Conservación. La provincia de Baja California se halla amenazada por la conversión de hábitats naturales para la agricultura, la extracción forestal del árbol de boojum (Fouquieria columnaris), el sobrepastoreo y la caza excesiva (Dinerstein et al., 1995). Una gran parte de su superficie presenta signos de perturbación (Flores-Villela y Gerez, 1994). En esta provincia se incluyen varias áreas naturales protegidas (Conanp, 2018):
Reserva de la Biosfera El Vizcaíno. Situada en la parte central de la provincia (Ortega y Arriaga, 1991). Es el sitio de mayor importancia mundial para la conservación de la ballena gris y el berrendo peninsular. En ella se encuentran particularmente representados diversos tipos de matorral desértico, como matorral sarcocaule, matorral sarcocrasicaule, matorral halófilo, matorral de dunas, matorral inerme y vegetación de dunas costeras y eriales (León-de la Luz et al., 1991). Especies representativas son el cardón (Pachycereus pringlei), el cirio (Fouquieria columnaris), el berrendo peninsular (Antilocapra americana peninsularis), el borrego cimarrón (Ovis canadensis cremnobates) y el águila real (Aquila chrysaetos).
Reserva de la Biosfera Isla Guadalupe. Posee un alto nivel de endemismo de la flora. Incluye bosques de coníferas, matorral xerófilo, palmar natural y vegetación inducida. Algunas especies representativas son el ciprés de Guadalupe (Cupressus guadalupensis), la palma de Guadalupe (Brahea edulis), la biznaga de Blossfeld (Mammillaria blossfeldiana), el enebro de California (Juniperus californica) y el junco de Guadalupe (Junco insularis). Alberga colonias reproductoras de mamíferos marinos, como el lobo fino de Guadalupe (Arctocephalus townsendi), el elefante marino (Mirounga angustirostris) y el lobo marino de California (Zalophus californianus).
Reserva de la Biosfera Sierra de La Laguna. Esta región tiene un grupo de ecosistemas diferentes como bosques de coníferas, selvas tropicales, palmares y matorrales, únicos en toda la Península de Baja California. Constituye el límite occidental de la selva seca en el hemisferio occidental (Arriaga, 2010). Especies representativas son el pino piñonero (Pinus cembroides var. lagunae), el guerivo o álamo (Populus brandegeei), un encino (Quercus tuberculata), el mauto (Lysiloma divaricatum), una biznaga (Ferocactus towsendianus), el venado bura peninsular (Odocoileus hemionus peninsulae), el colibrí de xantus o chuparrosa (Hylocharis xantusii) y la codorniz californiana (Callipepla californica).
Área de Protección de Flora y Fauna Cabo San Lucas. Zona de refugio y alimentación de diversas especies de peces marinos, como el marlin azul, dorado y pez vela, especies de gran importancia para la pesca deportiva. Especies representativas son una biznaga (Ferocactus towsendianus), el torote blanco (Bursera microphylla) y el lobo marino californiano (Zalophus californianus).
Provincia de Sonora
Provincia Arizoniana Smith, 1941: 109.
Provincia Sinaloense Smith, 1941: 109; Álvarez-Castañeda et al., 1995: 79.
Provincia de Sonora Goldman y Moore, 1945: 352; Ramírez-Pulido y Castro-Campillo, 1990: mapa; Arriaga et al., 1997: 60; Morrone, 2001: 19, 2005: 231; Espinosa-Organista et al., 2008: 60; Morrone et al., 2017: 278.
Provincia de Sinaloa Moore, 1945: 218.
Provincia Sonorense Stuart, 1964: 350; Ferrusquía-Villafranca, 1990: mapa; Brown et al., 1998: 32.
Región de Sonora y el Norte de Sinaloa West, 1964: 368.
Distrito Sonoriano (en parte) Cabrera y Willink, 1973: 35.
Provincia de la Planicie Costera del Noroeste Rzedowski, 1978: 106; Rzedowski y Reyna-Trujillo, 1990: mapa.
Provincia del Desierto Colorado-Sonorense Casas-Andreu y Reyna-Trujillo, 1990: mapa.
Provincia Guaymensis Álvarez-Castañeda et al., 1995: 81.
Ecorregión del Matorral de Cactos del Norte de Sonora Dinerstein et al., 1995: 110.
Ecorregión del Matorral Xerófilo de Sonora Dinerstein et al., 1995: 110.
Ecorregión del Mezquital Occidental de México Dinerstein et al., 1995: 109.
Área del Desierto de Sonora Marshall y Liebherr, 2000: 204.
Ecorregión de los Desiertos de América del Norte, Desiertos Cálidos (en parte) Challenger y Soberón, 2008: 103.
Esta provincia abarca la Planicie Costera Noroccidental de México, desde la porción nororiental de la península de Baja California hasta la cuenca del río Piaxtla en el sur, ocupando la mayor parte del estado de Sonora, la costa del estado de Sinaloa, y porciones de los estados de Arizona y California en los EUA (Morrone, 2001, 2006). Es un área relativamente baja, que no excede los 1,000 m de altitud, con llanuras interrumpidas por montañas bajas y dunas. La Planicie Costera Noroccidental de México limita al este con la Sierra Madre Occidental, existiendo en su parte septentrional un área de transición con elevaciones menores y pequeñas llanuras.
Predominan los matorrales xerófilos de Larrea tridentata y Ambrosia dumosa, alternando con bosques de cactos gigantes (Cabrera y Willink, 1973; Dinerstein et al., 1995). Entre los géneros dominantes de plantas se encuentran: Acacia, Agiabampoa, Bursera, Canotia, Carnegiea, Cercidium, Forchammeria, Ipomoea, Lysiloma, Olneya, Pachycereus, Prosopis y Yucca (Rzedowski, 1978).
Algunos taxones endémicos de esta provincia son el romerillo (Leucophyllum pringlei), el saguaro (Carnegeia gigantea), un escarabajo (Cotinis producta), una serpiente de cascabel (Crotalus cerastes), el gorrión bigotudo sonorense (Aimophila carpalis), una musaraña (Sorex arizonae), el chichimoco (Amnospermophilus harrisii), una rata canguro (Dipodomys deserti) y el ratón de abazones (Chaetodipus goldmani).
Evolución biótica. La flora del desierto de Sonora fue analizada por Axelrod (1979), quien concluyó que ésta evolucionó durante el Plioceno-Pleistoceno, a partir de comunidades que se desarrollaron en el área durante el Cretácico Tardío y Paleógeno (sabanas, pastizales, bosques secos y bosques de pino-encino). Rzedowski (1978) consideró que la flora de la provincia de Sonora es muy semejante a la de la provincia de Baja California, aunque con un menor número de endemismos.
Las afinidades bióticas principales de esta provincia son con las restantes provincias de la región Neártica. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia de Sonora se relaciona estrechamente con las provincias de la Sierra Madre Occidental y Tamaulipas, así como con la porción norte de la provincia del Desierto Chihuahuense. Un análisis filogeográfico comparado basado en especies de aves (Zink, 2002) postuló que la biota de esta provincia se relaciona con la de la provincia del Desierto Chihuahuense.
Un taxón interesante es el género de tortugas terrestres Gopherus, el único representante de las Testudinidae en América del Norte. Posee 6 especies, distribuidas en el sur de los EUA y norte de México. La especie G. morafkai es endémica del desierto de Sonora. Edwards et al. (2016) analizaron el papel de distintos eventos históricos en la distribución geográfica de esta especie y la asociación de distintos linajes con diferentes tipos de vegetación. A partir de 133 muestras de sangre de distintas localidades de México y Arizona, los autores aislaron ADN genómico y secuenciaron un gen mitocondrial y microsatélites. A partir de las secuencias, procedieron a reconstruir una hipótesis genealógica matrilineal y estimaron los tiempos de divergencia. Por otra parte, analizaron mapas de vegetación para estimar la distribución de los especímenes en bosque deciduo tropical, matorral espinoso de Sinaloa, matorral desértico de Sonora y sus ecotonos. La reconstrucción genealógica obtenida mostró una mayor relación entre los especímenes de Sinaloa con los del desierto del Mojave, mientras que los de Sonora resultaron el grupo hermano de los anteriores (Fig. 15). El tiempo de divergencia basal fue estimado en 5.77 ma. Los autores consideraron que los 2 linajes principales, a los que denominaron Sonorense y Sinaloense, se asocian con tipos de vegetación diferente, y se encuentran mezclados en los ecotonos, donde existe hibridación limitada. Dado que no existen barreras geográficas que restrinjan el flujo génico entre los linajes Sonorense y Sinaloense, Edwards et al. (2016) estimaron que esta especie habría evolucionado de acuerdo con un modelo parapátrido en respuesta a la adaptación a nichos ecológicos específicos. Al igual que otros taxones de la provincia de Sonora, el ancestro común de los 3 linajes se habría expandido en el Mioceno (23.7-5.3 ma) cuando el bosque tropical cubrió el área. Cuando la aridez se intensificó hace 15-8 ma, aparecieron nuevas condiciones en la porción norte y condujeron a la divergencia entre zonas áridas y tropicales. Sucesivas expansiones y contracciones de los ambientes áridos y tropicales del Pleistoceno habrían influenciado de manera diferencial a los linajes. El linaje Sinaloense se habría beneficiado con los inviernos suaves, en que el matorral espinoso y el bosque tropical subcaducifolio habrían persistido y se habrían expandido en los periodos glaciales. El linaje Sonorense, a su vez, se habría beneficiado de los interglaciales más calientes y secos, penetrando de manera más eficiente en el matorral espinoso.
Figura 15 Reconstrucción genealógica de los linajes de la tortuga del desierto de Sonora, Gopherus morafkai (Edwards et al., 2016).
Conservación. La provincia de Sonora se halla amenazada por la irrigación, los ranchos ganaderos, la extracción de leña, la explotación de la vida silvestre, la caza excesiva y el uso intensivo con fines turísticos (Dinerstein et al., 1995). Un análisis reciente de modelado de nicho ecológico de Carnegeia gigantea a escenarios de incremento de temperatura (Albuquerque et al., 2018) estimó que la distribución geográfica de este cacto podría contraerse de continuar las tendencias actuales de cambio climático. En esta provincia se incluyen varias áreas naturales protegidas (Conanp, 2018):
Reserva de la Biosfera Alto Golfo de California y Delta del Río Colorado (Baja California y Sonora). Posee matorral xerófilo, matorral desértico micrófilo, vegetación de desiertos arenosos, vegetación halófila y vegetación hidrófila. Algunas especies representativas son el tule (Typha domingensis), la gobernadora (Larrea tridentata), el pasto salado o nipa (Distichlis palmeri), la sosa (Suaeda puertopanascoa), el pez cachorrito del desierto (Cyprinodon macularius) y la lagartija cornuda cola plana (Phrynosoma mcallii).
Reserva de la Biosfera El Pinacate y Desierto de Altar (Sonora). Contiene elementos geomorfológicos únicos como cráteres, campos de lava y un escudo volcánico de reciente formación que en conjunto le dan la unicidad al sitio, el cual fue incluido por la UNESCO en la lista de Sitios Patrimonio de la Humanidad. La vegetación es matorral xerófilo, pastizal, vegetación hidrófila y vegetación inducida.
Reserva de la Biosfera Isla San Pedro Mártir (Sonora). La isla San Pedro Mártir es un sitio de gran riqueza biológica. Su vegetación es matorral xerófilo. Habitan esta isla 8 especies de aves marinas, entre ellas el bobo de patas azules (Sula nebouxi), el bobo café (Sula leucogaster) y el pelícano pardo (Pelecanus occidentalis).
Provincia del Desierto Chihuahuense
Provincia Chihuahuense Smith, 1941: 109; Ferrusquía-Villafranca, 1990: mapa; Ramírez-Pulido y Castro-Campillo, 1990: mapa; Brown et al., 1998: 30.
Provincia de Chihuahua-Zacatecas Goldman y Moore, 1945: 354; Stuart, 1964: 350.
Provincia del Desierto Chihuahense Moore, 1945: 218; Morrone et al., 2017: 278.
Distrito Chihuahuano (en parte) Cabrera y Willink, 1973: 35.
Provincia de la Altiplanicie Rzedowski, 1978: 106.
Provincia del Desierto de Chihuahua Casas-Andreu y Reyna-Trujillo, 1990: mapa.
Provincia del Altiplano Rzedowski y Reyna-Trujillo, 1990: mapa.
Ecorregión del Chaparral del Interior de México Dinerstein et al., 1995: 110.
Ecorregión del Matorral de Cactos del Centro de México Dinerstein et al., 1995: 110.
Ecorregión del Matorral Oriental de México Dinerstein et al., 1995: 110.
Ecorregión del Matorral Xerófilo de Chihuahua Dinerstein et al., 1995: 110.
Ecorregión del Mezquital del Centro de México Dinerstein et al., 1995: 110.
Ecorregión de los Pastizales del Centro de México Dinerstein et al., 1995: 103.
Provincia del Altiplano Mexicano Campbell, 1999: 115; Morrone, 2001: 22, 2005: 232.
Esta provincia se sitúa en el Altiplano Mexicano, que se extiende entre las Sierras Madre Occidental y Oriental, abarcando los estados de Zacatecas, San Luis Potosí, Guanajuato, Chihuahua, Jalisco, Michoacán, Tlaxcala, Puebla, Coahuila, Durango, Nuevo León y Sonora, y el sur de los estados de Nuevo México y Texas en los EUA (Morrone, 2001). La altitud del Desierto Chihuahuense varía entre los 1,000 y 2,000 m. En él abundan las cuencas endorreicas, algunas son relativamente grandes, como las de los ríos Nazas, Aguanaval y Casas Grandes; y otras más reducidas, como las que en conjunto constituyen los Bolsones de Mapimí y del Salado.
La vegetación de la provincia del Desierto Chihuahuense consiste en estepas de gramíneas de los géneros Bouteloua y Aristida, extendidas entre matorrales xerófilos, y bosques en los llanos y valles (Cabrera y Willink, 1973; Dinerstein et al., 1995). Entre las especies dominantes de plantas se encuentran Andropogon citratus, Aristida glauca, Bouteloua aristidoides, B. eriopoda, B. hirsuta y Larrea tridentata (Cabrera y Willink, 1973; Katinas et al., 2004).
Algunos taxones endémicos de esta provincia son el copalillo (Parthenium incanum), un carábido (Agonum extimum), la rata canguro (Dipodomys nelsoni), la rata magueyera (Neotoma goldmani) y el ratón de abazones sedoso (Perognathus flavus).
Regionalización biogeográfica. Varios autores han considerado la existencia de 2 provincias diferentes: Altiplano Norte y Altiplano Sur (Arriaga et al., 1997; Espinosa-Organista et al., 2008). Un análisis de endemicidad basado en especies de lagartijas (Ocampo-Salinas et al., 2019) ha corroborado la existencia de estas áreas, las cuales considero aquí como los distritos Coahuilense y Zacatecano.
Distrito Coahuilense, stat. nov.
Región de la Mesa del Norte West, 1964: 368.
Provincia Coahuilense Ferrusquía-Villafranca, 1990: mapa.
Provincia de Coahuila Ramírez-Pulido y CastroCampillo, 1990: mapa.
Provincia del Altiplano Norte Arriaga et al., 1997: 61.
Provincia del Desierto de Chihuahua, N Escalante et al., 1998: 284.
Ecorregión de los Desiertos Cálidos de América del Norte (en parte) Challenger y Soberón, 2008: 103.
Provincia del Altiplano Norte (Desierto Chihuahuense) Espinosa-Organista et al., 2008: 60.
Abarca desde el río Nazas hasta el Big Bend, en el sur de los EUA. Es más árido y predomina la vegetación de matorral xerófilo. Algunas especies endémicas son 2 helechos (Cheilanthes pringlei var. moncloviensis y Notholaena greggii), un pino (Pinus remota), el guayule (Parthenium argenteum), la biznaga partida de espinas curvas (Coryphantha recurvata), el peyote (Lophophora williamsii), 2 especies de lagartijas de árbol (Urosaurus ornatus caeruleus y U. ornatus schmidti) y un ratón (Peromyscus eremicus eremicus). Distrito Zacatecano, stat. nov.
Región de la Mesa Central West, 1964: 368.
Provincia Altiplanense Ferrusquía-Villafranca, 1990: mapa.
Provincia Zacatecana Ramírez-Pulido y CastroCampillo, 1990: mapa.
Provincia del Altiplano Sur (Zacatecano-Potosino) Arriaga et al., 1997: 62.
Provincia del Desierto de Chihuahua, S Escalante et al., 1998: 284.
Ecorregión de las Elevaciones Semiáridas Meridionales Challenger y Soberón, 2008: 103.
Provincia del Altiplano Sur (Tierras Áridas de Zacatecas-Mezquital) Espinosa-Organista et al., 2008: 60.
Desde el valle del Mezquital hasta al norte del río Aguanaval. Es menos seco que el distrito anterior. Ejemplos de taxones endémicos son: un helecho (Pellaea ribae), una variedad de enebro (Juniperus deppeana var. zacatecensis), 2 biznagas (Coryphantha macromeris runyonii y C. nickelsiae) y 1 lagartija (Sceloporus torquatus melanogaster).
Evolución biótica. Se ha propuesto que el Desierto Chihuahuense posee muchas especies de insectos de origen neotropical antiguo, que se dispersaron desde América del Sur en el Mioceno, sobrevivieron y se diversificaron en el Desierto Chihuahuense durante el Eoceno-Plioceno, a la vez que se extinguieron en otras áreas (Halffter, 1964, 1976, 2017). A estos elementos neotropicales antiguos se sumaron elementos holárticos durante el Plio-Pleistoceno (Halffter, 1964). La fauna de vertebrados del Desierto Chihuahuense, en contraposición, es típicamente neártica. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia del Desierto Chihuahuense se relaciona con la provincia de la Sierra Madre Oriental en su porción sur y con las provincias de la Sierra Madre Occidental y Sonora en su porción norte. De acuerdo con un análisis filogeográfico comparado basado en especies de aves (Zink, 2002), esta provincia se relaciona con la provincia de Sonora.
El género Agave (Asparagaceae) es uno de los elementos más característicos de los desiertos norteamericanos. La especie A. lechuguilla es una especie característica del Desierto Chihuahuense. Scheinvar et al. (2017) analizaron la estructura filogeográfica de esta especie con el propósito de analizar la influencia de factores climáticos y orográficos del pasado en su evolución, en particular para determinar la existencia de refugios durante el Pleistoceno. Los autores analizaron 15 poblaciones de A. lechuguilla que representaban la totalidad del área de distribución de la especie. Se les extrajo el ADN y se amplificaron y secuenciaron 4 fragmentos de 289 individuos. Se analizó la diversidad genética y la demografía, además de estimarse los tiempos de divergencia. Asimismo construyeron modelos de nicho ecológico para la actualidad, el Último Periodo Interglacial (120,000-140,000 años) y el Último Máximo Glacial (21,000 años), y se los superpuso con el objeto de identificar áreas estables que podrían haberse comportado como refugios. La diversidad genética encontrada fue alta, pero la diversidad intrapoblacional fue mucho menor, lo que indicaría una fuerte diferenciación entre las poblaciones. La red de haplotipos permitió identificar un grupo restringido al Altiplano Norte, otro en el sur restringido a la región del río Pánuco, uno central en la región de Mapimí y otro septentrional-central en las regiones Saladense y Trans-Pecos. El análisis demográfico detectó una expansión en el grupo más austral, que habría ocurrido en el Pleistoceno. La estimación de los tiempos de divergencia (Fig. 16) indicó que A. lechuguilla se habría originado hace 4.46 ma. La divergencia del grupo del Altiplano Norte habría ocurrido en la transición entre el Neógeno y el Cuaternario, mientras que los otros grupos habrían divergido en el Pleistoceno. De acuerdo con Scheinvar et al. (2017), A. lechuguilla se habría originado en el Neógeno, con 2 linajes ancestrales que corresponderían al Altiplano Norte y el Altiplano Sur. Durante el Pleistoceno, habría ocurrido la diversificación de los otros linajes, asociada con los cambios climáticos que habrían permitido el aislamiento de las poblaciones de especies de ambientes áridos en refugios. Cuando las condiciones lo permitieron, estas poblaciones se expandieron y pusieron en contacto, incrementando la variabilidad genética.

