











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Chiapas cuenta con una gran riqueza biológica, la cual está siendo fuertemente transformada, por lo que es importante realizar investigaciones para monitorear los recursos biológicos y culturales del lugar. En particular, los hongos son un grupo muy diverso y a pesar de que se han llevado a cabo estudios para conocer su riqueza y distribución, aún es necesario realizar inventarios para incrementar el conocimiento de estos organismos. Muchos ambientes naturales están siendo afectados por distintas causas de disturbio, tanto natural como antropogénico, por ello es importante conocer la diversidad actual, que representa uno de los más importantes medios para restaurar dichos ambientes. Con relación a los hongos, se han citado aproximadamente 850 especies en todo el territorio chiapaneco en aproximadamente 100 trabajos. Pérez-Moreno y Villarreal (1988) realizaron una revisión del estado de conocimiento de los hongos y Mixomicetes registrados para el estado y citaron 265 especies, provenientes de 83 localidades. Andrade-Gallegos y Sánchez-Vázquez (2005) citaron 441 especies de hongos, además, estimaron que puede haber un total de 20,000 en la entidad y señalaron que las regiones de los altos, el centro y la selva son las más estudiadas. Recientemente, Ruan-Soto et al. (2013) sugirieron una riqueza de 49,000 especies para Chiapas y registraron 611, por lo que detectaron un incremento de 170 taxones citados en 8 años. En cuanto a los estudios taxonómicos (listados) no se han publicado muchos, Robles-Porras et al. (2006), quienes enlistan 78 especies de macromicetos de la región de Los Altos de Chiapas, consideraron que de éstos, 18 fueron nuevos registro para el estado. Por otro lado, Chanona et al. (2007) registraron 144 especies de macromicetos procedentes de bosques de Quercus y de liquidámbar en Ocozocoautla de Espinosa. Así mismo, Chanona et al. (2014) enlistaron 12 especies de un parque urbano de la ciudad de Tuxtla Gutiérrez, y López-Guzmán et al. (2017) registraron 34 especies del municipio del Salto de Agua. Es necesario enfatizar los estudios en regiones poco exploradas como la fronteriza, en donde se ubica el Parque Nacional Lagunas de Montebello (PNLM) para el cual, a pesar de su relevancia, hay pocos estudios que registren los hongos que se desarrollan en los distintos ambientes, en particular sobre los hongos ectomicorrizógenos y el potencial que ofrecen para la adecuada salud del bosque y para reforestación. Sólo hay 56 especies citadas en el plan de Manejo del Parque Nacional Lagunas de Montebello (Conanp, 2007).
En el caso de los estudios etnomicológicos, se han desarrollado 27 estudios sobre todo en la región Altos de Chiapas, selva Lacandona y selva Zoque, aportando una buena cantidad de especies a los registros del estado. Por ejemplo, Ruan-Soto et al. (2007) registraron 58 especies con importancia cultural para poblaciones mestizas y lacandonas de la selva, de las cuales, 10 fueron nuevos registros para el estado. Lampman et al. (2007) y Sheppard et al. (2008) registran especies comestibles y medicinales para poblaciones tsotsiles y tzeltales de los Altos de Chiapas. En la zona de influencia del PNLM solamente existe un estudio publicado donde los autores registraron el conocimiento al respecto de 15 especies utilizadas como alimento, medicina y de manera recreativa en la comunidad de San Antonio Lindavista (Grajales-Vásquez et al., 2008).
El estudio y la identificación taxonómica de las especies de hongos ectomicorrizógenos que se desarrollan en ambientes particulares, es básico para entender la actividad de la comunidad forestal y su dinámica. También es necesario para la conservación, debido a que la propagación de las especies representa fuentes de inóculo potencial para el buen funcionamiento y repoblación, o restauración de los bosques durante o después de los disturbios. Muchas de las especies ectomicorrizógenas son fuente de alimento para animales y para distintos grupos humanos, quienes los consumen obteniendo beneficios a la salud. En muchos casos la venta de hongos silvestres micorrizógenos aporta ingresos económicos para el sustento familiar. En el caso de las zonas turísticas, la venta de éstos adquiere un valor agregado, ya que se ofrecen en diversos platillos para consumo, como ocurre en el PNLM.