Figura 16 Estimación de los tiempos de divergencia de Agave lechuguilla y especies afines desde el Mioceno hasta el Holoceno (Chseinvar et al., 2017).
Conservación. La provincia del Desierto Chihuahuense se halla amenazada por el sobrepastoreo y la consiguiente invasión de especies arbustivas, la extracción de sal y yeso, y la explotación de los recursos hídricos (Dinerstein et al., 1995). En esta provincia se incluyen varias áreas naturales protegidas (Conanp, 2018):
Reserva de la Biosfera Janos (Chihuahua). Gran extensión de pastizales en muy buen estado de conservación, lo que la hace la mayor área natural protegida representativa de estos ecosistemas en el país, asegurando también una conectividad de este hábitat en América del Norte. También incluye bosques de encino, bosques de encino-pino, bosques de pino, chaparral, pastizal halófilo y vegetación riparia. Se destaca por la migración de la única manada silvestre de bisonte americano (Bison bison) en México y el suroeste de los EUA y por la reproducción de la cotorra serrana occidental (Rhynchopsitta pachyrhyncha) y del oso negro de la Sierra Madre Occidental (Ursus americanus machetes). Asimismo, destaca por mantener la mayor población reproductiva del tecolote llanero (Athene cunicularia) en pastizales nativos norteamericanos y las colonias más importantes de perros llaneros de cola negra (Cynomys ludovicianus). Representa el sitio con mayores posibilidades para el establecimiento de una población silvestre viable de hurón de patas negras (Mustela nigripes).
Reserva de la Biosfera La Michilía (Durango). Comprende zonas montañosas, semidesérticas y ambientes complejos desde el punto de vista ecológico y topográfico. Incluye praderas de alta montaña, matorral espinoso tamaulipeco y selva baja espinosa subcaducifolia. Dos especies representativas son la abelia (Abelia occidentalis) y el viejito (Mammillaria senilis).
Reserva de la Biosfera Mapimí (Coahuila y Chihuahua). Se destaca por ser área de alimentación y refugio de aves migratorias de pastizal y hábitat de aves migratorias acuáticas y por contener el hábitat del ciclo de vida completo de la tortuga del bolsón (Gopherus flavomarginatus), el área de alimentación y descanso de la migración de la aguililla de Swainson (Buteo swainsoni) y el área de reproducción del águila real (Aquila chrysaetos). Barbault y Halffter (1981) editaron un libro con análisis de varios taxones de vertebrados de esta reserva (tortuga del bolsón, lagartijas, mamíferos pequeños y aves).
Provincia de Tamaulipas
Provincia Tamaulipeca Smith, 1941: 109; Arriaga et al., 1997: 60; Espinosa-Organista et al., 2008: 61.
Provincia de Tamaulipas Goldman y Moore, 1945: 354;
Casas-Andreu y Reyna-Trujillo, 1990: mapa; Ramírez-Pulido y Castro-Campillo, 1990: mapa; Rzedowski y Reyna-Trujillo, 1990: mapa; Morrone, 2001: 23, 2005: 233; Morrone et al., 2017: 278.
Provincia Tamaulipense Stuart, 1964: 350; Ferrusquía-Villafranca, 1990: mapa; Brown et al., 1998: 32.
Región de las Tierras Bajas Subhúmedas de Tamaulipas West, 1964: 368.
Distrito Chihuahuano (en parte) Cabrera y Willink, 1973: 35.
Provincia de la Planicie Costera del Noreste Rzedowski, 1978: 106; Rzedowski y Reyna-Trujillo, 1990: mapa. Provincia Tamaulipeca Casas-Andreu y ReynaTrujillo, 1990: mapa.
Ecorregión del Mezquital Oriental de México Dinerstein et al., 1995: 110.
Provincia de la Costa Este, N Escalante et al., 1998: 284.
Ecorregión de las Grandes Planicies, Planicie Semárida de Tamaulipas-Texas Challenger y Soberón, 2008: 103.
Esta provincia se ubica en la porción septentrional de la Planicie Costera Nororiental de México, al norte de la cuenca del río Pánuco, en los estados de Tamaulipas y Nuevo León, pequeñas porciones de Coahuila y San Luis Potosí, el norte de Veracruz y la porción adyacente del estado de Texas en los EUA (Morrone, 2001, 2006). La Planicie Costera Nororiental ocupa las tierras bajas al sur del río Bravo y limita hacia el oeste con la Sierra Madre Oriental.
La vegetación consiste de matorrales xerófilos y bosques espinosos, con agaves y cactos abundantes (Dinerstein et al., 1995; Rzedowski, 1978). Entre los géneros de plantas predominantes se encuentran Acacia, Aloysia, Celtis, Condalia, Prosopis y Ziziphus (Katinas et al., 2004).
Algunos taxones endémicos de esta provincia son la biznaga (Coryphantha clavata subsp. stipitata), 2 mezquites (Prosopis reptans var. cinerascens y P. tamaulipana), el guayacón de nadadores (Gambusia marshi), el gecko de bandas del desierto (Coleonyx brevis) y 2 lagartijas (Sceloporus serrifer cyanogenis y S. variabilis marmoratus).
Evolución biótica. Las afinidades principales de esta provincia se dan con las restantes provincias de la región Neártica, en particular con la del Desierto Chihuahuense. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia de Tamaulipas se relaciona estrechamente con las provincias de la Sierra Madre Occidental y Sonora, así como con el norte de la provincia del Desierto Chihuahuense.
Conservación. La provincia de Tamaulipas se halla amenazada por los ranchos ganaderos y el desarrollo industrial a lo largo de la frontera entre México y los EUA (Dinerstein et al., 1995).
Zona de Transición Mexicana
Heilprin (1887), Dugès (1902) y Darlington (1957), entre otros, sugirieron la existencia de una zona de transición entre las regiones Neártica y Neotropical en América Central y México. En una extensa serie de publicaciones referidas fundamentalmente a la entomofauna de zonas montañosas, Gonzalo Halffter definió y precisó la idea de la Zona de Transición Mexicana, definiéndola como el área compleja donde se superponen las faunas neotropical y neártica (Halffter, 1987, 2017; Halffter y Morrone, 2017; Halffter et al., 2008; Llorente, 1996; Zunino y Halffter, 1988). Esta zona no solo destaca por la mezcla de taxones neárticos y neotropicales, sino también por la evolución in situ de numerosas especies, lo cual la distingue de otras zonas de transición, como la del Sahara entre las regiones Paleártica y Afrotropical, en que predomina la ausencia de especies endémicas.
En su sentido más amplio la Zona de Transición Mexicana (Fig. 17) incluye el sudoeste de los EUA de América, México y América Central hasta las tierras bajas de Nicaragua (Halffter, 1987). El área situada al sur del lago Nicaragua, en Costa Rica y Panamá, posee afinidades más marcadas con la biota sudamericana, por lo que no pertenece a la Zona de Transición Mexicana. En el sentido más restringido, la Zona de Transición Mexicana abarca las provincias de la Sierra Madre Occidental, Sierra Madre Oriental, Faja Volcánica Transmexicana, Sierra Madre del Sur y Tierras Altas de Chiapas (Morrone, 2017a). Son estas montañas las que favorecen la dispersión de elementos septentrionales hacia el sur y de elementos meridionales hacia el norte (Zunino y Halffter, 1988). Esta zona de transición equivale a la región Mesoamericana de Montaña (Rzedowski, 1978), al componente Mexicano de Montaña (Morrone y Márquez, 2003) y a la Zona de Transición Mexicana de Montaña (Espinosa-Organista et al., 2008).
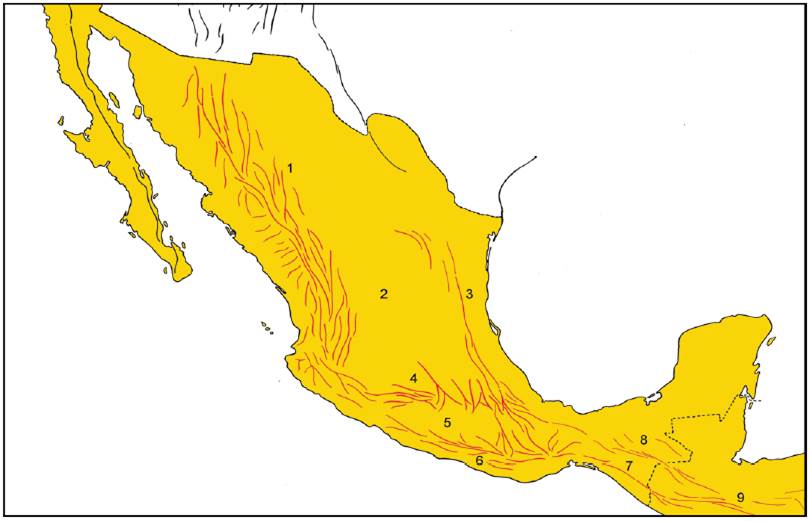
Figura 17 Sistemas orográficos de la Zona de Transición Mexicana, de acuerdo con Halffter (1987). 1, Sierra Madre Occidental; 2, Sierra Madre Oriental; 3, Altiplano Mexicano; 4, Eje Volcánico Transmexicano; 5, Cuenca del Balsas; 6, Sierra Madre del Sur; 7, Sierra Madre de Chiapas; 8, Macizo Central de Chiapas; 9, Núcleo Centroamericano.
Rzedowski (1978) analizó los bosques de la Zona de Transición Mexicana y los comparó con los de latitudes más septentrionales, típicamente neárticos, y latitudes meridionales, típicamente neotropicales. De acuerdo con este autor, si bien las especies dominantes, especialmente los árboles de los géneros Pinus, Abies y Liquidambar, manifiestan relaciones con la región Neártica, la mayoría de las especies de plantas muestran relaciones con taxones de la región Neotropical.
Regionalización biogeográfica
La Zona de Transición Mexicana comprende 5 provincias: Sierra Madre Occidental, Sierra Madre Oriental, Faja Volcánica Transmexicana, Sierra Madre del Sur y Tierras Altas de Chiapas (Morrone, 2017a). Al poseer afinidades neárticas y neotropicales, estas provincias pertenecen simultáneamente a las regiones Neártica y Neotropical.
Evolución biótica
Halffter (1976, 1987, 2003, 2017) analizó la distribución de varios taxones de escarabajos de coleópteros Scarabaeidae de la Zona de Transición Mexicana, teniendo en cuenta también la distribución de sus taxones más afines, la riqueza de especies, su grado de simpatría, la historia geológica del área y la diversidad de hábitats ocupada por las especies (Halffter et al., 1995; Halffter y Morrone, 2017; Liebherr, 1994). Como resultado reconoció 5 patrones de distribución o cenocrones (Fig. 18):
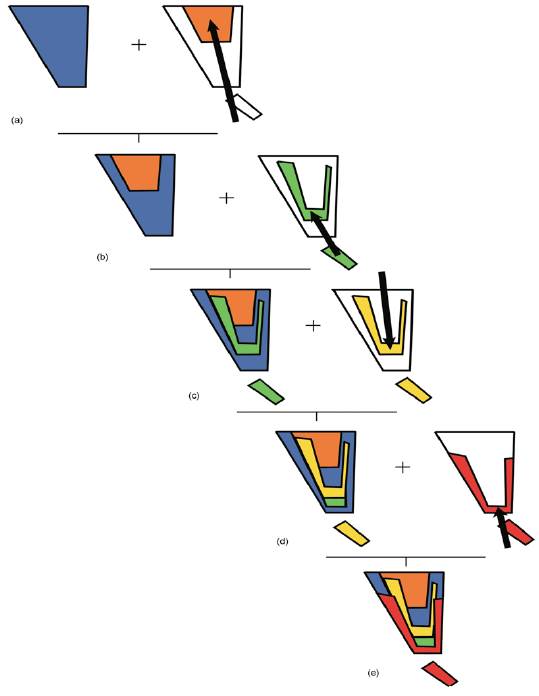
Figura 18 Ensamble de los cenocrones principales de México. (a) Biota Paleoamericana + cenocrón del Altiplano Mexicano; (b) cenocrón Mesoamericano de Montaña; (c) cenocrón Neártico; (d) cenocrón Neotropical Típico; (e) biota actual.
Patrón Paleoamericano. Representa la biota original de la Zona de Transición Mexicana y equivale al elemento Septentrional Antiguo de Savage (1982). Corresponde a taxones que evolucionaron en el Viejo Mundo, en particular de sus zonas tropicales, y que arribaron a América del Norte desde el fin del Cretácico hasta el Paleoceno, desde Eurasia. En general sus especies se distribuyen en el Altiplano Mexicano, la Faja Volcánica Transmexicana y las Sierras Madre; unas pocas especies se encuentran en tierras bajas tropicales. Generalmente pertenecen a géneros de distribución cosmopolita o casi cosmopolita, como Onthophagus (escarabajos) y Bombus (abejorros), cuya mayor riqueza y diversidad se encuentran en el Viejo Mundo.
Patrón del Altiplano Mexicano. Corresponde a taxones de origen neotropical que se dispersaron al Altiplano Mexicano durante el Oligoceno-Mioceno. Constituyen más de 80% de las especies de Scarabaeidae del Desierto Chihuahuense, contrastando su abundancia marcadamente con la de la fauna de vertebrados de esta provincia, que es neártica típica.
Patrón Neártico. Corresponde a taxones de origen norteamericano, ecológicamente asociados con climas templados, que habrían llegado a la Zona de Transición Mexicana en el Plioceno-Pleistoceno. Se distribuyen principalmente en la Sierra Madre Occidental, Sierra Madre Oriental, Sierra Madre del Sur, Faja Volcánica Transmexicana, Tierras Altas de Chiapas y otras áreas montañosas de América Central, generalmente se los encuentra por encima de los 1,700 m. Las especies de este cenocrón se asocian con bosques templados de coníferas y praderas de altura. Las especies de Scarabaeidae de este cenocrón constituyen el 95-100% de las especies en las montañas de la Zona de Transición Mexicana. Un ejemplo es el género Thanatophilus (Silphidae).
Patrón Mesoamericano de Montaña. Corresponde a taxones de bosques montanos que originalmente evolucionaron en América del Sur, luego se dispersaron a América Central, incluyendo las Tierras Altas de Chiapas, Guatemala, Honduras, El Salvador y Nicaragua al norte del lago Nicaragua, y de allí se dispersaron hacia el norte. Sus afinidades filogenéticas son sudamericanas antiguas, y se distribuyen principalmente en bosques de montaña y mesófilos; ocasionalmente penetran en bosques de pinoencino. En el Cretácico Tardío existieron algunas islas continentales en el área centroamericana, así como islas volcánicas entre Nicaragua y Panamá. Al final del Cretácico, el Núcleo Centroamericano se elevó definitivamente, y desde este momento y hasta el Mioceno, la dispersión entre América del Norte y del Sur fue posible para especies “saltadoras de islas”. Desde el fin del Mioceno hasta el Plioceno se elevó el sur de América Central y ambos continentes se unieron. Desde el Plioceno hasta el Reciente los taxones de este cenocrón que evolucionaron en el Núcleo Centroamericano se dispersaron hacia el norte, en las montañas de la Zona de Transición Mexicana. Un ejemplo es la tribu Proculini (Passalidae).
Patrón Neotropical Típico. Corresponde a taxones distribuidos en la región Neotropical, que evolucionaron en América del Sur; equivale al elemento Sudamericano de Savage (1982). Sus especies típicamente se encuentran en bosques tropicales y no se extienden al Altiplano Mexicano por sus condiciones muy secas; a lo largo de las costas del Golfo de México y el océano Pacífico extienden su distribución hacia el norte, pudiendo llegar al sur de los EUA. La dispersión de taxones sudamericanos hacia América del Norte y de taxones norteamericanos hacia Los patrones de distribución de la Zona de Transición América del Sur se conoce como Gran Intercambio Biótico Mexicana muestran una zonación relativamente marcada. Americano (ver Cione et al., 2015). Los elementos del Altiplano Mexicano, neotropicales típicos y mesoamericanos de montaña se encuentran a altitudes menores a los 2,000-3,000 m; entre los 2,000 y 3,000 m hay una fauna que combina elementos neárticos y paleoamericanos; y por encima de los 3,000 m los insectos son casi exclusivamente neárticos (Halffter et al., 1995; Halffter y Morrone, 2017).
Autores trabajando con otros grupos taxonómicos han postulado hipótesis semejantes, que enfatizan el carácter complejo de la biota de la Zona de Transición Mexicana y la antigüedad de algunos de sus elementos (Halffter et al., 2008). Por ejemplo, Bussing (1976, 1985) analizó la fauna de peces de agua dulce de América Central, incluyendo el sureste de México, y distinguió 2 conjuntos ictiofaunísticos. La paleoictiofauna correspondería a elementos neotropicales antiguos, que se habrían dispersado desde América del Sur en el Cretácico-Paleoceno. La neoictiofauna, más reciente, se habría dispersado desde América del Sur en el PlioPleistoceno, con el surgimiento del Istmo de Panamá. Savage (1982) analizó la herpetofauna centroamericana y distinguió 4 elementos: Mesoamericano, Septentrional Antiguo, Sudamericano y Septentrional Reciente. Morrone y Márquez (2001) llevaron a cabo un análisis de trazos de varios taxones de coleópteros de la Zona de Transición Mexicana, hallando un trazo generalizado septentrional, que comprende básicamente áreas montanas de la Sierra Madre Occidental, la Sierra Madre Oriental, la Faja Volcánica Transmexicana, la Cuenca del Balsas y la Sierra Madre del Sur. Un análisis más reciente (Espinosa-Organista et al., 2008) identificó 6 patrones que se asocian con sistemas montañosos particulares (Fig. 19):

Figura 19 Trazos generalizados principales de México obtenidos por Espinosa-Organista et al. (2008). 1, Costero-montano; 2, montano subhúmedo circumbalsas; 3, montano árido y semiárido circumaltiplano; 4, mesófilo montano; 5, montano de la vertiente del Golfo; 6, montano subhúmedo del Pacífico.
Patrón costero-montano. Especies que se distribuyen sobre las vertientes costeras de las Sierras Madre Occidental, Oriental, del Sur y de Chiapas, así como los extremos de la Faja Volcánica Transmexicana. Sustentado por la presencia de Cicindela fera (coleóptero) y Oryzomys melanotis (ratón). Algunas especies se extienden hacia el sur, hasta las cordilleras de América Central.
Patrón montano subhúmedo circumbalsas. Especies que se distribuyen en las laderas de la Faja Volcánica Transmexicana y la Sierra Madre del Sur, predominantemente en las vertientes orientadas hacia la Cuenca del río Balsas. Sustentado por la presencia del helecho Adiantum shepherdii, el pino Pinus pringlei, el maguey Agave cupreata, la lagartija Phrynosoma taurus y el rascador pinto colarejo (Pipilo ocai). Algunas especies se extienden hacia la vertiente seca de la Sierra de Juárez, en Oaxaca.
Patrón montano árido y semiárido circumaltiplano. Varias especies que se distribuyen de forma diferencial sobre las laderas de las cordilleras que rodean el Altiplano Mexicano. Sustentado por la presencia de la lagartija Phrynosoma orbiculare y el rascador pardo (Pipilo fuscus). Dentro de este patrón, muchas especies limitan su distribución en función del sustrato en que crecen, como rocas ígneas o rocas calizas.
Patrón mesófilo montano. Varias especies que se distribuyen discontinuamente sobre la parte más húmeda de la vertiente del Golfo de México. Sustentado por la presencia de un helecho (Cyathea bicrenata), el cucharillo (Clethra suaveolens), una salamandra (Eleutherodactylus buschi) y una lagartija (Sceloporus salvini). La distribución de su flora y fauna abarca desde los bosques mesófilos de montaña del Carso Huasteco hasta los de las Sierras de Guatemala.