Materiales y métodos
El Parque Nacional Lagunas de Montebello se ubica en el estado de Chiapas y ocupa parte de los municipios de La Independencia y La Trinitaria, en la frontera con Guatemala. Sus coordenadas extremas son 16°04’20” y 16° 09’38” N, 91° 38’14” y 91° 47’41” O y su altitud oscila entre los 1,200 y 1,800 m snm, con un promedio de 1,500 m. Presenta una superficie de 6,033 hectáreas y el 95% pertenece al municipio de La Trinitaria. La región en donde se encuentra el parque forma parte de la provincia fisiográfica de las sierras de Chiapas y Guatemala. Geológicamente dominada por rocas sedimentarias cretáceas de origen marino que al consolidarse formaron calizas y dolomitas. Los principales elementos del relieve son lomeríos de poca altura con laderas de inclinación variable que pueden formar paredes abruptas en las partes superiores. Estos lomeríos se formaron por plegamientos que en general tienen una orientación paralela en dirección noroeste a suroeste y están separados por depresiones de tamaño variable que en muchos casos están ocupadas por los lagos característicos de la región y de igual forma se presentan numerosas cuevas, grutas y oquedades. Hacia el noroeste en las partes bajas del parque, los lagos son irregulares y elongados y al este, en las partes altas los lagos son más circulares o igualmente irregulares. La mayoría de los suelos en el parque se originaron de rocas calizas y dolomitas, siendo los litosoles y las rendzinas los que ocupan mayor superficie y se encuentran principalmente en las partes altas, mientras que en las partes bajas son remplazados por vertisoles, acrisoles, gleysoles y fluvisoles. Hidrológicamente, la región se encuentra dentro de la gran cuenca del río Grijalva- Usumacinta, específicamente en la subcuenca del río Grande de Comitán que a su vez forma parte de la cuenca del río Lacantún. El clima varía de templado húmedo con lluvias todo el año, C(fm), a semicálido con lluvias en verano A(C)w, con una temperatura media anual de 17-18 °C y una precipitación media anual de 1,800-1,900 mm. La vegetación predominante en el PNLM es el bosque mesófilo de montaña. Ocupa el 50% de la superficie del parque y se ubica principalmente en laderas abruptas, protegidas de la exposición. Una de las características más distintivas de este bosque es la presencia de Liquidambar styraciflua, en codominancia con encinos como Quercus benthamii o Q. laurina y pinos como Pinus oocarpa y P. maximinoi. También son comunes en estos bosques: Podocarpus matudai, Clethra mexicana, Amphitecna silvicola, Dendropanax arboreus, Wimmeria montana, Beilschmieda steyermarkii, Cleyera theaeoides, Rapanea juergensenii, Parathesis chiapensis; géneros como: Persea, Nectandra, Ocotea, Psychotria y helechos arborescentes como Cyathea fulva, entre otros. El bosque de pino-encino se presenta en lugares menos húmedos o más expuestos y ocupa el 20% de la superficie del parque. Este tipo de vegetación a menudo se considera un estado sucesional, producto de la perturbación debido a que comparte más del 80% de las especies que se presentan en el bosque mesófilo de montaña. El resto de la vegetación está representada por elementos riparios, como tulares y carrizales, presente a las orillas de algunas lagunas, vegetación secundaria o acahuales o pastizales inducidos (Conanp, 2007; Mora-Palomino et al., 2017). Esta región fue seleccionada debido a que en la actualidad se carece de un inventario específico sobre los hongos potencialmente ectomicorrizógenos que se desarrollan dentro del parque.
Se realizaron 2 exploraciones por año (2004, 2005 y 2006), con duración de 7 días cada una, durante la época de lluvias (agosto-octubre) para la recolección de los hongos ectomicorrizógenos en el área de estudio. Se usaron las técnicas descritas por Brundrett et al. (1996) para la recolección, etiquetado y herborización de los hongos potencialmente ectomicorrizógenos. Éstas implicaron la selección de 12 sitios de recolección dentro del PNLM: 1) camino a Antelá, límites del PNLM, 2) camino a Pomarosa, 3) Cinco Lagos, 4) grutas de San Rafael, 5) laguna Bosque Azul, 6) laguna Encantada, 7) laguna Ensueño, 8) laguna Escondida, 9) laguna Montebello) 10) laguna Pojoj, 11) laguna San Lorenzo y 12) Tziscao (Fig. 1).
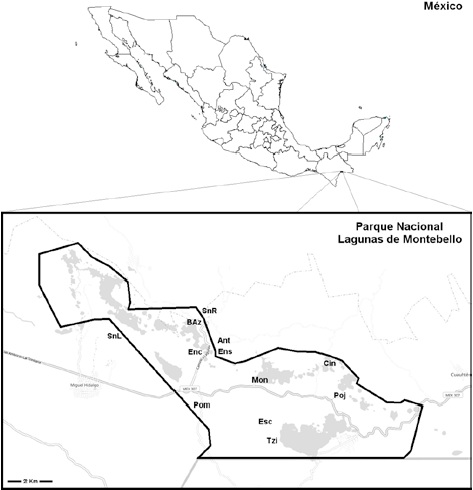
Figura 1 Localización del área de estudio y sitios de recolección de los hongos estudiados: Ant = camino a Antelá, BAz = laguna Bosque Azul, Cin = Cinco Lagos, Enc = laguna Encantada, Ens = laguna Ensueño, Esc = laguna Escondida, Mon = laguna Montebello, Poj = laguna Pojoj, Pom = camino a Pomarosa, SnL = laguna San Lorenzo, SnR = grutas de San Rafael, Tzi = laguna Tziscao.
El muestreo y la recolección de las estructuras reproductivas de los hongos se realizó de manera dirigida, abarcando los principales tipos de vegetación que se encontraron en el área de estudio. Los ejemplares recolectados fueron almacenados en bolsas de papel encerado para protegerlos de la desecación y transportados en canastas de mimbre para evitar su maltrato. Se registró cada espécimen recolectado y a cada uno se asignó un código numérico, incluyendo el nombre del recolector. Se registró el nombre de la localidad, incluyendo coordenadas geográficas, fecha de recolección y detalles sobre el hábitat, por ejemplo, el sustrato, el tipo de vegetación o asociación vegetal y altitud. La caracterización de ejemplares recolectados incluyó información sobre su tamaño, forma, textura, ornamentación, color, etc., poniendo particular atención a las variaciones que se pueden presentar con la edad y en el color con luz de día y con referencia a 2 cartas o tablas de color estandarizadas (Kornerup y Wannscher, 1978; Munsell Color, 1975). Se registraron algunas reacciones que presentan los hongos a ciertos reactivos como el KOH, FeSO4 o reactivo de Melzer. Se tomaron fotografías en fresco, mostrando ejemplares jóvenes y maduros, algunos en sección longitudinal y sobre un fondo de color neutro. Se obtuvo la esporada, colocando un píleo de un ejemplar maduro pero no viejo, sin el estípite, sobre papel blanco, mismo que se cubrió con un recipiente para protegerlo de la desecación. Se dejó por 12-24 h. Posteriormente, se registró el color que presentó el depósito de las esporas. Los ejemplares se deshidrataron utilizando una secadora eléctrica a una temperatura aproximada de 45 °C durante 24 h. Los ejemplares grandes o carnosos se seccionaron por la mitad para facilitar el proceso. Posteriormente, los ejemplares se empacaron en bolsas de papel o plástico, bien etiquetadas y rotuladas, junto con sus descripciones en fresco y notas de campo, para luego ser almacenadas en un lugar seco.
La información etnomicológica relativa a las especies recolectadas se obtuvo a través de entrevistas directas con habitantes de las zonas cercanas a los sitios de recolección. Se solicitó que algunas personas se involucraran en las salidas al campo, tanto para la recolección del material como para la obtención de información sobre usos y nombres de los hongos recolectados y durante todas las salidas se obtuvieron fotografías de los ejemplares en fresco, así como de la manera en que se usan.
Para la determinación taxonómica de los ejemplares recolectados, las técnicas empleadas se describen detalladamente en Largent et al. (1977) y Brundrett et al. (1996). Se realizaron disecciones de las diferentes partes que conforman las estructuras reproductivas de los hongos potencialmente ectomicorrizógenos y se realizaron observaciones microscópicas, utilizando para ello, diversos reactivos, soluciones y medios de montaje (solución acuosa de hidróxido de potasio, hidróxido de amonio, rojo Congo, azul de algodón en ácido láctico y reactivo de Melzer). Se tomaron medidas bajo el microscopio de las diferentes partes que conforman las estructuras reproductivas, incluyendo esporas, basidios, cistidios, pelis, etc., observándose también su forma, ornamentación, coloración, etc. Con esta información y con la descripción macroscópica de los caracteres en fresco, se realizó la determinación taxonómica, utilizando bibliografía especializada o material de referencia previamente identificado. Los ejemplares se depositaron en el Herbario TLXM del Centro de Investigaciones en Ciencias Biológicas de la Universidad Autónoma de Tlaxcala, con algunos duplicados en la colección de hongos de El Colegio de la Frontera Sur, en Tapachula, Chiapas.