Patrón montano de la vertiente del golfo. Especies que se distribuyen ampliamente sobre las montañas de la vertiente del Golfo de México, en ecotonos entre bosques de coníferas y encinos, bosques mesófilos de montaña y bosques húmedos. Sustentado por la presencia del helecho Anemia semihirsuta, el encino Quercus lancifolia, y el ratón Peromyscus furvus. Varias especies extienden su distribución hasta el Petén, en Guatemala y Belice, mientras que otras restringen su distribución a la porción sur del Golfo de México.
Patrón montano subhúmedo del Pacífico. Especies que habitan ecotonos entre bosques de coníferas y encinos y selvas bajas caducifolias de la vertiente del Pacífico. Sustentado por la presencia de los pinos Pinus douglasiana y P. herrerae, el maguey Agave rhodacantha, los encinos Quercus splendens y Q. urbanii, y el coleóptero Cicindela aeneicollis.
Un análisis de parsimonia de endemismos basado en especies de coleópteros de la familia Passalidae (Gutiérrez-Velázquez et al., 2013) llevó a identificar 3 zonas generales dentro de la Zona de Transición Mexicana. Una incluye una parte de la provincia de las Tierras Altas de Chiapas, otra se sitúa en la provincia de la Sierra Madre Oriental y la tercera en la provincia de la Sierra Madre del Sur. De acuerdo con los autores, la fauna de la primera corresponde a los cenocrones Mesoamericano de Montaña y Neotropical Típico, y sus especies se hallan entre los 800 y 3,000 m. La segunda y tercera zonas generales corresponden al cenocrón Mesoamericano de Montaña, distinguiéndose porque sus especies se distribuyen entre los 1,000 y 2,300 m y entre los 2,300 y 2,800 m, respectivamente.
Con el objeto de determinar las relaciones bióticas entre las provincias de la Zona de Transición Mexicana y otras áreas, se han llevado a cabo varios análisis biogeográficos cladísticos. Liebherr (1994) llevó a cabo un análisis basado en varios taxones de coleópteros Carabidae. Obtuvo un cladograma general de áreas en el cual la Sierra Madre Oriental resultó el área hermana de las restantes áreas estudiadas, las cuales constituyen una tricotomía entre el sur de la Sierra Madre Occidental; el resto de la Sierra Madre Occidental unido al Desierto Chihuahuense, el
Desierto de Sonora y las montañas de Arizona; y la Faja Volcánica Transmexicana y la Sierra Madre del Sur unidos a la cordillera de Talamanca y las Tierras Altas de Chiapas. Marshall y Liebherr (2000) llevaron a cabo otro análisis biogeográfico cladístico basado en un número mayor de taxones, que incluyeron géneros o grupos de especies de insectos, peces, saurios y plantas. En el cladograma general de áreas obtenido, estos autores encontraron una dicotomía básica entre un área septentrional (Sierras Madre Occidental y Oriental, junto con los desiertos de Sonora y Arizona, EUA) y otra meridional (Faja Volcánica Transmexicana, Sierra Madre del Sur y el sur de la Sierra Madre Occidental, junto con las Tierras Altas de Chiapas y la cordillera de Talamanca en América Central). De acuerdo con Marshall y Liebherr (2000), en la Zona de Transición Mexicana básicamente existen 2 biotas separadas por la Faja Volcánica Transmexicana.
Otro análisis biogeográfico cladístico de la Zona de Transición Mexicana fue llevado a cabo por Miguez-Gutiérrez et al. (2013). Los autores utilizaron 3 propuestas de regionalización diferentes (Flores-Villela y Goyenechea, 2001; Marshall y Liebherr, 2000; Morrone, 2006), que involucran 9, 13 y 16 áreas de endemismo, respectivamente. El análisis se basó en las filogenias de 10 taxones de coleópteros, lagartijas, serpientes y plantas. Para cada uno de ellos obtuvieron los cladogramas taxonómicos de áreas, remplazando las especies terminales por las áreas de endemismo habitadas por ellas, y luego obtuvieron cladogramas generales de áreas. A partir de los cladogramas generales de áreas obtenidos, los autores procedieron a reconciliarlos con las 3 regionalizaciones analizadas, con el objeto de determinar cuáles presentaban la codivergencia máxima, es decir que minimizaban el número de duplicaciones y pérdidas que era necesario postular. Miguez-Gutiérrez et al. (2013) obtuvieron 30 cladogramas taxonómicos de áreas (10 por cada regionalización) y a partir de ellos se obtuvieron los cladogramas generales de áreas empleando distintos supuestos. El análisis de árboles reconciliados indicó que los cladogramas generales de áreas que reflejaban mejor las relaciones entre las áreas de endemismo son lo resultantes de aplicar el supuesto 0 a las áreas de Marshall y Liebherr (2000). Los 3 cladogramas generales de áreas obtenidos con el supuesto recuperan las mismas relaciones generales: una división neártico-neotropical en la Faja Volcánica Transmexicana; una mayor relación de las Sierra Madre del Sur y la Faja Volcánica Transmexicana, o un grupo incluyendo la Faja Volcánica Transmexicana, la Cuenca del Balsas y la Sierra Madre del Sur; y la Cordillera de Talamanca relacionada con las Tierras Altas de Chiapas y Guatemala. Estos resultados confirman la importancia de la Faja Volcánica Transmexicana y el Istmo de Tehuantepec como eventos vicariantes para numerosos taxones.
Un análisis biogeográfico cladístico más reciente fue llevado a cabo por Corral-Rosas y Morrone (2017). Éste consistió en un análisis con rebanado temporal, para investigar el ensamblaje biótico de la Zona de Transición Mexicana en 3 ventanas temporales: Mioceno, Plioceno y Pleistoceno. Los autores consideraron como unidades de su análisis las 5 provincias de la Zona de Transición Mexicana, así como las regiones Neártica y Neotropical en sentido estricto. A partir de los cladogramas de 49 taxones de animales y plantas, construyeron los cladogramas taxonómicos de áreas respectivos, remplazando las especies terminales por las áreas. Asimismo, asignaron los taxones a su cenocrón más probable considerando la distribución de sus grupos hermanos y, cuando existiera, datación molecular. Luego, llevaron a cabo un análisis de parsimonia de subárboles libres de paralogía particionando la matriz de datos en 3 matrices, cada una correspondiente a una rebanada temporal: Mioceno (correspondiente al cenocrón Mesoamericano de Montaña), Plioceno (suma de los cenocrones Mesoamericano de Montaña y Neártico) y Pleistoceno (suma de los cenocrones anteriores más el Neotropical Típico). Como resultado del análisis, Corral-Rosas y Morrone (2017) obtuvieron 4 cladogramas generales de áreas (Fig. 20): 1 para el Mioceno, 1 para el Plioceno y 2 para el Pleistoceno. Algunas de las relaciones son coincidentes, como la de la región Neotropical y las Tierras Altas de Chiapas; entre la Sierra Madre Oriental y la Sierra Madre del Sur; y entre la región Neártica, la Sierra Madre Occidental y la Faja Volcánica Transmexicana. Los resultados del análisis de Corral-Rosas y Morrone (2017) coinciden en parte con otros análisis biogeográficos cladísticos, aunque el analizar separadamente las rebanadas temporales permitió esclarecer mejor algunos aspectos. Por ejemplo, la Faja Volcánica Transmexicana posee un papel diferente en el tiempo, comportándose como una ruta de dispersión norte-sur para el cenocrón Neártico, como una ruta de dispersión sur-norte para el cenocrón Mesoamericano de Montaña y como un evento vicariante para el cenocrón Neotropical Típico.
Figura 20 Cladogramas generales de áreas obtenidos por Corral-Rosas y Morrone (2017). (a) Mioceno; (b), Plioceno; (c) Pleistoceno.
Provincia de la Sierra Madre Occidental
Provincia de la Sierra Madre Occidental Goldman y Moore, 1945: 353; Moore, 1945: 218; Stuart, 1964: 350; Rzedowski, 1978: 102; Casas-Andreu y Reyna-Trujillo, 1990: mapa; Ramírez-Pulido y Castro-Campillo, 1990: mapa; Rzedowski y Reyna-Trujillo, 1990: mapa; Arriaga et al., 1997: 64; Morrone, 2001: 34, 2005: 234; Espinosa-Organista et al., 2008: 56; Morrone, 2014a: 27, 2017a: 61; Morrone et al., 2017: 278.
Región de la Sierra Madre Occidental West, 1964: 368.
Provincia Sierra Madre-Occidentalense Ferrusquía-Villafranca, 1990: mapa.
Ecorregión de los Bosques de Pino y Roble de la Sierra Madre Occidental Dinerstein et al., 1995: 102.
Provincia Madreana (en parte) Brown et al., 1998: 30.
Área de la Sierra Madre Occidental Katinas et al., 2004: 166.
Esta provincia se ubica en el oeste de México, en los estados de Sonora, Chihuahua, Sinaloa, Durango, Zacatecas, Nayarit y el norte de Jalisco, entre los 200 y 3,000 m de altitud, con la mayor parte de su superficie situada por encima de los 2,000 m (Morrone, 2017a). Ocupa el sistema montañoso de la Sierra Madre Occidental, que es el más largo y continuo de los sistemas orográficos mexicanos, y corre desde la costa del océano Pacífico, al sur de la frontera con los Estados Unidos, hasta la altura de Nayarit y Jalisco, conectando las Montañas Rocallosas con la Faja Volcánica Transmexicana. La Sierra Madre Occidental posee unos 1,400 km de largo y en algunos sitios supera los 200 km de ancho.
En esta provincia predominan los bosques de pino y de pino-encino (Rzedowski, 1978; Dinerstein et al., 1995). Entre los géneros de plantas dominantes se encuentran Abies, Picea, Pinus, Pionocarpus, Pippenalia, Quercus, Stenocarpha y Trichocoryne (Rzedowski, 1978).
Algunos taxones endémicos de esta provincia son un escarabajo (Onthophagus brevifrons), una serpiente de cascabel (Crotalus willardi), el carpintero de Arizona (Picoides arizonae), el mirlo azteca (Ridwayia pinicola), la ardilla voladora sureña (Glaucomys volans madrensis) y la ardilla de Nayarit (Sciurus nayaritensis).
Regionalización biogeográfica. Dentro de esta provincia se han reconocido 2 distritos (Morrone, 2014a, 2017a): Apache y Durangueño.
Distrito Apache
Provincia Apache Smith, 1941: 109 Distrito Tarahumara Moore, 1945: 218.
Provincia de la Sierra Madre Occidental, N Escalante et al., 1998: 284.
Área de la Sierra Madre Occidental-Altiplano Central (en parte) Marshall y Liebherr, 2000: 205.
Distrito Apache Morrone, 2014a: 29.
Abarca la parte septentrional de la provincia, en los estados de Chihuahua, Sonora y parte de Sinaloa. Sustentado por la presencia de 1 helecho (Cheilanthes arizonica), 2 enebros (Juniperus deppeana var. pachycephala y J. scopolorum) y 2 encinos (Quercus tarahumara y Q. toumeyi).
Distrito Durangueño
Provincia Durangueña Smith, 1941: 109.
Distrito Tepehuano Moore, 1945: 218.
Provincia de la Sierra Madre Occidental, S Escalante et al., 1998: 284.
Área de la Sierra Madre Occidental-Altiplano Central (en parte) Marshall y Liebherr, 2000: 205.
Distrito Durangueño Morrone, 2014a: 29.
Abarca la parte meridional de la provincia, en los estados de Jalisco, Nayarit, Sonora, Zacatecas y partes de Sinaloa y Chihuahua. Sustentado por la presencia de 2 especies de encino (Quercus radiata y Q. undata).
Si bien la regionalización biogeográfica de esta provincia ha reconocido un distrito septentrional y otro meridional, un análisis basado en especies de aves (Kobelkowsky-Vidrio et al., 2014) halló una división diferente. Estos autores llevaron a cabo un análisis de parsimonia de endemismos y un análisis de clusters basados en especies de aves. Los autores analizaron los datos de distribución de 148 especies de aves terrestres y residentes en el área (es decir que excluyeron las migratorias). Para estas especies obtuvieron los modelos de distribución potencial y los superpusieron, con el objeto de identificar áreas de mayor riqueza. Asimismo, crearon 2 matrices de presenciaausencia en celdas hexagonales de tamaños diferentes y las analizaron con algoritmos de parsimonia y de análisis de clusters. Como resultado de la superposición de los modelos de distribución, Kobelkowsky-Vidrio et al. (2014) determinaron que la vertiente occidental de la Sierra Madre Occidental posee mayor riqueza específica, especialmente por debajo de los 1,700 m de altitud. De acuerdo con el cladograma obtenido en el análisis de parsimonia de endemismos con celdas de mayor tamaño, existe un agrupamiento de la mayor parte de las áreas, dejando por fuera las del noreste. En el análisis con celdas de menor tamaño se observan 2 clados principales, uno en el sudoeste y otro en el norte-noreste. En el análisis de clusters se obtuvieron 2 grupos principales: uno en la vertiente pacífica y otro en el norte. Los autores concluyeron que la influencia combinada de la elevación y la temperatura era la que tenía mayor influencia en los patrones de riqueza específica. Los análisis de parsimonia de endemismos y de clusters permitieron separar un área sudoeste (vertiente pacífica) de un área noreste (vertiente oriental), las cuales coincidirían con un cambio en la vegetación: tropical en el área sudoeste, templada en la porción oriental interna y xérica en el norte y extremo este. Las aves del oeste tendrían preferencia por bosques deciduos tropicales, mientras que las del noreste preferirían bosques templados en las partes altas, matorral xerófilo en el desierto de Sonora y pastizales templados en el desierto de Chihuahua. Por otra parte, las afinidades de las aves de ambas áreas son diferentes: las del oeste son principalmente mesoamericanas (es decir, neotropicales) y las del este poseen afinidad neártica.
Evolución biótica. Las afinidades bióticas principales de esta provincia se dan con otras provincias de la Zona de Transición Mexicana, como la Faja Volcánica Transmexicana, la Sierra Madre Oriental y la Sierra Madre del Sur, así como con otras provincias neárticas y neotropicales. En un análisis de biogeografía cuantitativa basado en especies de aves, Hernández-Baños et al. (1995) hallaron similitudes entre esta provincia y la de la Faja Volcánica Transmexicana. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia de la Sierra Madre Occidental se relaciona estrechamente con las provincias de Sonora y de Tamaulipas, así como con la porción norte de la provincia del Desierto Chihuahuense. Un análisis de trazos basado en especies de coleópteros (Morrone y Márquez, 2001) mostró que esta provincia está relacionada con las provincias de la Cuenca del Balsas, Sierra Madre del Sur, Sierra Madre Oriental y Faja Volcánica Transmexicana.
Conservación. La provincia de la Sierra Madre Occidental se halla amenazada por la extracción forestal comercial, la conversión de tierras para la agricultura y ganadería, el sobrepastoreo y los cultivos para la producción de drogas ilegales (Dinerstein et al., 1995). La avifauna asociada con los bosques templados de la Sierra Madre Occidental está amenazada por la deforestación intensiva, con algunas aves señaladas por su estado crítico (Kovelkowsky-Vidrio et al., 2014), como el carpintero imperial (Campophilus imperialis), el quetzal orejón (Euptilotis neoxenus) y la chara pinta (Cyanocorax dickeyi). En esta provincia se incluyen varias áreas naturales protegidas (Conanp, 2018):
Parque Nacional Cascada de Bassaseachic (Chihuahua). Incluye ecosistemas diversos que van desde el bosque de coníferas hasta la selva baja y en su centro la Barranca de Candameña, por la que desemboca la cascada de Bassaseachic, suministro de agua permanente para los seres vivos del lugar. Especies representativas son el pino real (Pinus durangensis), 3 especies de encinos (Quercus sideroxila, Q. rugosa y Q. arizonica), el venado cola blanca (Odocoileus virginianus) y la nutria (Lontra longicaudis annectens).
Parque Nacional Sierra de Órganos (Zacatecas). Destaca la composición y estructura de la vegetación por gradiente altitudinal, con densidades grandes de rodales con pino piñonero (Pinus cembroides), alamillo tamblón (Populus tremuloides) y encino rojo (Quercus aduardii). Algunas especies representativas son la rata canguro de Phillips (Dipodomys phillipsii), la matraca serrana (Campylorhynchus gularis), el zorzal mexicano (Catharus occidentalis), la cardellina rubra (Ergaticus ruber), la calandria de flancos negros (Icterus abeillei), el mirlo azteca (Ridgwayia pinicola), el gorrión de Worthen (Spizella wortheni), el pino piñonero (Pinus cembroides), la biznaga (Mammillaria moelleriana), el gatuño (Mimosa laxiflora), el coyote (Canis latrans), el lince (Lynx rufus), el halcón peregrino (Falco peregrinus), la zorra gris (Urocyon cinereoargenteus) y el cacomixtle (Bassariscus sumichrasti).
Provincia de la Sierra Madre Oriental
Provincia de la Sierra Madre Oriental Goldman y Moore, 1945: 356; Moore, 1945: 218; Stuart, 1964: 350; Rzedowski, 1978: 103; Casas-Andreu y Reyna-Trujillo, 1990: mapa; Ramírez-Pulido y Castro-Campillo, 1990: mapa; Rzedowski y Reyna-Trujillo, 1990: mapa; Arriaga et al., 1997: 64; Morrone, 2001: 35, 2001: 34, 2005: 235; Espinosa-Organista et al., 2008: 56; Morrone, 2014a: 29, 2017a: 65; Morrone et al., 2017: 278.
Región de la Sierra Madre Oriental West, 1964: 368.
Provincia Sierra-Madre-Orientalense Ferrusquía-Villafranca, 1990: mapa.
Ecorregión de los Bosques de Pino y Roble de la Sierra Madre Oriental Dinerstein et al., 1995: 102.
Provincia Madreana (en parte) Brown et al., 1998: 30.
Área de la Sierra Madre Orienta Marshall y Liebherr, 2000: 206.
Esta provincia se ubica en el este de México, en los estados de Coahuila, Durango, Nuevo León, Zacatecas, San Luis Potosí, Tamaulipas, Guanajuato, Querétaro, Hidalgo, Veracruz y Puebla, a una altitud superior a los 1,500 m (Morrone, 2017a). Corresponde al sistema montañoso de la Sierra Madre Oriental, el segundo en extensión del país, que se extiende desde el centro de Nuevo León en el norte hasta Puebla y Veracruz en el sur, donde se une con la Faja Volcánica Transmexicana. Comprende un conjunto de serranías, por lo general con recorrido norte-sur, donde dominan rocas cretácicas. La ladera occidental es seca, mientras que la oriental, dirigida hacia el Golfo de México, es húmeda. Esto, aunado a la existencia de valles alargados, facilita la existencia de elementos tropicales a elevaciones relativamente altas y de elementos montanos a altitudes menores. El clima es muy variado, presentándose el 99% de los climas del país (Hernández-Cerda y Carrasco-Anaya, 2004).
Predominan los bosques de encino, aunque también hay bosques mesófilos, bosques de pino y otras comunidades vegetales (Rzedowski, 1978; Dinerstein et al., 1995). En la vertiente oriental (húmeda) predominan los bosques templados, mientras que en la porción occidental, en el límite con el Altiplano (provincia del Desierto Chihuahuense), también se encuentran áreas con matorral xerófilo (Sanginés-Franco et al., 2011). Entre los géneros predominantes de plantas se destacan: Carpinus, Cornus, Fagus, Greenmaniella, Hamamelis, Loxothysanus, Mathiasella, Pinus, Platanus y Quercus (Rzedoswki, 1978; Katinas et al., 2004).