Para los análisis de diversidad de los hongos potencialmente ectomicorrizógenos del PNLM, fueron considerados 9 sitios de estudio de los 12 descritos anteriormente. Las lagunas Cinco Lagos, San Lorenzo y Tziscao, no fueron incluidas en los análisis por la escasa representatividad obtenida en estos sitios (pocas recolecciones de hongos).
La abundancia fue estimada por el número de recolecciones y cada recolección representa una unidad de muestreo. Las unidades de muestreo estuvieron separadas una de otra por al menos 5 m. Para estimar la completitud de los inventarios de hongos de cada laguna, así como la completitud total o regional del inventario del PNLM, se asumió como incidencia (presencia-ausencia) el número de unidades de muestreo (recolecciones) de cada especie en cada laguna. El porcentaje de completitud se estimó a través de la cobertura de la muestra (Cm) para datos de incidencia. La Cm calcula la proporción del número de muestras totales en la comunidad que pertenecen a las especies representadas en cada inventario (Chao y Jost, 2012; Chao et al., 2013). El déficit de cobertura (1-Cm) representa la probabilidad de que la próxima muestra recolectada pertenezca a una especie aún no registrada previamente en el inventario (Chao y Jost, 2012).
Debido a la variación de la completitud de los inventarios, la riqueza de especies acumulada en cada laguna se comparó a través de un análisis de rarefacción- extrapolación, estandarizando a 34 unidades de muestreo (el doble de las muestras de la comunidad de hongos con el menor tamaño de la muestra, Pom, n = 17). La interpolación y extrapolación fueron realizadas a 100 aleatorizaciones con intervalos de confianza (IC) a 95% en el programa iNETX versión 1.3.0 (https://chao.shinyapps.io/iNEXTOnline/; Hsieh et al., 2013).
Para el análisis de disimilitud se utilizó el índice beta de Jaccard (1-βJ). Este índice compara la diferencia en composición de especies de 2 comunidades en función de las especies compartidas. Los valores van de 0 a 1 o bien, en términos de porcentaje de 0 a 100, en donde los pares de lagunas que no comparten ninguna especie representan el valor de máxima diferenciación (100%) (Jost et al., 2011).
A través de los índices de diversidad taxonómica se puede incorporar información histórico-evolutiva de una comunidad tanto en las medidas de diversidad alfa como en la diversidad beta, para lo cual se utiliza la clasificación jerárquica linneana por arriba de la categoría de especie. Así, una comunidad tendrá mayor diversidad taxonómica si sus especies pertenecen a distintos géneros, y más aún, si los géneros pertenecen a distintas familias, que si sus especies son cogenéricas y están relacionadas de manera cercana (García-de Jesús et al., 2016). Bajo esta aproximación se calculó la diversidad taxonómica promedio y la disimilitud taxonómica de las comunidades de hongos.
La diversidad taxonómica se midió con el índice de distancia taxonómica promedio (Δ+) entre pares de especies de cada comunidad (Clarke y Warwick, 1998). La distancia taxonómica promedio toma en cuenta la categoría taxonómica en la cual está relacionada cualquier pareja de especies de hongos y mide la separación taxonómica promedio entre 2 especies seleccionadas al azar de la comunidad. La estimación del índice se realizó en el programa PRIMER v5 calculada a partir de 1,000 aleatorizaciones (Clarke y Gorley, 2001).
La disimilitud taxonómica se midió como 1-ΔT, donde ΔT es el índice de similitud taxonómica descrito por Bacaro et al. (2007), que se basa en el índice de Jaccard. Este índice mide la proporción que representan los taxones compartidos del número total de taxones en 2 sitios. Los valores de disimilitud taxonómica (1-ΔT) van de 0, cuando la estructura taxonómica del conjunto de especies de ambas comunidades es idéntica, a 1 cuando la estructura taxonómica de las especies de ambas comunidades es totalmente diferente (Bacaro et al., 2007; García-de Jesús et al., 2016).
Para comparar las 9 comunidades de hongos de acuerdo con las medidas obtenidas de disimilitud de especies (1- βJ) y disimilitud taxonómica (1- ΔT), se realizaron análisis de ordenación no paramétrica NMDS (Nonmetric Multidimensional Scaling) en el programa Past 3.0 (Hammer et al., 2001).
Las categorías taxonómicas que se utilizaron para los análisis de diversidad taxonómica fueron, reino, filo, subfilo, clase, orden, familia, género y especie, siguiendo la propuesta de clasificación de Index Fungorum (http://www.indexfungorum.org, 2017).
La información de los ejemplares identificados a especie se incorporó a la base de datos Biótica, que se elaboró de acuerdo con el instructivo para la conformación de bases de datos de la Conabio.
Resultados
Se recolectaron 671 ejemplares de hongos ectomicorrizógenos que corresponden con 93 especies. La lista de especies encontradas en cada sitio muestreado dentro del parque, se muestran en la Tabla 1.
Tabla 1 Especies de hongos potencialmente ectomicorrizógenos presentes en 12 localidades de muestreo en el Parque Nacional Lagunas de Montebello, Chiapas (2004-2006).
| Taxón | Localidades | |||||||||||
| Ant | Pom | Cin | SnR | BAz | Enc | Ens | Esc | Mon | Poj | SnL | Tzi | |
| Amanita arocheae Tulloss, Ovrebo y Halling | 1 | |||||||||||
| Amanita brunnescens G.F. Atk. | 2 | |||||||||||
| Amanita cokeri (E.-J. Gilbert y Kühner) E.-J. Gilbert | 1 | |||||||||||
| Amanita farinosa Schwein | 1 | 2 | ||||||||||
| Amanita fuligineodisca Tulloss, Ovrebo y Halling | 1 | 1 | ||||||||||
| Amanita multisquamosa Peck | 1 | |||||||||||
| Amanita polypyramis (Berk. y M.A. Curtis) Sacc. | 1 | 4 | ||||||||||
| Amanita novinupta Tulloss y J. Lindgr. | 1 | 1 | 3 | 4 | 7 | 1 | ||||||
| Amanita aff. virosa (Fr.) Bertillon | 3 | |||||||||||
| Amanita volvata (Peck) Lloyd | 1 | |||||||||||
| Amanita xylinivolva Tulloss, Ovrebo y Halling | 1 | 2 | ||||||||||
| Boletellus chrysenteroides (Snell) Snell | 3 | 1 | 2 | 2 | ||||||||
| Boletellus jalapensis (Murrill) E.-J. Gilbert | 5 | 9 | 3 | |||||||||
| Cantharellus lateritius (Berk.) Singer | 2 | |||||||||||
| Cantharellus minor Peck | 4 | 2 | 1 | 4 | 5 | 2 | 7 | |||||
| Coltricia cinnamomea (Jacq.) Murrill | 1 | 2 | 1 | 2 | 1 | 2 | 10 | 4 | 2 | |||
| Cortinarius pholideus (Lilj.) Fr. | 2 | |||||||||||
| Cortinarius violaceus (L.) Gray | 1 | 1 | 1 | |||||||||
| Craterellus ignicolor (R.H. Petersen) Dahlman, Danell y Spatafora | 10 | 2 | 12 | 2 | 1 | 7 | 1 | |||||
| Craterellus fallax A.H. Sm. | 1 | 1 | 3 | 4 | 1 | 2 | ||||||
| Craterellus lutescens (Pers.) Fr. | 2 | 1 | 4 | 5 | 8 | 7 | 11 | 1 | ||||
| Geoglossum simile Peck | 1 | 3 | 1 | 9 | ||||||||
| Heimioporus betula (Schwein.) E. Horak | 1 | 1 | 3 | |||||||||
| Helvella crispa Bull. | 1 | 4 | 1 | 1 | 4 | |||||||
| Helvella macropus (Pers.) P. Karst. | 1 | 1 | 1 | 1 | 2 | |||||||
| Helvella palustris Peck | 1 | 6 | ||||||||||
| Humaria hemisphaerica (Hoffm.) Fuckel | 1 | |||||||||||
| Hydnum albidum Peck | 1 | |||||||||||
| Hydnum repandum L. | 1 | 1 | 2 | 3 | 1 | |||||||
| Hydnum umbilicatum Peck | 1 | 4 | 2 | |||||||||
| Hygrocybe cantharellus (Schwein.) Murril | 1 | 1 | 2 | |||||||||