Algunos taxones endémicos de esta provincia son el nogal encarcelado (Juglans mollis), el maguey de la Sierra Madre Oriental (Agave tenuifolia), una mariposa (Priamides erostratus erostratinus), una salamandra (Eleutherodactylus longipes), el ratón ocotero (Peromyscus aztecus), la musaraña de la Sierra Madre Oriental (Cryptotis obscura) y el mochuelo tamaulipeco (Glaucidium sanchezi). La provincia alberga varias especies endémicas de gimnospermas, como Ceratozamia hildae, C. sabatoi, Picea martinezi, Pinus culminicola y Zamia fisheri (Contreras-Medina, 2004). En cuanto a las plantas vasculares, Salinas-Rodríguez et al. (2017) detectaron 1,135 especies, siendo las familias Asteraceae (232 especies), Cactaceae (170), Fabaceae (66) y Lamiaceae (66) las más diversas. La endemicidad de anfibios y reptiles es muy alta, con 43 especies restringidas a la Sierra Madre Occidental (Canseco-Márquez et al., 2004).
Regionalización biogeográfica. Dentro de esta provincia se han reconocido 2 subprovincias y 4 distritos (Espinosa et al., 2004; Espinosa-Organista et al., 2008; Morrone, 2014a, 2017a). Las subprovincias y distritos están definidos por especies endémicas de angiospermas, gimnospermas, mariposas y ratones (Espinosa et al., 2004).
Subprovincia Austro-Oriental
Provincia Austro-Oriental Smith, 1941: 108.
Provincia de la Sierra Madre Oriental N Escalante et al., 1998: 284.
Subprovincia Meridional Espinosa et al., 2004: 294. Subprovincia Austro-Oriental Morrone, 2014a: 30.
Corresponde a la porción septentrional, al norte del río Moctezuma. Posee 2 distritos: Saltillo-Parras y Potosí. Distrito Saltillo-Parras
Distrito Saltillo-Parras Espinosa et al., 2004: 294; Morrone, 2014a: 31.
Distrito de las Sierras Transversales Espinosa-Organista et al., 2008: 56.
Corresponde al Pliegue Saltillo-Parras, las Sierras Transversales y parte de la Gran Sierra Plegada. Está sustentado por la presencia de 2 especies de lagartijas (Sceloporus parvus y Xenosaurus newmanorum).
Distrito Potosí
Distrito Potosí Espinosa et al., 2004: 294; Morrone, 2014a: 30.
Distrito de la Gran Sierra Plegada Espinosa-Organista et al., 2008: 56.
Corresponde a parte de la Gran Sierra Plegada y las Sierras y Llanuras Occidentales en la Sierra de Potosí. Está sustentado por la presencia de 2 serpientes de cascabel (Crotalus lepidus castaneus y C. lepidus morulus), 1 lagartija (Phrynosoma orbiculare orientale), la culebra listonada de montaña (Thamnophis exsul) y la culebra de agua listonada tamaulipeca (T. mendax).
Subprovincia Hidalguense
Provincia Hidalguense Smith, 1941: 108.
Provincia de la Sierra Madre Oriental S Escalante et al., 1998: 284.
Subprovincia Septentrional Espinosa et al., 2004: 294. Subprovincia Hidalguense Morrone, 2014a: 30.
Se sitúa al sur del río Moctezuma. Comprende 2 distritos: Sierra Gorda y Zacualtipán.
Distrito de la Sierra Gorda
Distrito de la Sierra Gorda Espinosa et al., 2004: 294; Morrone, 2014a: 30.
Delimitado por los ríos Moctezuma y Verde. Sustentado por la presencia de una especie de maguey (Agave tenuifolia), el shishote (Pseudosmodingium virletii) y el xenosaurio de cabeza plana (Xenosaurus platyceps).
Distrito Zacualtipán
Distrito Zacualtipán Espinosa et al., 2004: 294; Morrone, 2014a: 30.
Situado hacia el oriente, en la Sierra Norte de Puebla y la Sierra de Zacualtipán. Sustentado por la presencia del piñón enano del Potosí (Pinus culminicola), 1 encino (Quercus sinuata var. breviloba) y la rata montera (Neotoma mexicana navus).
Evolución biótica. Esta provincia es claramente transicional, mostrando afinidades con provincias neárticas (Desierto Chihuahuense) y neotropicales (Veracruzana y Cuenca del Balsas), si bien las afinidades más evidentes son con las otras provincias transicionales, como la Sierra Madre Occidental, la Faja Volcánica Transmexicana, la Sierra Madre del Sur y los Altos de Chiapas. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia de la Sierra Madre Oriental se relaciona estrechamente con la porción sur de la provincia del Desierto Chihuahuense, mientras que otro análisis de parsimonia de endemismos basado en coleópteros (Márquez y Morrone, 2004) la asocian con las provincias Veracruzana y de Los Altos de Chiapas. Un análisis de trazos basado en especies de coleópteros (Morrone y Márquez, 2001) mostró que esta provincia se relaciona con las provincias de la Sierra Madre Occidental, Sierra Madre del Sur, Cuenca del Balsas y Faja Volcánica Transmexicana. Otro análisis de trazos, basado en helechos (Sanginés-Franco et al., 2011), destacó las complejas relaciones bióticas de esta provincia, en particular de su parte más septentrional. Algunos análisis biogeográficos (Sánchez-González et al., 2008; Arbeláez-Cortés et al., 2010) han sugerido que la provincia de la Sierra Madre Oriental podría ser una unidad compuesta. La porción norte se relacionaría más estrechamente con la Sierra Madre Occidental, y la sur con la Sierra Madre del Sur y la Faja Volcánica Transmexicana.
Ferro et al. (2017) analizaron la distribución geográfica de las aves de la Sierra Madre Oriental para localizar con precisión el área de transición entre las regiones Neártica y Neotropical. Para ello, evaluaron las afinidades ecológicas (afinidad corológica) e históricas (afinidad biogeográfica evolutiva). El área de estudio abarcó la Sierra Madre Oriental, a la que se le sumaron áreas aledañas de las provincias del Desierto Chihuahuense y Veracruzana. Se analizaron 1,517 localidades y 264 especies. Se construyó una matriz de presencia-ausencia con celdas de 1° × 1° y otra con celdas de 0.5° × 0.5°. En la primera parte del análisis se identificaron corotipos como patrones resultantes de la superposición significativa de las distribuciones de distintas especies. En la segunda se identificaron cenocrones, de acuerdo con la información filogenética y la distribución geográfica de los taxones afines. Los gradientes de afinidad se representaron en mapas y se calcularon índices de diversidad. Ferro et al. (2017) hallaron 11 corotipos, que abarcan 89% de las especies analizadas. Estos corotipos corresponden a diferentes afinidades corológicas: (1) región Neártica; (2) oeste de América del Norte; (3) región Neotropical; (4) planicies costeras del Golfo de México; (5) Sierras Madre y Faja Volcánica Transmexicana; (6) igual que el anterior, pero extendiéndose hasta América del Sur; (7) tierras bajas de América Central; (8) costa atlántica de América Central y costa pacífica de América del Sur; (9) Sierra Madre Oriental, Sierra Madre del Sur y Sierra Madre de Chiapas; (10) tierras bajas del este de América Central y costa Pacífica de América del Sur hasta Ecuador; y (11) tierras bajas de ambos océanos de México y América Central. En cuanto a los cenocrones, los autores identificaron 4: (1) cenocrón Oligoceno de diversificación holártica y ancestría gondwánica; (2) cenocrón Eoceno de diversificación neotropical y ancestría gondwánica; (3) cenocrón Mioceno de diversificación neotropical y ancestría holártica; y (4) cenocrón Plio-Pleistoceno de diversificación neotropical y ancestría neártica. Al analizar su frecuencia en las distintas celdas, los 3 primeros cenocrones resultaron más frecuentes a mayores altitudes, mientras que el cuarto lo fue en zonas bajas. Los índices de diversidad indicaron que las áreas de mayor mezcla de corotipos se encuentran a altitudes intermedias, entre los 1,200-1,500 m y en la vertiente oriental. Por otra parte, la mezcla de cenocrones es mayor entre 1,500-2,000 m, en el centro y sur de la Sierra Madre Oriental. Si bien las transiciones halladas son relativamente graduales, es posible hallar una transición más marcada que se extiende desde los bosques mesófilos del Carso Huasteco hasta los cañones de la cuenca del río Pánuco y luego hacia el este, siguiendo al Pánuco, hasta su desembocadura en el Golfo de México. Esta transición coincide con una disminución abrupta de la altitud de 4,000 a 2,500 m. La cuenca del Pánuco, a su vez, coincide con el límite de las subprovincias Austro-Oriental e Hidalguense.
Conservación. La provincia de la Sierra Madre Oriental se halla amenazada por la extracción forestal, la construcción de carreteras, el sobrepastoreo y las presiones de asentamientos humanos (Dinerstein et al., 1995). El análisis de trazos de especies de helechos de la Sierra Madre Oriental de Sanginés-Franco et al. (2011) permitió identificar 5 nodos (Nuevo León, Tamaulipas, Landa, Sierra Gorda y Orizaba), que coinciden con áreas geológicamente complejas y serían áreas particularmente relevantes para la conservación. Los nodos Sierra Gorda y Landa ya han sido identificados por el gobierno mexicano como reservas de la biosfera. En esta provincia se incluyen varias áreas naturales protegidas (Conanp, 2018):
Reserva de la Biosfera Sierra Gorda (Querétaro). Posee gran diversidad altitudinal, de 200 a 3,300 m, derivando en una complejidad fisiográfica con cañones, cumbres, abruptas sierras, valles intermontanos y dolinas. Incluye bosques de coníferas, bosques de encino, bosques mesófilos de montaña, matorral xerófilo, selva caducifolia, selva perennifolia y vegetación inducida.
Reserva de la Biosfera Sierra Gorda de Guanajuato (Guanajuato). Dominada por paisajes volcánicos, montañosos sedimentarios hasta paisajes cársticos. Posee vegetación acuática y subacuática, bosques de coníferas, bosques de encino, bosques mesófilos de montaña, pastizal inducido, matorral xerófilo y bosques tropicales caducifolios.
Parque Nacional Los Mármoles (Hidalgo). Bosques de encino, vegetación inducida y bosques de pino. Algunas especies representativas son las dalias (Dahlia sp.), el pino ocote (Pinus pseudostrobus), el cedro (Cupressus lusitanica), la palma de todos los santos (Ceratozamia mexicana), el ahuimo bermellón (Beschorneria tubiflora), la palmilla enana azul (Brahea moorei), el laurel mexicano (Litsea glaucescens), la salamandra de cueva (Chiropterotriton mosaueri), la rana de cascada (Lithobates pustulosus), el ajolote de tierra (Isthmura bellii), el tlaconete regordete (Aquiloeurycea cephalica), la serpiente de cascabel oscura de Querétaro (Crotalus aquilus), la serpiente cascabel cola negra (Crotalus molossus), el eslizón encinero (Plestiodon lynxe), la lagartija nocturna de Gaiger (Lepidophyma gaigeae), la culebra chata de Baird (Salvadora bairdi), el cincuate mexicano (Pituophis deppei), el zorzal corona negra (Catharus mexicanus) y la mariposa monarca (Danaus plexippus).
Reserva de la Biosfera Sierra del Abra Tanchipa (San Luis Potosí). Se sitúa en la parte media-este de la Sierra Madre Oriental y constituye el límite boreal de los ecosistemas neotropicales con bosques caducifolios de la vertiente del Golfo de México. Posee altos niveles de diversidad estimados de flora y fauna. Algunas especies representativas son el oyate pata de elefante (Beaucarnea recurvata), el chamal (Dioon edule), el mocoque (Pseudobombax ellipticum), el ébano (Ebenopsis ebano), la ceiba (Ceiba pentandra), el jaguar (Panthera onca), el tigrillo (Leopardus wiedii), el ocelote (Leopardus pardalis), la guacamaya verde (Ara militaris) y el loro cabeza tamaulipeco (Amazona viridigenalis).
Parque Nacional Cumbres de Monterrey (Nuevo León). Bosques de encino, de pino, de pino-encino y, de ayarín, matorral submontano, matorral desértico rosetófilo y matorral desértico micrófilo. Especies representativas incluyen el pinabete (Pseudotsuga menziesii), el maguey (Agave montana), el pino blanco (Pinus pseudostrobus), el palmito (Brahea berlandieri), el oso negro (Ursus americanus), la cotorra serrana oriental (Rhynchopsitta terrisi), el escorpión pigmeo (Gerrhonotus parvus), la salamandra pie plano primitiva (Chiropterotriton priscus) y la serpiente de cascabel tropical (Crotalus totonacus).
Área Natural Protegida Altas Cumbres (Tamaulipas). Área contigua a Ciudad Victoria; posee diferentes tipos de vegetación, como selva baja caducifolia, bosque de galería, bosque mesófilo de montaña y bosque de pino-encino. Posee una gran riqueza florística, con varias especies de plantas endémicas (García-Morales et al., 2014).
Provincia de la Faja Volcánica Transmexicana Provincia Austro-Central Smith, 1941: 108.
Provincia Volcánica Transversa Goldman y Moore, 1945: 356; Moore, 1945: 218; Stuart, 1964: 350; Ramírez-Pulido y Castro-Campillo, 1990: mapa.
Provincia de las Serranías Meridionales (en parte) Rzedowski, 1978: 103; Rzedowski y Reyna-Trujillo, 1990: mapa.
Provincia del Eje Neovolcánico Casas-Andreu y Reyna-Trujillo, 1990: mapa; Rzedowski y Reyna-Trujillo, 1990: mapa; Arriaga et al., 1997: 64; Espinosa-Organista et al., 2008: 56.
Provincia Neovolcanense Ferrusquía-Villafranca, 1990: mapa.
Ecorregión de los Bosques de Pino y Roble Transvolcánicos de México Dinerstein et al., 1995: 102.
Ecorregión de la Tundra Alpina de México Dinerstein et al., 1995: 107.
Provincia del Eje Volcánico Arriaga et al., 1997: 64.
Provincia Transvolcánica Brown et al., 1998: 29.
Provincia de la Faja Volcánica Escalante et al., 1998: 284.
Área del Eje Transvolcánico Flores-Villela y Goyenechea, 2001: 174.
Provincia del Eje Volcánico Transmexicano Morrone, 2001: 36, 2005: 236.
Área Volcánica Transmexicana Katinas et al., 2004: 166.
Área Transvolcánica Flores-Villela y Martínez-Salazar, 2009: 820.
Provincia de la Faja Volcánica Transmexicana Morrone, 2014a: 31, 2017a: 69; Morrone et al., 2017: 278.
Esta provincia se ubica en el centro de México, en los estados de Aguascalientes, Guanajuato, México, Ciudad de México, Jalisco, Michoacán, Nayarit, Puebla, Oaxaca, Tlaxcala y Veracruz, por encima de los 1,800 m de altitud (Morrone, 2017a). Corresponde al sistema montañoso de la Faja Volcánica Transmexicana, conocido también como Eje Neovolcánico Transversal, que es un conjunto de cordilleras de volcanes que cruza el país de oeste a este, desde Cabo Corrientes (Nayarit) hasta la Sierra Chinconquiaco (Veracruz), aproximadamente entre los paralelos 19 y 21º de latitud norte (Gámez et al., 2012). Entre los volcanes de la Faja Volcánica Transmexicana destacan el Ceborruco (Nayarit), Nevado de Colima (Jalisco y Michoacán), Pico de Tancítaro (Michoacán), Nevado de Toluca (Estado de México) e Iztaccíhuatl y Popocatépetl (Estado de México y Puebla). Constituye el límite austral del Desierto Chihuahuense y lo separa de la Cuenca del Balsas. La Faja Volcánica Transmexicana comenzó a desarrollarse durante el Mioceno en el centro de México y luego se expandió hacia el este y oeste, culminando su configuración actual en el Holoceno. La faja de volcanes se desarrolló durante 19 ma, a lo largo de 4 episodios de vulcanismo que afectaron el área de manera asincrónica, primero en el oeste y luego en el este. La historia geológica de la Faja Volcánica Transmexicana y sus variadas conexiones bióticas con otras provincias biogeográficas la hacen una de las provincias más complejas y heterogéneas del país (Gámez et al., 2012). Su posición es privilegiada, ya que conecta entre sí las Sierras Madre Occidental, Oriental y del Sur, representando el núcleo de la Zona de Transición Mexicana (Espinosa y Ocegueda, 2007; Halffter y Morrone, 2017).
La vegetación de esta provincia es muy variada. Predominan los bosques de pino-encino; también hay bosques tropicales caducifolios, así como matorrales xerófilos en áreas con derrames lávicos y zacatonales (tundra alpina) cerca de la cima de los grandes volcanes (Dinerstein et al., 1995; Rzedowski, 1978). Entre los géneros dominantes de plantas, destacan: Achillea, Alchemilla, Arenaria, Cerastium, Geranium, Hintonella,
Microspermum, Omiltemia, Peyritschia, Pinus, Quercus, Ranunculus y Silviella (Rzedoswki, 1978; Katinas et al., 2004).
Algunos taxones endémicos de esta provincia son: un helecho (Cheilanthes decomposita), la tronadora (Montanoa frutescens), 2 magueyes (Agave horrida y A. inaequidens), un escarabajo (Onthophagus hippopotamus), la tuza de nariz pelona (Cratogeomys tylorhinus), el teporingo o conejo de los volcanes (Romerolagus diazi) y la musaraña colilarga de Orizaba (Sorex oryzabae). De acuerdo con Alcántara y Paniagua (2007), la Faja Volcánica Transmexicana posee 63 especies de plantas vasculares endémicas, la mayor parte de las cuales se distribuyen en sitios con altitudes intermedias (2,100-3,000 m).
Regionalización biogeográfica. Dentro de esta provincia se han reconocido 2 subprovincias y 4 distritos (Escalante et al., 2007; Gámez et al., 2012; Morrone, 2014a, 2017a; Torres-Miranda y Luna, 2007). La distinción entre ambas subprovincias es marcada (Gámez et al., 2012). Cada subprovincia posee especies endémicas propias y se han identificado diferencias en el clima: templado subhúmedo y semifrío subhúmedo (subprovincia Este) y semicálido subhúmedo (subprovincia Oeste); orogenia: MiocenoPlioceno (subprovincia Este) y Pleistoceno-Cuaternario (subprovincia Oeste); altitud: 2,000-3,000 m (subprovincia Este) y menores de 2,000 m (subprovincia Oeste); y en la vegetación: bosques de pino-encino y matorrales xerófitos (subprovincia Este) y bosques tropicales caducifolios (subprovincia Oeste).
Subprovincia Oeste
Subprovincia Meridional (en parte) Ferrusquía-Villafranca, 1990: mapa.
Subprovincia Septentrional (en parte) Ferrusquía-Villafranca, 1990: mapa.
Distrito Oeste Escalante et al., 2007: 496.
Zona Oeste Suárez-Mota et al., 2013: 101.
Subprovincia Oeste Morrone, 2014a: 32.
Corresponde al occidente de la provincia. Comprende 2 distritos: Otomí y Tarasco.
Distrito Otomí
Distrito Otomí Moore, 1945: 218; Morrone, 2014a: 33.
En la porción centro-norte, en los estados de Jalisco, Guanajuato, Querétaro, Hidalgo, Michoacán y México.
Distrito Tarasco
Distrito Tarasco Moore, 1945: 218; Morrone, 2014a: 33.
En la porción centro-sur, en los estados de Jalisco, Michoacán y México.
Subprovincia Este
Subprovincia Meridional (en parte) Ferrusquía-Villafranca, 1990: mapa.
Subprovincia Septentrional (en parte) Ferrusquía-Villafranca, 1990: mapa.
Distrito Este Escalante et al., 2007: 496.
Zona Este Suárez-Mota et al., 2013: 101.
Subprovincia Este Morrone, 2014a: 31.
Corresponde al oriente de la provincia. Posee 2 distritos: Azteca y Orizaba-Zempoaltepec.
Distrito Azteca
Distrito Azteca Moore, 1945: 218; Morrone, 2014a: 32.
Distrito Toluca-México-Puebla Miranda y Luna, 2007: 513.
En los estados de México, Hidalgo, Tlaxcala, Puebla, Morelos y Guerrero.
Distrito Orizaba-Zempoaltepec
Distrito Orizaba-Zempoaltepec Moore, 1945: 218; Morrone, 2014a: 32.
Provincia del Valle de Tehuacán-Cuicatlán Rzedowski, 1978: 107; Rzedowski y Reyna-Trujillo, 1990: mapa.
Distrito Árido del Sur de México Cabrera y Willink, 1973: 37.
Provincia Oaxaqueña (en parte) Ferrusquía-Villafranca, 1990: mapa.
Subprovincia Cañadiana Ferrusquía-Villafranca, 1990: mapa.
Subprovincia Mixteco-Zapotecana Ferrusquía-Villafranca, 1990: mapa.
Provincia Oaxaco-Tehuacanense (en parte) Ramírez-Pulido y Castro-Campillo, 1990: mapa. Ecorregión de los Bosques Húmedos de Oaxaca Dinerstein et al., 1995: 91.
Ecorregión de los Bosques Secos de Oaxaca Dinerstein et al., 1995: 99.
Ecorregión del Matorral Xerófilo de Puebla Dinerstein et al., 1995: 110.
Distrito Veracruzano Este Miranda y Luna, 2007: 513.
En el sureste de Puebla, norte de Oaxaca y oeste de Veracruz.
Evolución biótica. La provincia de la Faja Volcánica Transmexicana es la que muestra más claramente la condición transicional, constituyendo el "núcleo" de la Zona de Transición Mexicana (Espinosa y Ocegueda, 2007; Halffter y Morrone, 2017). Muestra relaciones con las restantes provincias de la Zona de Transición Mexicana, así como con provincias neotropicales típicas (Tierras Bajas del Pacífico y Cuenca del Balsas). En un análisis de biogeografía cuantitativa basado en especies de aves, Hernández-Baños et al. (1995) hallaron similitudes entre esta provincia y la de la Sierra Madre Occidental. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia de la Faja Volcánica Transmexicana se relaciona estrechamente con las provincias de las Tierras Bajas del Pacífico, Cuenca del Balsas y Sierra Madre del Sur. Dos análisis de trazos basados en especies de coleópteros (Corona et al., 2007; Morrone y Márquez, 2001) mostraron que esta provincia se relaciona con las provincias de la Sierra Madre Occidental, Sierra Madre del Sur, Sierra Madre Oriental, Tierras Bajas del Pacífico y la Cuenca del Balsas. De acuerdo con un análisis biogeográfico cladístico basado en taxones de insectos, peces, reptiles y plantas (Marshall y Liebherr, 2000), la provincia de la Faja Volcánica Transmexicana se relaciona con la provincia de la Sierra Madre del Sur.
Existen varios análisis filogeográficos que correlacionan el desarrollo de la Faja Volcánica Transmexicana con la diversificación asociada con la diversidad de ambientes. El análisis filogeográfico de la especie Nolina parviflora (Ruiz-Sánchez y Specht, 2013) es uno de ellos. Estos autores evaluaron los eventos históricos que habrían inducido la diversificación de esta especie, revelando la historia ecológica y evolutiva de sus poblaciones a lo largo de la Faja Volcánica Transmexicana. El análisis se basó en material colectado en 28 poblaciones, que representan la distribución completa de la especie. Se extrajo el ADN y se amplificaron y secuenciaron 2 marcadores de cloroplastos. Se realizó un análisis filogenético con inferencia Bayesiana y se calcularon los tiempos de divergencia. También se calcularon parámetros poblacionales y se hizo un análisis filogeográfico. El análisis filogenético permitió identificar 2 clados principales. Uno de ellos incluye los especímenes de Jalisco y Zacatecas, y el otro las poblaciones restantes. La divergencia (Fig. 21) de Nolina de su género hermano se calculó en 20.9 ma (Oligoceno-Mioceno Medio). Dentro de N. parviflora, los eventos de divergencia se dataron entre el Oligoceno y el Plioceno Tardío (13.9 ma). Los clados menores dentro de los 2 mayores habrían divergido entre el Mioceno Medio y el Pleistoceno. El análisis filogeográfico dio por resultado 5 haplotipos en la Faja Volcánica Transmexicana, uno de ellos compartido con la Sierra Madre Oriental. En el este de la Faja Volcánica Transmexicana hay una mayor diversidad de haplotipos. La divergencia entre las poblaciones es alta, indicando una fuerte estructuración geográfica. Existen barreras importantes, separando las poblaciones de Jalisco y Zacatecas, las de Guanajuato y Querétaro de la Sierra Madre Oriental, y las de Oaxaca y la Sierra Madre del Sur. De acuerdo con Ruiz-Sánchez y Specht (2013), las estimaciones de divergencia coinciden con los 2 episodios volcánicos principales, ocurridos hace 21-10 ma y 7.5-3 ma, respectivamente. El clado del este habría diversificado entre el Mioceno Temprano y el Plioceno Medio, y algunos de sus subclados entre el Plioceno y el Pleistoceno. Estos periodos concuerdan con el cuarto episodio de vulcanismo ocurrido en la Faja Volcánica Transmexicana (21-1.8 ma). El desarrollo de la Faja Volcánica Transmexicana jugó un papel fundamental en la diversificación de N. parviflora, limitando el flujo génico entre las poblaciones.

Conservación. La provincia de la Faja Volcánica Transmexicana se halla amenazada por la cría de ovejas, la conversión de hábitats naturales para la agricultura, la recolección de leña, la explotación de la vida silvestre y la urbanización intensiva (Dinerstein et al., 1995). El libro editado por Velázquez y Romero (1999) incluye capítulos que analizan las características abióticas, flora, fauna, producción rural, aprovechamiento de la vida silvestre y planeación regional de la Cuenca de México, en la parte central de esta provincia. En esta provincia se incluyen varias áreas naturales protegidas (Conanp, 2018):
Reserva de la Biosfera Mariposa Monarca (Michoacán). En esta reserva todos los años se completa el ciclo migratorio de millones de mariposas monarca, las cuales hibernan en las formaciones elevadas de la Faja Volcánica Transmexicana, entre los estados de Michoacán y México. Posee bosque de coníferas, bosques de encino y vegetación inducida.
Reserva de la Biosfera Tehuacán-Cuicatlán (Oaxaca y Puebla). Cuenta con poblaciones de cactáceas columnares únicas en el mundo, con especies como el tetecho (Neobuxbaumia tetetzo) y los cardones u órganos (Cephalocereus columna trajani, Neobuxbaumia mezcalaensis, N. macrocephala, Pachycereus grandis y P. weberi, entre otros). La selva baja caducifolia ocupa casi el 38% de su extensión y se encuentra en buen estado de conservación, formando un corredor biológico en el que se ha documentado la presencia de jaguar, puma, tigrillo, lince y jaguarundi.
Parque Nacional Bosencheve (Estado de México y Michoacán). Área boscosa con bosque de coníferas y bosque mesófilo de montaña.
Parque Nacional Cumbres del Ajusco (Ciudad de México). Posee varias especies endémicas, lo que se atribuye a la compleja historia geológica y a las características fisiográficas de la región, que generaron una gran variedad de microhábitats.
Parque Nacional Desierto del Carmen o de Nixcongo (Estado de México). Bosques de coníferas y vegetación inducida.
Parque Nacional Desierto de los Leones (Ciudad de México). Bosques de coníferas y vegetación inducida. Algunas especies representativas son el oyamel (Abies religiosa), el cedrón (Juniperus monticola), el madroño borracho (Comarostaphylis discolor), el cedro blanco (Cupressus lusitanica), la mazorquita (Morchella conica), la serpiente de cascabel transvolcánica (Crotalus triseriatus), el escorpión (Barisia imbricata), la codorniz coluda neovolcánica (Dendrortyx macroura), el lince (Lynx rufus), el coyote (Canis latrans) y el ajolote (Ambystoma altamirani).
Reserva Ecológica del Pedregal de San Ángel de Ciudad Universitaria (Ciudad de México). Se ubica dentro del área urbana de la Ciudad de México, en el campus central de la UNAM. Comprende el área donde se solidificaron los flujos de lava de la erupción del volcán Xitle, que ocurrió hace unos 1,670 años. La vegetación consiste en matorral xerófilo, con abundancia de palo loco (Senecio praecox). La obra de Lot y Cano-Santana (2009) compendia el conocimiento abiótico, florístico, faunístico y ecológico de la reserva.
Parque Nacional Iztaccíhuatl-Popocatéptl (Estado de México, Morelos y Puebla). Posee bosques de coníferas, bosques de encino, pastizales y vegetación inducida. Algunas especies representativas son el pino de las alturas (Pinus hartwegii), el lupino (Lupinus montanus), el zacatonal (Calamagrostis tolucensis), el enebro azul (Juniperus monticola), el cardo santo (Cirsium ehrenbergii), el lince (Lynx rufus escuinapae), el teporingo o zacatuche (Romerolagus diazi), la chara copetona (Cyanocitta stelleri), el carpintero de Strickland (Picoides stricklandi), el escorpión (Barisia imbricata), la lagartija escamosa del mezquite (Sceloporus grammicus), el eslizón de Cope (Eumeces copei) y la culebra listonada del sur mexicano (Thamnophis eques).
Área de Protección de Flora y Fauna Corredor Biológico Chichinautzin (Morelos). Es importante por los servicios ecosistémicos que presta a la Ciudad de México y el estado de Morelos, principalmente de agua para zonas urbanas y rurales.
Área de Protección de Flora y Fauna Nevado de Toluca (Estado de México). Posee bosques de coníferas y de encino, pastizales y vegetación inducida. Algunas especies representativas son el oyamel (Abies religiosa), el pino de altura (Pinus hartwegii), la habichuela (Lupinus mexicanus), el encino (Quercus laurina) y el ratón de los volcanes (Neotomodon alstoni).
Un análisis de las plantas vasculares endémicas de la Faja Volcánica Transmexicana y su superposición con el mapa de áreas naturales protegidas y regiones terrestres prioritarias (Alcántara y Paniagua, 2007) permitió determinar que las áreas más ricas de esta provincia están incluidas al menos parcialmente en alguna área protegidas. Algunas áreas no incluidas en el sistema de áreas protegidas que los autores identificaron son Singuilucan y Tozayuca (Hidalgo), Amelco (Querétaro y Michoacán), el oeste del nevado de Toluca (Estado de México), y Tapalpa y Tecolotlán (Jalisco).
Provincia de la Sierra Madre del Sur
Provincia Austro-Occidental Smith, 1941: 108.
Provincia de la Sierra Madre del Sur Goldman y Moore, 1945: 358; Moore, 1945: 218; Stuart, 1964: 350; Casas-Andreu y Reyna-Trujillo, 1990: mapa; Ramírez-Pulido y Castro-Campillo, 1990: mapa; Arriaga et al., 1997: 65; Morrone, 2001: 39, 2005: 237; Espinosa-Organista et al., 2008: 57; Morrone, 2017a: 74, 2017b: 711; Morrone et al., 2017: 278.
Región de la Sierra y Mesa del Sur West, 1964: 368.
Provincia de las Serranías Meridionales (en parte) Rzedowski, 1978: 103; Rzedowski y Reyna-Trujillo, 1990: mapa.
Provincia Jalisqueño-Guerrerense Ferrusquía-Villafranca, 1990: mapa.
Ecorregión de los Bosques de Pino y Roble de la Sierra Madre del Sur Dinerstein et al., 1995: 103.
Área de las Tierras Altas de la Sierra Madre del Sur Flores-Villela y Goyenechea, 2001: 174.
Área de la Sierra Madre del Sur Marshall y Liebherr, 2000: 206.
Ecorregión de la Sierra Madre del Sur Abell et al., 2008: 408.
Área de las Tierras Altas del Sur de México Flores-Villela y Martínez-Salazar, 2009: 820.
Esta provincia se ubica en el centro-sur de México, en los estados de Jalisco, Colima, Michoacán, Guerrero, Oaxaca y partes de Puebla, por encima de los 1,000 m de altitud (Espinosa et al., 2016; Morrone, 2017a, b). Corresponde al sistema montañoso de la Sierra Madre del Sur, que corre en dirección noroeste-sureste paralelamente a la costa del océano Pacífico, desde Jalisco hasta el istmo de Tehuantepec. Su continuidad está interrumpida por una serie de valles, cuyos ríos se sitúan generalmente por encima de los 1,000 m, pudiendo dividirse en 4 porciones: Jalisciense, Michoacana, Guerrerense y Oaxaqueña. Posee una gran diversida climática, con climas subhúmedos con lluvias de verano, climas húmedos con lluvias abundantes todo el año, climas secos y climas áridos y semiáridos (Hernández-Cerda et al., 2016).
En la vegetación predominan los bosques templados, en especial los de coníferas, pino-encino y mesófilos de montaña; también hay matorrales xerófitos con cactos y selva baja caducifolia en las zonas de contacto con las provincias de las Tierras Bajas del Pacífico y de la Cuenca del Balsas (Espinosa et al., 2016; Dinerstein et al., 1995; Rzedowski, 1978). Entre los géneros dominantes de plantas figuran Hintonella, Microspermum, Omiltemia, Peyritschia, Pinus, Juniperus, Quercus y Silviella (Rzedoswki, 1978).
Algunos taxones endémicos de esta provincia son el acahual (Montanoa grandifolia), el zomplante (Bursera glabrifolia), 1 gorgojo (Pantomorus longulus), el vencejo frentiblanco (Cypseloides storeri), el colibrí oaxaqueño (Eupherusa cyanophrys), el colibrí cola blanca (E. policerca), el rascador sureño (Arremon kuehneri) y el ratón pardo (Peromyscus megalops).
Regionalización biogeográfica. Dentro de esta provincia se han reconocido 3 subprovincias y 5 distritos (Fig. 22) (Santiago-Alvarado et al., 2016; Morrone, 2017b).
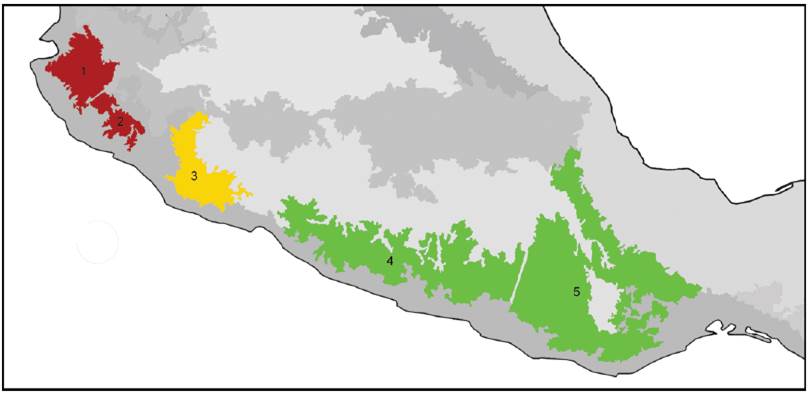
Subprovincia de la Sierra Madre del Sur Occidental
Subprovincia Occidental Ferrusquía-Villafranca, 1990: mapa.
Subprovincia de la Sierra Madre del Sur Occidental (en parte) Santiago-Alvarado et al., 2016: 439.
Subprovincia de la Sierra Madre del Sur Occidental Morrone, 2017b: 711.
Corresponde a los estados de Jalisco, Colima y Michoacán. Comprende los distritos Jalisciense y Jalisciense-Manantlán.
Distrito Jalisciense
Distrito Jalisciense Moore, 1945: 218; Morrone, 2014a: 32, 2017b: 711.
Distrito Oeste Miranda y Luna, 2007: 512.
Distrito Jaliscience-Tuito Santiago-Alvarado et al., 2016: 439.
En la parte norte de la subprovincia, en el estado de Jalisco. Sustentado por la presencia de los encinos Quercus cualensis y Q. tuitensis.
Distrito Jalisciense-Manantlán
Distrito Jaliscience-Manantlán Santiago-Alvarado et al., 2016: 440; Morrone, 2017b: 711.
En la parte sur de la subprovincia, en los estados de Jalisco, Michoacán y norte de Colima. Sustentado por la presencia de una especie de Lauraceae (Beilschmiedia manantlanensis), un álamo (Populus guzmanantlanensis), una especie de nemátodo (Rhabdias manantlanensis) y un escarabajo (Canthon occidentalis).
Subprovincia de la Sierra Madre del Sur Central
Subprovincia de la Sierra Madre del Sur Occidental (en parte) Santiago-Alvarado et al., 2016: 439.
Subprovincia de la Sierra Madre del Sur Central Morrone, 2017b: 711.
Comprende el centro de la provincia, en el estado de Michoacán. Solo comprende el distrito de Michoacán.
Distrito de Michoacán
Distrito de Michoacán Santiago-Alvarado et al., 2016: 440; Morrone, 2017b: 711.
Con la misma circunscripción que la provincia. Sustentado por la presencia del pino de Rzedowski (Pinus rzedowskii), el torote (Bursera confusa) y el ratón zagaz (Peromyscus sagax).
Subprovincia de la Sierra Madre del Sur Oriental
Subprovincia de la Sierra Madre del Sur Oriental Santiago-Alvarado et al., 2016: 440; Morrone, 2017b: 711.
Corresponde al oriente de la provincia, en los estados de Guerrero, Oaxaca y parte de Puebla. Posee 2 distritos: Guerrerense y de las Tierras Altas Oaxaqueñas.
Distrito Guerrerense
Provincia Guerrerense Smith, 1941: 108; Ramírez-Pulido y Castro-Campillo, 1990: mapa; Brown et al., 1998: 30. Provincia de la Sierra Madre del Sur-Guerrero Escalante et al., 1998: 284.
Distrito Guerrerense Morrone, 2014a: 33, 2017b: 712.
En la parte más occidental, situada en el estado de Guerrero. Sustentado por la presencia de la flor de tigre (Tigridia hintonii), 1 encino (Quercus rubramenta), 1 carábido (Elliptoleus whiteheadi), 2 escarabajos (Cotinis ibarrai y Onthophagus bassariscus), el lagarto-escorpión (Mesaspis gadovii gadovii), la coqueta cresticorta (Lophornis brachylophus), 2 ratones (Handleyomys guerrerensis y Reithrodontomys bakeri) y el conejo de Omiltemi (Sylvilagus insonus).
Distrito de la Altiplanicie Oaxaqueña
Provincia de la Altiplanicie Oaxaqueña Smith, 1941: 107.
Subprovincia Valles-Centraliana Ferrusquía-Villafranca, 1990: mapa.
Subprovincia Ístmica Ferrusquía-Villafranca, 1990: mapa.
Ecorregión de los Bosques de Pino y Roble de la Sierra Juárez Dinerstein et al., 1995: 102.
Provincia de Oaxaca Arriaga et al., 1997: 65; Espinosa-Organista et al., 2008: 57.
Provincia del Nudo de Zempoaltépetl Escalante et al., 1998: 284.
Provincia de la Sierra Madre del Sur-Oaxaca Escalante et al., 1998: 284.
Provincia de la Sierra de Miahuatlán Escalante et al., 1998: 284.
Distrito de los Valles Centrales Morrone, 2014a: 33.
Distrito Ístmico Morrone, 2014a: 33.
Distrito del Nudo de Zempoaltépetl Morrone, 2014a: 33.
Distrito de la Sierra de Mihuatlán Morrone, 2014a: 33.
Distrito de las Tierras Altas Oaxaqueñas Morrone, 2014a: 33,2017b: 712.
Distrito Oaxaqueño Santiago-Alvarado et al., 2016: 442.
En el estado de Oaxaca, principalmente en las Sierras Mixteca y Norte. Sustentado por la presencia de especies de cícadas (Ceratozamia mixeorum y C. whitelockiana), de copal (Bursera altijuga, B. arida, B. biflora y B. heliae), 1 cacto (Weberocereus alliodorus), 2 especies de carábidos (Elliptoleus crepericornis y E. zapotecorum), 1 gorgojo (Phacepholis globicollis), 2 ratones (Habromys chinanteco y Microtus oaxacensis) y 2 especies de musarañas (Cryptotis peregrina y C. phillipsii).