| Hygrocybe conica (Schaeff.) P. Kumm. | 3 | 2 | 2 | 1 | 1 | |||||||
| Hygrocybe miniata (Fr.) P. Kumm. | 4 | 1 | 1 | 2 | 7 | 7 | ||||||
| Hygrophorus russula (Schaeff.) Kauffman | 1 | |||||||||||
| Inocephalus murrayi (Berk. y M.A. Curtis) Rutter y Watling | 5 | 1 | 1 | 5 | 6 | |||||||
| Inocybe cf. calamistrata (Fr.) Gillet | 1 | |||||||||||
| Inocybe dulcamara (Pers.) P. Kumm. | 1 | 2 | 5 | |||||||||
| Laccaria amethystina Cooke | 2 | 3 | 16 | 10 | 19 | |||||||
| Laccaria laccata (Scop.) Cooke | 1 | 1 | ||||||||||
| Laccaria striatula (Peck) Peck | 5 | 5 | 4 | |||||||||
| Laccaria trichodermophora G.M. Muell. | 1 | 2 | 1 | 1 | ||||||||
| Lactarius chrysorrheus Fr. | 2 | 7 | 2 | 1 | 1 | 1 | 1 | |||||
| Lactarius costaricensis Singer | 6 | 5 | ||||||||||
| Lactarius haugiae Bandala, Montoya y Ramos | 1 | 1 | 1 | 3 | ||||||||
| Lactarius indigo (Schwein.) Fr. | 16 | 1 | ||||||||||
| Lactarius miniatosporus Montoya y Bandala | 1 | 3 | 3 | |||||||||
| Lactarius smithii Montoya y Bandala | 1 | 1 | 7 | 2 | ||||||||
| Lactarius subplinthogalus Coker | 1 | |||||||||||
| Lactifluus corrugis (Peck) Kuntze | 1 | |||||||||||
| Lactifluus deceptivus (Peck) Kuntze | 1 | |||||||||||
| Lactifluus gerardii (Peck) Kuntze | 1 | 3 | ||||||||||
| Lactifluus volemus (Fr.) Kuntze | 1 | |||||||||||
| Leccinum cartagoense (Wolfe y Bougher) Halling y G.M. Muell. | 2 | |||||||||||
| Leotia lubrica (Scop.) Pers. | 1 | 3 | 1 | 6 | ||||||||
| Multifurca furcata (Coker) Buyck y V. Hofstetter | 1 | |||||||||||
| Neoboletus pseudosulphureus (Kallenb.) Klofac | 1 | 1 | 1 | |||||||||
| Phellodon melaleucus (Sw. ex Fr.) P. Karst. | 1 | |||||||||||
| Phellodon niger (Fr.) P. Karst. | 2 | 1 | ||||||||||
| Phylloporus centroamericanus Singer y L.D. Gómez | ||||||||||||
| Phylloporus phaeoxanthus Singer y L.D. Gómez | 1 | 3 | 1 | |||||||||
| Pisolithus arhizus (Scop.) Rauschert | 2 | |||||||||||
| Pseudocraterellus calyculus (Berk. y M.A. Curtis) D.A. Reid | 2 | 1 | 2 | 1 | ||||||||
| Pulveroboletus ravenelii (Berk. y M.A. Curtis) Murrill | 2 | 3 | 3 | |||||||||
| Phaeoclavulina cyanocephala (Berk. y M.A. Curtis) Giachini | 1 | |||||||||||
| Ramaria cystidiophora var. citronella (Kauffman) Corner Marr y D.E. | 1 | |||||||||||
| Stuntz | ||||||||||||
| Ramaria fennica var. griseolilacina (P. Karst.) Ricken Schild | 1 | |||||||||||
| Retiboletus griseus (Frost) Manfr. Binder y Bresinsky | 2 | 1 | ||||||||||
| Russula acrifolia Romagn. | 4 | |||||||||||
| Russula cf. amoenolens Romagn. | 1 | 1 | 1 | |||||||||
| Russula austromontana Singer | 1 | |||||||||||
| Russula cf. cyanoxantha (Schaeff.) Fr. | 1 | 1 | ||||||||||
| Russula delica var. centroamericana Singer | 2 | |||||||||||
| Russula densifolia Secr. ex Gillet. | 1 | |||||||||||
| Russula foetentula Peck | 1 | 1 | 1 | 1 | ||||||||
| Russula laurocerasi Melzer | 1 | 3 | ||||||||||
| Russula mexicana Burl. | 8 | 2 | 3 | 1 | 2 | |||||||
| Russula michiganensis Shaffer | 1 | |||||||||||
| Russula modesta Peck | 1 | 2 | 1 | |||||||||
| Russula pectinatoides Peck | 1 | 3 | ||||||||||
| Russula pulverulenta Peck | 1 | |||||||||||
| Russula silvicola Shaffer | 2 | 1 | 2 | 1 | 8 | |||||||