Evolución biótica. Esta provincia muestra afinidades bióticas con todas las demás provincias de la Zona de Transición Mexicana, así como también con las provincias neotropicales de las Tierras Altas del Pacífico y la Cuenca del Balsas. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia de la Sierra Madre del Sur se relaciona estrechamente con las provincias de las Tierras Bajas del Pacífico, Faja Volcánica Transmexicana y Cuenca del Balsas. Un análisis de trazos basado en especies de coleópteros (Morrone y Márquez, 2001) mostró que esta provincia se relaciona con las provincias de la Sierra Madre Occidental, Cuenca del Balsas, Sierra Madre Oriental y Faja Volcánica Transmexicana. Un análisis de parsimonia de endemismos basado en especies de mamíferos (Carleton et al., 2002) mostró una relación estrecha con la provincia de las Tierras Altas de Chiapas. Otro análisis de parsimonia de endemismos, basado en aves y utilizando subcuencas como unidades del análisis (Blancas-Calva et al., 2010), halló un patrón interesante, pues los agrupamientos correspondieron a áreas con semejanzas climáticas más que reflejar relaciones históricas. De acuerdo con un análisis biogeográfico cladístico basado en taxones de insectos, peces, reptiles y plantas (Marshall y Liebherr, 2000), la provincia de la Sierra Madre del Sur se relaciona con la provincia de la Faja Volcánica Transmexicana.
Conservación. La provincia de la Sierra Madre del Sur se halla amenazada por la construcción de carreteras, el desarrollo turístico, el sobrepastoreo, la caza excesiva y las presiones por la urbanización (Dinerstein et al., 1995). En esta provincia se encuentra la siguiente área natural protegida:
Parque Ecológico Omiltemi (Guerrero). Área con bosques de pino, pino-encino y mesófilo de montaña, entre otros. Luna-Vega y Llorente-Bousquets (1993) compendiaron el conocimiento geográfico, geomorfológico, climático, florístico, faunístico y biogeográfico del parque.
Provincia de las Tierras Altas de Chiapas
Provincia de la Altiplanicie Chiapaneca Smith, 1941: 109.
Provincia las Tierras Altas de Chiapas Goldman y Moore, 1945: 358; Morrone, 2014a: 34, 2017a: 77; Morrone et al., 2017: 278.
Provincia Chiapanense Barrera, 1962: 101.
Provincia de las Tierras Altas de Chiapas-Guatemala Ryan, 1963: 23; Stuart, 1964: 350.
Región de las Tierras Altas de América Central West, 1964: 368.
Centro del Bosque Montano de América Central Müller, 1973: 14.
Provincia de las Serranías Transístmicas Rzedowski, 1978: 103; Rzedowski y Reyna-Trujillo, 1990: mapa.
Provincia de la Sierra Madre de Chiapas Casas-Andreu y Reyna-Trujillo, 1990: mapa; Rzedowski y ReynaTrujillo, 1990: mapa.
Provincia Chiapasense Ferrusquía-Villafranca, 1990: mapa.
Provincia de Chiapas Ramírez-Pulido y CastroCampillo, 1990: mapa; Morrone, 2001: 45, 2005: 240. Ecorregión de los Bosques Montanos de América Central Dinerstein et al., 1995: 92.
Ecorregión de los Bosques de Pino-Encino de América Central Dinerstein et al., 1995: 97.
Ecorregión de los Bosques Húmedos de la Sierra Madre Dinerstein et al., 1995: 92.
Provincia de Los Altos de Chiapas Arriaga et al., 1997: 64; Espinosa-Organista et al., 2008: 58.
Área de las Tierras Altas de Chiapas y Guatemala Flores-Villela and Goyenechea, 2001: 174.
Ecorregión de Chiapas-Fonseca Abell et al., 2008: 408.
Esta provincia se ubica en el sur de México, Guatemala, Honduras, El Salvador y Nicaragua; básicamente corresponde a la Sierra Madre de Chiapas, desde los 500 hasta los 4,000 m de altitud (Morrone, 2017a). Topográficamente presenta un patrón complejo, con volcanes de hasta 4,000 m de altura y valles profundos, y 2 sistemas montañosos diferentes, la Sierra Madre de Chiapas y el Macizo Central de Chiapas, que constituye la proyección septentrional del sistema montañosos Centroamericano y que alcanza su mayor altitud (2,860 m) en la región de San Cristóbal. Entre ambos sistemas se sitúa la Cuenca o Depresión Central de Chiapas, que es el amplio valle del río Grijalva.
La vegetación consiste en bosques de pino-encino, sabanas, bosque tropical caducifolio y matorrales (Dinerstein et al., 1995).
Algunos taxones endémicos de esta provincia son: un estafilínido (Bledius stenuus), la rata arborícola chiapaneca (Tylomys bullaris) y la nauyaca verdinegra (Bothriechis aurifer). Un aspecto importante de la provincia de las Tierras Altas de Chiapas es que en ella se encuentra la distribución más austral de varios taxones neárticos; un ejemplo lo constituye el género de ratones Microtus (Conroy et al., 2001).
Regionalización biogeográfica. Dentro de esta provincia se han reconocido 6 distritos (Morrone, 2014a, 2017a): Sierramadrense, Comitanense, Lacandón, Soconusco, Tierras Altas de Guatemala y Nicaragüense.
De ellos, los 4 primeros se encuentran en nuestro país.
Distrito Sierramadrense
Subprovincia Sierra-Madreña Ferrusquía-Villafranca, 1990: mapa.
Provincia de la Sierra Madre de Chiapas Escalante et al., 1998: 284.
Distrito Sierramadrense Morrone, 2014a: 35.
Corresponde a la porción más occidental de la provincia, en el este de Oaxaca.
Distrito Comitanense
Subprovincia Comitania Ferrusquía-Villafranca, 1990: mapa.
Distrito Comitanense Morrone, 2014a: 35.
En el este de Oaxaca y Chiapas.
Distrito Lacandón
Subprovincia Lacandoniana Ferrusquía-Villafranca, 1990: mapa.
Distrito Lacandón Morrone, 2014a: 35.
En el norte de la provincia, en el estado de Chiapas.
Distrito del Soconusco
Provincia del Soconusco Rzedowski, 1978: 109; Rzedowski y Reyna-Trujillo, 1990: mapa; Arriaga et al., 1997: 64; Espinosa-Organista et al., 2008: 58.
Distrito del Soconusco Morrone, 2014a: 35.
En partes bajas de la Sierra Madre de Chiapas y Guatemala. Sustentado por la presencia de: 1 helecho (Asplenium solmsii), 1 cícada (Zamia soconuscensis), 1 encino (Quercus durantifolia) y 1 lagartija (Abronia smithi).
Evolución biótica. Además de las afinidades bióticas con otras provincias de la Zona de Transición Mexicana, esta provincia exhibe afinidades con las provincias Veracruzana y de Las Tierras Bajas del Pacífico. Rzedowski (1978) destaca las relaciones de la flora de esta provincia con la de las Sierras Madre Occidental, Oriental y del Sur, y la de la Faja Volcánica Transmexicana, aunque carece de algunos géneros neárticos como Cercocarpus, Heuchera, Pedicularis, Silene y Taxus. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia de las Tierras Altas de Chiapas se relaciona estrechamente con la provincia Veracruzana. Otro análisis de parsimonia de endemismos, basado en especies de mamíferos (Carleton et al., 2002), mostró una relación estrecha entre la provincia de la Sierra Madre del Sur y la de las Tierras Altas de Chiapas. Un análisis de trazos basado en especies de coleópteros (Morrone y Márquez, 2001) mostró que esta provincia se relaciona con las provincias Veracruzana y de las Tierras Bajas del Pacífico.
Gutiérrez-García y Vázquez-Domínguez (2012) analizaron la historia evolutiva del género monotípico de roedores Ototylomys. La distribución de su especie O. phyllotis desde el istmo de Tehuantepec hasta Costa Rica permite realizar inferencias biogeográficas sobre esta interesante área, que incluye la provincia de las Tierras Altas de Chiapas. Los autores analizaron 129 muestras de O. phyllotis que cubren el área de distribución. Aislaron su ADN y amplificaron y secuenciaron un gen mitocondrial.
Realizaron análisis filogenéticos y estimaron los tiempos de divergencia, la diversidad genética y la historia demográfica. El análisis filogenético permitió identificar 3 linajes principales (Fig. 23): uno de la Península de Yucatán, otro de las Tierras Altas de Chiapas-Guatemala y un tercero en el Núcleo Centroamericano. El tiempo de divergencia inicial se calculó en unos 3.85 ma y la especie se habría originado en un área cercana a Honduras y El Salvador. De allí se habría dispersado hacia el norte hacia Guatemala y Chiapas, hace unos 2.84 ma. Durante los últimos 800,000 años, O. phyllotis habría expandido su área de distribución hacia la Península de Yucatán y las tierras bajas de Nicaragua. El análisis filogeográfico encontró 2 grupos de haplotipos, uno en América Central (excepto Belice y el norte de Guatemala) y el otro en el resto de la distribución. De acuerdo con Gutiérrez-García y Vázquez-Domínguez (2012), los resultados obtenidos pueden explicarse teniendo en cuenta la historia tectónica del área. La especie O. phyllotis pertenece a un grupo norteamericano que se habría dispersado hacia el sur hace unos 3.35 ma, en que habría llegado a América Central y desde allí se habría dispersado a las otras áreas. Los eventos de dispersión y diversificación de esta especie se relacionarían con el clima cálido y la vegetación tropical que habrían existido durante lo que se conoce como el Gran Intercambio Biótico Americano.
Figura 23 Reconstrucción genealógica de Ototylomys phyllotis llevado a cabo por Gutiérrez-García y Vázquez-Domínguez (2012), con las áreas de distribución de los linajes.
Conservación. La provincia de las Tierras Altas de Chiapas se halla amenazada por la conversión de hábitats para la producción de café, la recolección de leña, la conversión de hábitats naturales para la agricultura, el desmonte para la creación de pastizales, el sobrepastoreo y los fuegos antropogénicos (Dinerstein et al., 1995).
Región Neotropical
La región Neotropical comprende los trópicos del Nuevo Mundo, en el centro y sur de México, América
Central, las Antillas y la mayor parte de América del Sur (Morrone, 2017a). Sus límites varían de acuerdo con distintos autores; en América del Sur la porción Andina y la Patagonia se asignan a la región Andina, sobre todo teniendo en cuenta la distribución de numerosos taxones de plantas y animales (Morrone, 2014a, 2017a).
La región Neotropical integra el reino Holotropical, el cual incluye las áreas tropicales del mundo entre los 30° de latitud sur y los 30° de latitud norte. Las otras regiones incluidas dentro del reino Holotropical son la Etiópica (África central, Madagascar e islas del océano Pacífico occidental) y la Oriental (India, Burma, Malasia, Indonesia, las Filipinas y las islas del Pacífico). El reino Holotropical corresponde a la Gondwana occidental.
La región Neotropical se caracteriza por la presencia de numerosos taxones endémicos de plantas y animales (Morrone, 2014a, 2017a; Müller, 1973). Entre las plantas, destacan las familias Bromeliaceae, Cactaceae y Heliconiaceae. Beron (2018) ha considerado a la región Neotropical como una de las más ricas en taxones de arácnidos, pertenecientes a los Palpigradi, Solifugae, Ricinulei, Scorpiones, Pseudoscorpiones, Opiliones, Amblypygi, Uropygi, Schizomida, Araneae y Acari, citando numerosos endemismos, en especial de Opiliones. Existen varias familias endémicas de aves, como Trochilidae (colibríes), Ramphastidae (tucanes), Furnariidae (horneros), Thamnophilidae (hormigueros, batarás y tiluchíes) y Thraupidae (tángaras). Familias endémicas de mamíferos incluyen Didelphidae (tlacuaches), Bradypodidae (perezosos), Myrmecophagidae (osos hormigueros), Aotidae (micos nocturnos), Atelidae (monos aulladores), Callitrichidae (titíes), Cebidae (monos capuchinos), Pitheciidae (sahuíes y sakíes), Caviidae (cuyos y capibaras), Octodontidae (degúes), Echimyidae (puercoespines con cola), Natalidae (murciélagos) y Desmodontidae (vampiros). Existen también numerosos géneros y especies endémicas de esta región.
Regionalización biogeográfica
En México, integran la región Neotropical las 5 provincias asignadas a la Zona de Transición Mexicana junto con las provincias de las Tierras Bajas del Pacífico, Cuenca del Balsas, Veracruzana y Península de Yucatán. Estas 3 últimas se han asignado al dominio Mesoamericano, perteneciente a la subregión Brasileña (Morrone, 2017a). Ríos-Muñoz (2013) ha reconocido esta área como "Mesoamérica biótica", considerando, sin embargo, que sus límites no son fijos sino que corresponden a gradientes de intercambio con áreas aledañas.
Evolución biótica
Morrone (2014b) llevó a cabo un análisis biogeográfico cladístico de la región Neotropical basado en 36 taxones (insectos, arácnidos, vertebrados y plantas). El objetivo principal fue corroborar que ésta es una región natural. El análisis utilizó 8 áreas previamente reconocidas como unidades (Mesoamérica, las Antillas, noroeste de América del Sur, norte de Amazonia, sureste de Amazonia, sudoeste de Amazonia, Chaco y Paranaense), así como 4 áreas "externas" (las regiones Andina y Neártica y las zonas de transición Mexicana y Sudamericana). Se seleccionaron 36 taxones para los cuales había análisis filogenéticos disponibles. A partir de sus cladogramas taxonómicos de áreas, se extrajeron los subárboles libres de paralogía, se codificaron sus componentes y se los volcó a una matriz de datos. La matriz se analizó con el programa TNT (Goloboff et al., 2008). El cladograma general de áreas obtenido muestra que las áreas neotropicales constituyen una unidad natural, la Zona de Transición Mexicana resulta hermana de la región Neártica y la Zona de Transición Sudamericana resulta hermana de la región Andina (Fig. 24). Dentro de la región Neotropical, las Antillas resultan el grupo hermano de las áreas continentales, las cuales a su vez se agrupan en una porción noroccidental y otra suroriental. Dentro de la noroccidental se encuentra Mesoamérica, que corresponde a las tierras bajas tropicales de nuestro país y América Central, y el área hermana del noroeste de la Amazonia. Como resultado de este análisis, Mesoamérica se considera un dominio de la subregión Brasileña, de la región Neotropical (Morrone, 2004a). La dispersión de la biota neotropical a Mesoamérica habría ocurrido entre el Jurásico Temprano y el Cretácico Temprano (190148 ma) al conectarse América del Norte y América del Sur. La vicarianza entre Mesoamérica y el noroeste de la Amazonia sería un evento más reciente (Plioceno).

Figura 24 Cladograma general de áreas de la región Neotropical obtenido por Morrone (2014b), superpuesto con el mapa del área.
En cuanto a la evolución biótica del dominio Mesoamericano en particular, Becerra (2005) llevó a cabo un análisis del origen y dispersión del bosque tropical estacionalmente seco en México. La autora analizó al género Bursera (Burseraceae), considerándolo como un buen indicador de la historia de este bioma, obteniendo una filogenia calibrada, a partir de la cual estimó las tasas de diversificación en los últimos 60 ma. Los resultados mostraron que hace unos 20-30 ma Bursera comenzó una diversificación rápida, cuando las condiciones fueron favorables para el establecimiento del bosque tropical estacionalmente seco. La dispersión ocurrió inicialmente en el oeste y luego hubo una expansión hacia el sur y el este. La formación de los sistemas montañosos de México habría sido fundamental para el establecimiento y la persistencia de este bioma. En un análisis posterior, DeNova et al. (2012) estimaron una antigüedad mayor, concluyendo que al menos algunos linajes de Bursera resultaron de un largo aislamiento más que de especiación rápida. La expansión de los bosques tropicales estacionalmente secos del dominio Mesoamericano hacia el centro y norte de México se asociaría con la aridez creciente del Mioceno.
Provincia de las Tierras Bajas del Pacífico
Región de las Tierras Bajas del Pacífico West, 1964: 368.
Subregión de las Tierras Bajas Occidentales Savage, 1966: 736.
Provincia Xerófila de Montaña (en parte) Cabrera y Willink, 1973: 34.
Centro Pacífico Centroamericano Müller, 1973: 19.
Provincia Guerrerense Udvardy, 1975: 41.
Provincia de la Costa Pacífica Rzedowski, 1978: 107; Rzedowski y Reyna-Trujillo, 1990: mapa; Arriaga et al., 1997: 62.
Provincia de la Costa Pacífica de América Central Samek et al., 1988: 28.
Provincia Mexicana Occidental Casas-Andreu y Reyna-Trujillo, 1990: mapa.
Provincia Pacifiquense Ferrusquía-Villafranca, 1990: mapa
Ecorregión de los Bosques Secos del Pacífico Centroamericano Dinerstein et al., 1995: 94.
Área de las Tierras Bajas del Pacífico y Cuenca del Balsas (en parte) Flores-Villela y Goyenechea, 2001: 174.
Provincia de la Costa Pacífica Mexicana Morrone, 2001: 40, 2005: 239.
Provincia del Pacífico Espinosa-Organista et al., 2008: 60.
Provincia de las Tierra Bajas del Pacífico Morrone, 2014a: 46, 2017a: 167; Morrone et al., 2017: 278.
Esta provincia corresponde a una franja angosta e ininterrumpida en la costa pacífica de México (estados de Sinaloa, Nayarit, Jalisco, Colima, Michoacán, Guerrero, Oaxaca y Chiapas), Guatemala, El Salvador, Honduras, Nicaragua y Costa Rica (Morrone, 2017a). Usualmente no supera los 400 m de altitud. También incluye el archipiélago de las Islas Revillagigedo, situado a 350 km del continente.
La vegetación consiste en bosques tropicales estacionalmente secos, bosques tropicales húmedos, sabanas y palmares (Dinerstein et al., 1995). Esta provincia alberga la mayor concentración de bosques tropicales estacionalmente secos del país (Linares-Palomino et al., 2011). La flora incluye numerosas especies de Fabaceae, Euphorbiaceae, Cactaceae, Burseraceae, Asteraceae, Malpighiaceae, Rubiaceae y Anacardiaceae (Trejo, 2010). Para estos bosques se han identificado distintos patrones de distribución basados en especies de plantas (Lott y Atkinson, 2010): noroeste, Jalisco, costa central, México, ChiapasGuatemala, América Central y de distribución amplia.
Algunos taxones endémicos de esta provincia son: el maguey del Grijalva (Agave kewensis), el copal (Bursera excelsa), un gorgojo (Phacepholis albicans), el copetón piquiplano (Deltarhynchus flammulatus), el murciélago con bonete de Underwood (Eumops underwoodi), el zorrillo pigmeo (Spilogale pygmaea) y la rata arrocera (Oryzomys melanotis).
Regionalización biogeográfica. Dentro de esta provincia se han identificado 6 distritos (Morrone, 2017a): Sinaloense, de las Islas Revillagigedo, Nayarit-Guerrero, Tehuano, de las Islas Tres Marías y Tapachulteco. El último se extiende fuera de nuestro país.
Distrito Sinaloense
Provincia Sinaloense Goldman y Moore, 1945: 355; Stuart, 1964: 350; Udvardy, 1975: 41; Ferrusquía-Villafranca, 1990: mapa; Brown et al., 1998: 32.
Distrito Costero Sinaloense Moore, 1945: 218.
Distrito Tebaca Moore, 1945: 218.
Ecorregión de los Bosques Secos de Sinaloa Dinerstein et al., 1995: 99.
Provincia de la Costa Oeste, M Escalante et al., 1998: 284.
Provincia de la Costa Oeste, N Escalante et al., 1998: 284.
Sección Norte García-Trejo y Navarro, 2004: 177.
Distrito Sinaloense Morrone, 2014a: 47.
Porción más septentrional de la provincia, en el estado de Sinaloa. Corresponde al área septentrional delimitada por García-Trejo y Navarro (2004), con base en la distribución de especies de aves. Sustentado por la presencia del cuervo de Sinaloa (Corvus sinaloae) y la urraca de lomo morado (Cyanocorax beechei).
Distrito de las Islas Revillagigedo
Provincia de Revillagigedo Goldman y Moore, 1945: 352.
Provincia de las Islas Revillagigedo Rzedowski, 1978: 108; Samek et al., 1988: 28; Rzedowski y Reyna-Trujillo, 1990: mapa; Escalante et al., 1998: 284.
Provincia Revillagigedo Espinosa-Organista et al., 2008: 59.
Distrito de las Islas Revillagigedo Morrone, 2014a: 47.
Archipiélago situado a 382 km del Cabo San Lucas, Baja California Sur, y a 661 km de Manzanillo, Colima (Conanp, 2004). Comprende 4 islas volcánicas: Socorro, San Benedicto, Clarión y Roca Partida. Socorro es la mayor, alcanzando los 1,000 m de altitud en el volcán Evermann, cuyas erupciones comenzaron en el Mioceno y la última data de 1948. La isla San Benedicto es la cúspide de una montaña submarina, originada probablemente en el Pleistoceno; el volcán Bárcena, cuya última erupción ocurrió en 1952, alcanza los 374 m de altitud. La isla Clarión es la más alejada del continente y se formó a partir de erupciones volcánicas del Mioceno al Eoceno. Roca Partida es la cima de un estratovolcán submarino. Solo Socorro y Clarión poseen vegetación, ya que la de San Benedicto fue destruida en la última erupción y Roca Partida no posee ningún suelo para sustentarla. La vegetación de la isla Socorro está bien caracterizada (Flores-Palacios et al., 2009), incluyendo matorrales, pastizales, bosque tropical caducifolio, bosque tropical perennifolio y bosque mesófilo de montaña. La vegetación de Clarión es básicamente arbustiva, con numerosas plantas anuales y cactáceas. La endemicidad del archipiélago es muy marcada (Rzedowski, 1991), con 30 especies de plantas endémicas de la isla Socorro y numerosos artrópodos y vertebrados. Entre las aves destacan el perico de Socorro (Aratinga holochlora brevipes), el saltapared (Troglodytes sissonni), el rascador moteado de isla Socorro (Pipilo socorroensis), la tortolita de Socorro (Columbina passerina socorroensis), el cenzontle (Mimodes graysoni) y el halcón cola roja de Socorro (Buteo jamaicensis socorrensis). Tres especies de aves han sido reportadas como extintas: la tórtola de Socorro (Zenaida graysoni), el tecolote enano (Micrathene whytneyi graysoni) y la matraquita de San Benedicto (Salpinctes obsoletus exsuls). De la tórtola de Socorro existen poblaciones en cautiverio en Alemania, EUA y México y se planea reintroducirla en la isla. La herpetofauna de las islas Revillagigedo comprende 4 especies: 3 lagartijas (Urosaurus clarionensis, U. auriculatus y Hemidactylus frenatus, esta última introducida) y 1 serpiente, Masticophis anthonyi, endémica de Clarión. En cuanto a los artrópodos, existe más de un centenar de especies de insectos y arácnidos, siendo pocas de ellas endémicas del archipiélago (Conanp, 2004).
Distrito de Nayarit-Guerrero
Provincia de Nayarit-Guerrero Goldman y Moore, 1945: 355; Stuart, 1964: 350.
Área de las Tierras Bajas Costeras de Nayarit-Sinaloa West, 1964: 368.
Provincia Guerrerense Udvardy, 1975: 41.
Provincia Nayaritense Ferrusquía-Villafranca, 1990: mapa.
Ecorregión de los Bosques Secos de Jalisco Dinerstein et al., 1995: 99.
Ecorregión del Matorral de Cactos de Guerrero Dinerstein et al., 1995: 111.
Ecorregión de las Sabanas de Palmas de Jalisco Dinerstein et al., 1995: 106.
Provincia de Nayarit Brown et al., 1998: 28.
Provincia de la Costa Oeste, S Escalante et al., 1998: 284.
Sección Central García-Trejo y Navarro, 2004: 177. Distrito de Nayarit-Guerrero Morrone, 2014a: 47.
En el área costera de Nayarit, Jalisco, Colima, Michoacán y Guerrero. Corresponde al área central delimitada por García-Trejo y Navarro (2004), con base en la distribución de especies de aves.
Distrito de las Islas Tres Marías
Provincia de las Islas Tres Marías Escalante et al., 1998: 284.
Distrito de las Islas Tres Marías Morrone, 2014a: 47.
Situadas a 112 km de la costa de Nayarit. Consta de 3 islas, llamadas María Madre, María Magdalena y María Cleofas. Algunos taxones endémicos de estas islas incluyen al ratón de las Islas Tres Marías (Peromyscus madrensis), el mapache de las Islas Tres Marías (Procyon insularis), el turpial de fuego (Icterus graysonii) y el loro de cabeza amarilla de las Islas Tres Marías (Amazona tresmariae).
Distrito Tehuano
Provincia Tehuana Smith, 1941: 110.
Provincia Tapachulteca Smith, 1941: 110.
Provincia de Tehuantepec Goldman y Moore, 1945: 358; Stuart, 1964: 350.
Provincia Tehuantepecana Barrera, 1962: 101.
Provincia del Pacífico de México-Nicaragua (en parte) Stuart, 1964: 350.
Área de las Tierras Bajas Costeras de América Central West, 1964: 368.
Provincia del Oeste de México Savage, 1966: 736.
Subprovincia Istmeña Ferrusquía-Villafranca, 1990: mapa.
Subprovincia Tehuantepecana Ferrusquía-Villafranca, 1990: mapa.
Ecorregión de las Sabanas de Tehuantepec Dinerstein et al., 1995: 104.
Provincia de las Tierras Bajas del Istmo Escalante et al., 1998: 284.
Sección Sur García-Trejo y Navarro, 2004: 177. Distrito Tehuano Morrone, 2014a: 48.
En la costa pacífica del Istmo de Tehuantepec; corresponde al área más austral de García-Trejo y Navarro (2004). Sustentado por la presencia del cucarachero de Chiapas (Campylorhynchus chiapensis), el colorín ventrirrosado (Passerina rositae) y el gorrión oaxaqueño (Peucaea sumichrasti).
Evolución biótica. Ésta es una provincia claramente neotropical, mostrando sus principales afinidades con las provincias de la Cuenca del Balsas y Veracruzana, aunque también posee elementos que relacionan su biota con la de las provincias transicionales de la Sierra Madre del Sur y las Tierras Altas de Chiapas. Müller (1973) consideró que su fauna de vertebrados se relaciona estrechamente con la de la Península de Yucatán. Rzedowski (1978) destacó el número elevado de especies de plantas endémicas de esta provincia, aunque muchas son comunes con la provincia de la Cuenca del Balsas. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia de las Tierras Bajas del Pacífico se relaciona estrechamente con las provincias de la Sierra Madre del Sur, Faja Volcánica Transmexicana y Cuenca del Balsas. Un análisis de trazos basado en especies de coleópteros (Morrone y Márquez, 2001) mostró que esta provincia se relaciona con las provincias Puntarenas-Chiriquí, Veracruzana y de las Tierras Altas de Chiapas.
Arbeláez-Cortés et al. (2014) analizaron la estructura filogeográfica de 3 especies de aves endémicas de las Tierras Bajas del Pacífico, con el objeto de determinar su existía concordancia entre estas especies y si ésta se correlacionaba con la geografía, ecología y clima. Los autores analizaron 3 especies pertenecientes a diferentes familias: el momoto mexicano (Momotus mexicanus, Momotidae), el carpintero enmascarado (Melanerpes chrysogenis, Picidae) y el colorín ventridorado (Passerina leclancherii, Cardinalidae). Obtuvieron muestras de tejidos de colecciones y trabajo de campo, extrajeron el ADN y amplificaron y secuenciaron genes nucleares y mitocondriales para las 3 especies. Realizaron análisis filogenéticos y analziaron la diversidad y divergencia genéticas. Por otra parte, a partir de variables abióticas analizaron si los resultados anteriores se correlacionaban con algunas de éstas en particular. Finalmente, a partir del modelado de nicho ecológico identificaron áreas climáticamente estables. El análisis de ADN mitocondrial de las 3 especies permitió identificar una fuerte estructuración, con quiebres en las mismas áreas. Una división filogeográfica mayor ocurrió en el sureste, aproximadamente en el límite entre Oaxaca y Guerrero; y otra ocurrió en el noroeste, en Michoacán-Jalisco. Las divisiones principales entre las 3 especies habrían ocurrido en el Pleistoceno, especialmente dentro de los últimos 586,000 años. Las áreas climáticamente estables fueron relativamente pequeñas, con coincidencia parcial entre las 3 especies. Los resultados obtenidos permiten deducir la existencia de un patrón común a las especies analizadas, que implicaría una asociación con la historia paleoecológica del área, aunque sin una asociación clara con barreras a la dispersión. Las fluctuaciones climáticas serían el factor principal para explicar la diversificación de estas especies.
Las principales afinidades biogeográficas de las islas Revillagigedo son con la costa Pacífica de México (Brattstrom, 1990). El análisis filogenético de las especies de Urosaurus (Feldman et al., 2011) mostró que las especies de Clarión (U. clarionensis) y Socorro (U. auriculatus) son hermanas, y la especie hermana de ambas se distribuye en el noroeste de México y suroeste de los EUA, con lo cual habría bastado un solo evento de dispersión del continente a las islas y otro posterior entre éstas.
La especie Icterus graysonii, ave endémica de las Islas Tres Marías, fue estudiada por Cortés-Rodríguez et al. (2008), quienes hallaron que su separación de las especies afines del continente habría ocurrido hace relativamente bastante tiempo, aunque estos autores no descartan la posibilidad de un evento de dispersión más reciente y la fijación de una mutación rara o actualmente extinguida en el continente.
Conservación. La provincia de las Tierras Bajas del Pacífico se halla amenazada por la expansión de los cultivos de café y cítricos, el sobrepastoreo, la deforestación, los incendios provocados, la explotación de la vida silvestre, la ganadería extensiva, la minería y la demanda de agua (Maass et al., 2010). Los bosques tropicales estacionalmente secos se consideran entre los ecosistemas más amenazados, principalmente por sus altas tasas de deforestación (García-Oliva y Jaramillo, 2011). De acuerdo con Trejo (2010), la tasa de deforestación en ciertas áreas llega al 1.4%, aunque no existe información precisa para toda el área. Balvanera et al. (2011) han analizado los servicios ambientales que proveen estos bosques, destacando la regulación de la fertilidad del suelo, la erosión, el ciclo del agua, polinizadores, vectores de enfermedades, especies invasoras e impacto a eventos meteorológicos extremos. En esta provincia se incluyen las siguientes áreas naturales protegidas (Ceballos et al., 2010; Conanp, 2018):
Reserva de la Biosfera Archipiélago de Revillagigedo (Colima). Es un laboratorio natural dadas sus características insulares. La vegetación incluye pastizal, matorral de crotón, vegetación halófila, selva mediana caducifolia, pradera, matorral crasicaule, selva subperinnifolia, selva baja perennifolia y matorral de guayabillo. Existe un programa de conservación y manejo (Conanp, 2004), destinado a garantizar la integridad del área, promover la conservación de su biota, fomentar la participación social, promover la investigación científica y asegurar el financiamiento de la reserva. Algunas especies representativas son el churristate de playa (Ipomoea pescaprae), el haba de playa (Canavalia rosea), el croton (Croton masonii), el cucubano (Guettarda elliptica) y la pardela (Puffinus auricularis).
Reserva de la Biosfera Chamela-Cuixmala (Jalisco y Colima). Bosque tropical caducifolio y humedales. Constituye uno de los pocos sitios de nuestro país creados específicamente para conservar selva seca (Ceballos y García, 2010). El libro de Noguera et al. (2002) compendia el conocimiento del ambiente físico, fauna, flora, ecosistemas y patrones de uso de esta reserva.
Área de Protección de Flora y Fauna Meseta de Cacaxtla (Sinaloa). Meseta que no supera los 380 m de altitud, en que predomina la vegetación de selva seca. Posee varias especies de plantas amenazadas y su endemicidad de reptiles es alta (Rubio et al., 2010).
Ceballos et al. (2010) realizaron un análisis exhaustivo de la cobertura de los bosques tropicales estacionalmente secos del Pacífico mexicano y de las áreas prioritarias que se han establecido para su conservación. Algunas de las áreas prioritarias más importantes identificadas por estos autores son la Sierra Mayo-Yaqui (Sonora y Chihuahua), la Meseta de Cacaxtla (Sinaloa), la Llanura Costera del Pacífico-Pie de la Sierra de Sinaloa, la Cuenca Alta del Río Santiago (Jalisco, Zacatecas, Nayarit y Durango), las Islas Marías, Chamela-Cuixmala (Jalisco y Colima), Colima Centro (Colima, Jalisco y Michoacán), El Infiernillo (Michoacán y Guerrero), Acahuizotla (Guerrero), Pinotepa-Ometepec (Guerrero y Oaxaca) y Nizanda (Oaxaca).
Provincia de la Cuenca del Balsas
Región de las Cuencas Tropicales Interiores Secas (en parte) West, 1964: 368.
Provincia de la Depresión del Balsas Rzedowski, 1978: 108; Rzedowski y Reyna-Trujillo, 1990: mapa; Arriaga et al., 1997: 62; Morrone, 2001: 38.
Subprovincia Balsasana Ferrusquía-Villafranca, 1990: mapa.
Provincia del Balsas Ramírez-Pulido y CastroCampillo, 1990: mapa; Espinosa-Organista et al., 2008: 61.
Ecorregión de los Bosques Secos de Balsas Dinerstein et al., 1995: 99.
Provincia de la Cuenca del Balsas Morrone, 2001: 38, 2014a: 48, 2017a: 170; Morrone et al., 2017: 278.
Área de las Tierras Bajas del Pacífico y la Cuenca del Balsas (en parte) Flores-Villela y Goyenechea, 2001: 174.
Esta provincia se ubica en el centro de México, en los estados de Guerrero, México, Jalisco, Michoacán, Morelos, Oaxaca y Puebla, por debajo de los 2,000 m de altitud (Morrone, 2017a). Corresponde a la Cuenca del río Balsas, que se sitúa entre la Faja Volcánica Transmexicana y la Sierra Madre del Sur.
La vegetación consiste en bosques estacionalmente secos y pastizales (Dinerstein et al., 1995). Entre los géneros dominantes de plantas se encuentran Backenbergia, Bursera, Castela, Hymenostephium y Lopezia (Rzedowski, 1978). La diversificación de Bursera (Burseraceae) en esta provincia es muy importante.
Algunos taxones endémicos de esta provincia son: 1 helecho (Notholaena lemmonii var. australis), 1 cacto (Coryphantha bummama), el cuajiote amarillo (Bursera bolivarii), 2 copales (Bursera longipes y B. submoniliformis), 1 escarabajo (Cotinis pueblensis), el cuinique (Spermophilus adocetus), el zacatonero pecho negro (Peucaea humeralis) y el tecolote del Balsas (Megascops seductus).
Regionalización biogeográfica. Dentro de esta provincia se han identificado 2 distritos (Morrone, 2017a): Balsas Inferior y Balsas Superior. La distinción entre ambas áreas ha sido corroborada por varios estudios (e.g., Espinosa-Organista et al., 2008; Gámez et al., 2014; Smith, 1941) y su separación ha sido identificada con una lengüeta de esquistos ocasionada por la Sierra de TaxcoTeloloapan (Sotuyo et al., 2010). El análisis de Gámez et al. (2014), sin embargo, halló una mayor relación entre el distrito del Balsas Inferior con la provincia de las Tierras Bajas del Pacífico y entre el distrito del Balsas Superior con el distrito Orizaba-Zempoaltepec de la Faja Volcánica Transmexicana.
Distrito del Balsas Inferior
Provincia del Balsas Inferior Smith, 1941: 110.
Subprovincia Occidental Ferrusquía-Villafranca, 1990: mapa.
Provincia de la Cuenca del Balsas, O Escalante et al., 1998: 284.
Distrito del Balsas Inferior Morrone, 2014a: 48.
Corresponde a la parte occidental de la provincia.
Distrito del Balsas Superior
Provincia del Balsas Superior Smith, 1941: 110.
Área de la Cuenca del Balsas-Tepalcatepec West, 1964: 368.
Subprovincia del Balsas Ferrusquía-Villafranca, 1990: mapa.
Provincia de la Cuenca del Balsas, E Escalante et al., 1998: 284.
Distrito del Balsas Superior Morrone, 2014a: 48. Corresponde a la parte oriental de la provincia.
Castro-Torreblanca et al. (2014) analizaron los patrones de distribución de las aves de la Cuenca del Balsas, con el objetivo de reconocer unidades dentro de esta provincia biogeográfica. Los autores obtuvieron registros a partir del Atlas de las Aves de México (Navarro-Sigüenza et al., 2003), considerando tanto las especies residentes como las migratorias. Utilizaron celdas de 0.5° de latitud por 0.5° de longitud superpuestas al polígono de la Cuenca del Balsas. A partir de una matriz de datos de presencia-ausencia, llevaron a cabo un análisis de biogeografía numérica o fenética, mediante coeficientes de similitud. El patrón de distribución de la riqueza para la avifauna total y la residente fue semejante. El patrón de diversidad permitió identificar un conjunto avifaunístico característico de la provincia y la existencia de 2 grupos menores de celdas, uno situado al este y otro al oeste. De acuerdo con Castro-Torreblanca et al. (2014), es posible reconocer un área oriental y otra occidental, cada una con sus especies características. En el área oriental, donde se registran las altitudes mayores, hay mayor proporción de especies de climas templados, mientras que en la occidental, que está en contacto con la provincia de las Tierras Bajas del Pacífico, hay una mayor proporción de especies tropicales. Los autores consideraron que estos patrones, sumados a la evidencia de otros taxones, permitirían reconocer 2 distritos biogeográficos.
Evolución biótica. Esta provincia es claramente neotropical, aunque Halffter (1964) consideró que en ella coexisten especies de insectos de filiación neotropical junto con otras del Desierto Chihuahuense y de la Faja Volcánica Transmexicana. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia de la Cuenca del Balsas se relaciona estrechamente con las provincias de las Tierras Bajas del Pacífico, Faja Volcánica Transmexicana y Sierra Madre del Sur. Un análisis de trazos basado en especies de coleópteros (Morrone y Márquez, 2001) mostró que esta provincia se relaciona con las provincias de la Sierra Madre Occidental, Sierra Madre del Sur, Sierra Madre Oriental y Faja Volcánica Transmexicana.
Conservación. La provincia de la Cuenca del Balsas se halla amenazada por la conversión de hábitats naturales para el cultivo intensivo de frutales y ganadería, y la polución asociada con estas actividades (Dinerstein et al., 1995). La ganadería extensiva de cabras, la extracción de leña y la construcción de caminos son las amenazas principales en la Sierra de Huautla, Morelos (Maass et al., 2010). En esta provincia se incluye la siguiente área natural protegida (Conanp, 2018):
Reserva de la Biosfera Sierra de Huautla (Morelos). Posee bosques de encino, selva caducifolia y vegetación inducida. Algunas especies representativas son: el copal (Bursera copallifera), el cuachalalate (Amphipterygium adstringens), el tecolhuixtle (Mimosa benthamii), el tlahuitol (Lysiloma divaricatum), el amate (Ficus petiolaris), una mariposa (Baronia brevicornis), la iguana negra (Ctenosaura pectinata), la chachalaca pálida (Ortalis poliocephala) y el trogón elegante (Trogon elegans).
Provincia Veracruzana
Provincia Veracruzana Smith, 1941: 110; Stuart, 1964: 350; Casas-Andreu y Reyna-Trujillo, 1990: mapa; Morrone, 2014a: 49, 2017a: 173; Morrone et al., 2017: 278.
Provincia de Veracruz Goldman y Moore, 1945: 356.
Provincia Veracrucense Barrera, 1962: 101.
Provincia de Lempira-Tegucigalpa Ryan, 1963: 25.
Región de las Tierras Bajas del Caribe-Golfo (en parte) West, 1964: 368.
Subregión de las Tierras Bajas del Este Savage, 1966: 736.
Provincia Xerófila Mexicana (en parte) Cabrera y Willink, 1973: 34.
Centro del Bosque Lluvioso Centroamericano Müller, 1973: 10.
Provincia Campechana (en parte) Udvardy, 1975: 41.
Provincia de la Costa del Golfo de México Rzedowski, 1978: 109.
Provincia del Golfo de México Samek et al., 1988: 28; Rzedowski y Reyna-Trujillo, 1990: mapa; Arriaga et al., 1997: 64; Morrone, 2001: 42, 2005: 240; Espinosa-Organista et al., 2008: 61.
Provincia Veracruzense Ferrusquía-Villafranca, 1990: mapa.
Provincia del Golfo Ramírez-Pulido y Castro-Campillo, 1990: mapa.
Ecorregión de los Pastizales del Oriente de México Dinerstein et al., 1995: 104.
Provincia de la Costa Este, M Escalante et al., 1998: 284.
Ecorregión del Grijalva-Usumacinta Abell et al., 2008: 408.
Esta provincia se ubica en la costa del Golfo de México, en el este de México (estados de Veracruz, Tabasco, Tamaulipas, San Luis Potosí, Hidalgo, Puebla, Oaxaca, Chiapas y Campeche), Belice y norte de Guatemala (Morrone, 2017a). Es una provincia muy húmeda, generalmente recibe más de 1,000 mm anuales de lluvia.
La vegetación consiste en bosques tropicales húmedos, además de bosques de encinos y de coníferas (donde predomina Pinus caribaea), sabanas y palmares (Dinerstein et al., 1995). Entre los géneros dominantes de plantas se encuentran Dialium, Pimenta, Scheelea y Vochysia (Rzedowski, 1978). Junto con la provincia de la Península de Yucatán, esta provincia alberga la mayor concentración de bosques tropicales húmedos o lluviosos del país, caracterizados por la cantidad de lluvia que reciben anualmente y la temperatura alta, entre otras características (Corlett y Primack, 2011).
Algunos taxones endémicos de esta provincia son la mariposa Troilides torquatus tolus, el escarabajo Cotinis punctatostriata, las lagartijas Sceloporus serripes plioporus y S. variabilis variabilis, y el conejo tapetí (Sylvilagus brasiliensis truei).
Regionalización biogeográfica. Dentro de esta provincia se han identificado 4 distritos (Morrone, 2017a): Bosque Deciduo del Norte de Veracruz, Los Tuxtlas, Valle de Chiapas y Bosque Lluvioso del Sur de Veracruz-Tabasco.
Distrito del Bosque Deciduo del Norte de Veracruz
Área del Bosque Deciduo del Norte de Veracruz West, 1964: 368.
Ecorregión de los Bosques Secos de Tamaulipas y Veracruz Dinerstein et al., 1995: 99.
Ecorregión de los Bosques de Pino y Roble de Veracruz Dinerstein et al., 1995: 102.
Ecorregión de los Bosques Montanos de Veracruz Dinerstein et al., 1995: 103.
Ecorregión de los Bosques Secos de Veracruz Dinerstein et al., 1995: 99.
Ecorregión de las Sabanas de Palmas de Veracruz Dinerstein et al., 1995: 106.
Distrito del Bosque Deciduo del Norte de Veracruz Morrone, 2014a: 51.
En la porción más septentrional de la provincia.
Distrito de Los Tuxtlas
Área de Los Tuxtlas West, 1964: 368.
Subprovincia Tuxtleña Espinosa-Organista et al., 2008: 60.
Provincia de la Sierra de Los Tuxtlas Escalante et al., 1998: 284.
Distrito de Los Tuxtlas Morrone, 2014a: 51.
Corresponde al área de Los Tuxtlas, en el sur de Veracruz. Villaseñor et al. (2018) prroporcionaron una lists de 53 especies de plantas endémicas de este distrito.
Distrito del Valle de Chiapas
Área del Valle de Chiapas West, 1964: 368.
Provincia de las Tierras Bajas de Chiapas C Escalante et al., 1998: 284.
Provincia de las Tierras Bajas de Chiapas S Escalante et al., 1998: 284.
Distrito del Valle de Chiapas Morrone, 2014a: 51.
En el valle enmarcado por las tierras altas de Chiapas.
Distrito del Bosque Lluvioso del Sur de Veracruz-Tabasco
Provincia Tabasqueño-Palencana Barrera, 1962: 101.
Área del Bosque Lluvioso del Sur de Veracruz-Tabasco West, 1964: 368.
Subprovincia Planiciense Ferrusquía-Villafranca, 1990: mapa.
Ecorregión de los Bosques de Pino de Belice Dinerstein et al., 1995: 97.
Ecorregión de los Bosques Húmedos de Tehuantepec Dinerstein et al., 1995: 91.
Ecorregión de las Sabanas de Tabasco y Veracruz Dinerstein et al., 1995: 104.
Distrito del Bosque Lluvioso del Sur de VeracruzTabasco Morrone, 2014a: 51.
En el sur del estado de Veracruz, Tabasco y Belice. La obra de Bueno et al. (2005) compila la información florística y faunística del estado de Tabasco, cuya mayor parte se asigna a este distrito.
Evolución biótica. Si bien esta provincia es netamente neotropical, Rzedowski (1978) consideró que en las porciones secas hay mayor influencia de la flora de la provincia de las Tierras Bajas del Pacífico y de la región Neártica. Wendt (1998) analizó la flora, concluyendo que ésta incluía un elemento antiguo descendiente de linajes laurásicos presentes en América del Norte entre el Cretácico y el Eoceno; y un elemento más reciente descendiente de linajes gondwánicos llegados desde América del Sur en el Neógeno. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia Veracruzana se relaciona estrechamente con la provincia de las Tierras Altas de Chiapas. Un análisis de trazos basado en especies de coleópteros (Morrone y Márquez, 2001) mostró que esta provincia se relaciona con las provincias PuntarenasChiriquí, de las Tierras Bajas del Pacífico y de las Tierras Altas de Chiapas.
Conservación. La provincia Veracruzana se halla amenazada por la explotación petrolera, la construcción de vías, la expansión de plantaciones de cítricos y banano, la explotación de la vida silvestre, la recolección de leña, los fuegos antropogénicos, el uso turístico, la caza excesiva y los ranchos ganaderos (Dinerstein et al., 1995). En esta provincia se incluyen varias áreas naturales protegidas (Conanp, 2018):
Reserva de la Biosfera Los Tuxtlas (Veracruz). Representa el relicto de selva tropical húmeda situado más al norte. Posee bosques de coníferas, bosques de encino, bosques mesófilos de montaña, selva perennifolia, vegetación hidrófila, vegetación inducida y manglar. La obra de González-Soriano et al. (1997) compendia el conocimiento de la flora y fauna de esta reserva, mientras que el libro de Guevara et al. (2006) analiza la ecología del paisaje de esta reserva, con capítulos referidos a la topografía, población humana, geomorfología, deforestación, suelo, clima y vegetación. Algunas especies endémicas son una cícada (Ceratozamia miqueliana), un cedro (Cedrela odorata), la palma cola de pescado (Chamaedorea ernesti-augusti), el mangle rojo (Rhizophora mangle), el ocote (Pinus oocarpa), la paloma tuxtleca (Zentrygon carrikeri), la tucaneta verde (Aulacorhynchus prasinus), el tepezcuintle (Cuniculus paca), el tigrillo (Leopardus wiedii) y el mono aullador (Alouatta palliata).
Reserva de la Biosfera Pantanos de Centla (Tabasco). Llanura de inundación que aporta nutrientes a las pesquerías del Golfo de México, debido al arrastre de materia orgánica proveniente de las partes altas de los ríos Grijalva y Usumacinta. Incluye amplias zonas de refugio, alimentación y de reproducción para aves migratorias y residentes; así como humedales interiores que sirven como refugio y zona de alimentación a diversas especies de importancia comercial, como peces y cangrejos.
Área de Protección de Flora y Fauna Laguna de Términos (Campeche): es el sistema lagunar estuarino de mayor volumen y extensión del país. La región se incluye en la principal ruta migratoria de aves acuáticas norteamericanas, que se conoce como Ruta del Golfo de México o Ruta del Mississippi. Los manglares cubren más de 259,000 ha de costa y junto con los de Tabasco, forman una unidad ecológica costera que es considerada por su productividad natural y biodiversidad, como la más importante en Mesoamérica. Villalobos-Zapata et al. (1999) han propuesto pautas para el manejo de los manglares de esta área.
Provincia de la Península de Yucatán
Provincia Yucateca Smith, 1941: 110; Udvardy, 1975: 41; Espinosa-Organista et al., 2008: 62.
Provincia de la Península de Yucatán Goldman y Moore, 1945: 360; Ryan, 1963: 22; Stuart, 1964: 350; Rzedowski, 1978: 109; Rzedowski y Reyna-Trujillo, 1990: mapa; Morrone, 2001: 43, 2005: 241, 2014a: 51, 2017a: 175; Morrone et al., 2017: 278.
Provincia Yucatanense Barrera, 1962: 101.
Provincia de Yucatán Ryan, 1963: 22; Casas-Andreu y Reyna-Trujillo, 1990: mapa; Ramírez-Pulido y Castro-Campillo, 1990: mapa; Rzedowski y Reyna-Trujillo, 1990: mapa; Arriaga et al., 1997: 63.
Región de las Tierras Bajas del Caribe-Golfo (en parte) West, 1964: 368.
Provincia Campechana Savage, 1966: 736; Brown et al., 1998: 31.
Centro de Yucatán Müller, 1973: 16.
Ecorregión de Yucatán Abell et al., 2008: 408.
Esta provincia se ubica en la Península de Yucatán, en el sureste de México (estados de Campeche, Yucatán y Quintana Roo) y el norte de Guatemala y Belice, en áreas cuya altitud no supera los 300 m (Morrone, 2017a).
La Península de Yucatán es una de las pocas grandes penínsulas del mundo cuya base se encuentra más al sur que su ápice, lo cual sumado a su relieve plano, acentúa la influencia climática marina y la sitúa en la trayectoria de los vientos y ciclones caribeños. Varios autores han detectado la existencia de patrones de distribución, abundancia y riqueza y su relación con los gradientes latitudinal, altitudinal y de humedad de la península (Maya-Martínez et al., 2011). Su historia geológica es compleja, ligada con la del Caribe y América Central. Durante su historia ha sufrido episodios de regresiones y transgresiones marinas de distinta intensidad, e incluso ha estado sumergida completamente por el mar durante el Cretácico (Suárez-Morales, 2003).
La vegetación consiste en bosques tropicales húmedos, bosques estacionalmente secos, sabanas arbustivas, pastizales inundables y manglares en las ciénagas costeras salobres (Dinerstein et al., 1995). Los distintos tipos de vegetación generalmente se asocian con características específicas del suelo (Barrera, 1962).
Algunos taxones endémicos de esta provincia son el kitam ché (Caesalpinia gaumeri), el carábido Amblygnathus subtinctus, la paloma montaraz (Leptotila jamaicensis), una musaraña (Cryptotis mayensis), el venado temazate café (Mazama pandora) y la ardilla de Yucatán (Sciurus yucatanensis). Durán et al. (1998) reconocieron un total de 168 especies de plantas endémicas de la Península de Yucatán.
Barrera (1962) llevó a cabo un análisis biogeográfico de la Península de Yucatán, especialmente basado en especies de vertebrados, con el objeto de determinar si se justificaba considerarla como una provincia biogeográfica. Luego de presentar algunos antecedentes acerca de su situación geográfica, topografía, hidrografía, paleogeografía, geología, clima, suelos y vegetación, este autor discutió las afinidades florísticas de la península, destacando las relaciones de su flora con la del sureste de México, América Central y las Antillas. Al analizar la fauna de vertebrados, Barrera halló que los peces y anfibios endémicos eran escasos, siendo básicamente su fauna derivada de la de América Central, mientras que la endemicidad de los reptiles alcanzaba un 26%, mostrando las mismas afinidades. En cuanto a los mamíferos, 9 especies (17%) resultaron endémicas y 22 especies (39%) estuvieron representadas por subespecies de distribución más o menos restringida a la península. Barrera (1962) concluyó que debido al efecto de peninsularidad, que determina gradientes para la flora y la fauna, los límites de esta provincia biogeográfica dados por diferentes autores varían de acuerdo con los taxones analizados. De acuerdo con Challenger (1988), 66.7% de las especies de plantas de esta provincia son endémicas de Mesoamérica, 25.6% se encuentran en las Antillas y 5.1% en la cuenca del Amazonas, en América del Sur.
Regionalización biogeográfica. La regionalización biogeográfica de esta provincia ha tenido una historia compleja, con distintas unidades reconocidas de acuerdo con los taxones analizados (Arriaga et al., 1997; Barrera, 1962; Duno-de Stefano et al., 2012; Espadas-Manrique et al., 2003; Ibarra-Manríquez et al., 2002). Dentro de esta provincia se han reconocido 4 distritos (Morrone, 2017a): Norte de Yucatán, Rooeño, Petén y Bosques Pantanosos Beliceños. Los 3 primeros distritos corresponden a nuestro país.
Distrito del Norte de Yucatán
Provincia de Yucatán Barrera, 1962: 79; Escalante et al., 1998: 284; Brown et al., 1998: 31.
Área del Norte de Yucatán West, 1964: 368.
Provincia Yucateca Udvardy, 1975: 41.
Provincia Yucatenense Ferrusquía-Villafranca, 1990: mapa.
Ecorregión de los Bosques Secos de Yucatán Dinerstein et al., 1995: 99.
Ecorregión de los Bosques Húmedos de Yucatán Dinerstein et al., 1995: 91.
Distrito Noroccidental Ibarra-Manríquez et al., 2002: 24.
Área de Yucatán Espadas-Manrique et al., 2003: 324.
Área de la Zona Seca de Yucatán Espadas-Manrique et al., 2003: 324.
Distrito Árido/Seco de Yucatán Ramírez-Barahona et al., 2009: 784.
Distrito Yucatán Duno-de Stefano et al., 2012: 1063. Distrito del Norte de Yucatán Morrone, 2014a: 52.
Se sitúa en la parte norte de la península, y sería el más típico o característico. Su clima es tropical subhúmedo, con una franja semiárida en el noroeste, predominando las selvas bajas caducifolias. Sustentado por la presencia de una enredadera (Dictyanthus aeneus), el sak tojoso (Plagiolophus millspaughi), 2 lagartijas (Sceloporus chrysostictus y S. lundelli), 1 víbora de cascabel (Crotalus durissus tzabcan), 1 ratón (Peromyscus yucatanicus yucatanicus) y 1 conejo (Sylvilagus floridanus yucatanicus).
Distrito Rooeño
Subprovincia Rooeña Ferrusquía-Villafranca, 1990: mapa.
Ecorregión de los Humedales de Quintana Roo Dinerstein et al., 1995: 102.
Provincia de Cozumel-Isla Mujeres Escalante et al., 1998: 284.
Distrito Rooeño Morrone, 2014a: 52.
Se encuentra en el este de la península.
Distrito del Petén
Provincia del Petén Smith, 1941: 110; Stuart, 1964: 350; Escalante et al., 1998: 284; Espinosa-Organista et al., 2008: 62.
Provincia Petenense Barrera, 1962: 101.
Área del Bosque Lluvioso de Petén-Yucatán West, 1964: 368.
Provincia Campechana (en parte) Udvardy, 1975: 41.
Provincia de El Petén Casas-Andreu y Reyna-Trujillo, 1990: mapa; Rzedowski y Reyna-Trujillo, 1990: mapa; Arriaga et al., 1997: 63.
Provincia Campechano-Petenense Ferrusquía-Villafranca, 1990: mapa.
Área de El Petén Espadas-Manrique et al., 2003: 325.
Distrito de El Petén Ramírez-Barahona et al., 2009: 783.
Distrito Petén Duno-de Stefano et al., 2012: 1063.
Distrito del Petén Morrone, 2014a: 52.
En el sur de la península, abarca el área del Petén, en el sur de Campeche y Quintana Roo, noroeste de Guatemala y Belice. Su clima es más húmedo, por lo que predominan las selvas altas perennifolias. Sustentado por la presencia del zapotillo (Pouteria amygdalina), 1 asterácea (Ageratum lundellii) y 1 ratón (Peromyscus yucatanicus badius).
Duno-de Stefano et al. (2012) llevaron a cabo un análisis de los patrones de distribución de las especies de leguminosas de la Península de Yucatán, con el objeto de contribuir a su regionalización y esclarecer sus relaciones biogeográficas. Los autores dividieron la Península de Yucatán en celdas de 0.5° de latitud por 0.5° de longitud, fusionando aquellas con superficies pequeñas (en relación con una gran porción en el mar) con los cuadros adyacentes. Se construyó una matriz de presencia-ausencia, a partir de datos de herbarios y bases de datos, y se aplicó un análisis de clusters y otro de parsimonia de endemismos. Además se llevó a cabo un análisis de trazos incluyendo información de la Península de Yucatán así como de América Central, América del Sur, América del Norte, las Antillas y otras áreas tropicales del mundo. El análisis de clusters separó un grupo de celdas septentrionales de otro con celdas meridionales. Los resultados del análisis de parsimonia de endemismos fueron semejantes. Los trazos individuales mostraron un mayor número de especies compartidas con América Central (32), luego América del Norte (11), América del Sur (8) y Antillas (1). De acuerdo con los resultados del análisis se similitud global, los autores reconocieron 2 unidades, una al norte y otra al sur, que consideraron equivalentes a los distritos de Yucatán y del Petén, respectivamente. El análisis de aves de la península de Cortés-Ramírez et al. (2012) coincide con esta división. En cuanto a las relaciones generales evidenciadas por los trazos, la Península de Yucatán aparece más relacionada con Guatemala y Belice, y luego con América Central, siendo mínima la relación biótica con las Antillas.
Evolución biótica. Las afinidades bióticas más evidentes de esta provincia son con las provincias Veracruzana, de la Cuenca del Balsas y de las Tierras Bajas del Pacífico. Su flora también muestra numerosos elementos antillanos, aunque de acuerdo con Rzedowski (1978), esta influencia sería menor que la de la provincia Veracruzana. Durán et al. (1998) detectaron la presencia de elementos florísticos antillanos, centroamericanos y del sureste de México. Ibarra-Manríquez et al. (2002) hallaron relaciones entre la flora arbórea de la Península de Yucatán con la de Chiapas, Oaxaca y Veracruz, mientras que las relaciones con la flora antillana resultaron poco marcadas. De acuerdo con un análisis de parsimonia de endemismos basado en especies de plantas, insectos y aves (Morrone et al., 1999), la provincia de la Península de Yucatán se relaciona estrechamente con las provincias de las Tierras Altas de Chiapas, Veracruzana, Sierra Madre Oriental, Sierra Madre del Sur, Cuenca del Balsas, Tierras Bajas del Pacífico y Faja Volcánica Transmexicana.
Conservación. La provincia de la Península de Yucatán se halla amenazada por la expansión de los ranchos ganaderos, la extracción forestal, los cultivos de cítricos, el crecimiento poblacional, las presiones de los reasentamientos humanos y el desarrollo turístico (Dinerstein et al., 1995). Los suelos de esta provincia se hallan afectados por la pérdida de nutrientes (Sánchez-Colón et al., 2009). En el estado de Campeche, la vegetación original no se halla tan alterada como en otras partes del país (Flores-Villela y Gerez, 1994).