| Russula subobscura Murrill | 2 | |||||||||||
| Russula vesicatoria Burl | 4 | 1 | ||||||||||
| Scleroderma areolatum Ehrenb. | 2 | 2 | ||||||||||
| Scleroderma cepa Pers. | 1 | 2 | ||||||||||
| Strobilomyces confusus Singer | 1 | 1 | 1 | |||||||||
| Suillus decipiens (Peck) Kuntze | 5 | 8 | 1 | 15 | 6 | 3 | 1 | |||||
| Suillus hirtellus (Peck) Snell | 1 | 2 | 1 | |||||||||
| Suillus subvariegatus Snell y E.A. Dick | 1 | 1 | ||||||||||
| Sutorius eximius (Peck) Halling, M. Nuhn y Osmundson | 3 | 1 | ||||||||||
| Tricholoma pardinum (Pers.) Quél. | 1 | |||||||||||
| Turbinellus floccosus (Schwein.) Earle | 5 | 4 | 2 | 1 | 4 | 2 | ||||||
| Xanthoporus peckianus (Cooke) Audet | 1 | |||||||||||
| Total (especies) | 27 | 12 | 5 | 19 | 37 | 38 | 35 | 13 | 44 | 18 | 4 | 4 |
Ant = camino a Antelá; Pom = camino a Pomarosa; Cin = Cinco Lagos; SnR = grutas de San Rafael; BAz = laguna Bosque Azul; Enc = laguna Encantada; Ens = laguna Ensueño; Esc = laguna Escondida; Mon = laguna Montebello; Poj = laguna Pojoj; SnL = laguna San Lorenzo; Tzi = laguna Tziscao.
El número de especies de hongos ectomicorrizógenos presentes en el PNLM seguramente es mayor, ya que algunos géneros como Boletus sensu lato, Cortinarius, Hydnellum, Inocybe y Phellodon no se revisaron con detalle. La localidad más rica en especies fue el sitio aledaño a la Laguna de Montebello con 44, seguido del sitio que rodea a la laguna Encantada con 38 y el cercano a la laguna Bosque Azul con 37, además del sitio que circunda la laguna Ensueño con 35. Las localidades con menor número de especies fueron laguna San Lorenzo, Cinco Lagos y Tziscao con 5, 6 y 4 especies, respectivamente, las cuales no fueron incluidas en el análisis de diversidad.
Las especies identificadas pertenecen a 38 géneros, 4 de la clase Ascomycetes y 34 de la clase Basidiomycetes. El género con mayor número de especies fue Russula con 16, seguido de Amanita con 11 y Lactarius con 7. Al menos 10 especies fueron registradas de la mitad de las localidades: Amanita novinupta, Craterellus ignicolor, Craterellus lutescens, Cantharellus minor, Coltricia cinnamomea, Craterellus fallax, Turbinellus floccosus, Hygrocybe miniata, Lactarius chrysorrheus y Suillus decipiens. Estas especies fueron las más representativas tomando en cuenta el número de bosques aledaños a las lagunas del PNLM, en que se encontraron y porque fueron las que presentaron la mayor frecuencia. Treinta y siete especies, que representan el 39.8% de los taxones registrados fueron encontradas en una sola localidad, muchas veces con una sola recolección. Ocho especies, Amanita fuligineodisca, Cortinarius pholideus, Leccinum cartagoense, Lactarius costaricensis, Pseudocraterellus calyculus, Russula austromontana, Russula pulverulenta y Russula versicatoria son nuevos registros para la República Mexicana (Fig. 2).

Figura 2 Especies de hongos potencialmente ectomicorrizógenos que se registran por primera vez para la República Mexicana y que se recolectaron en el Parque Nacional Lagunas de Montebello, Chiapas: A = Amanita fuligineodisca, B = Cortinarius pholideus, C = Leccinum cartagoense, D = Lactarius costaricensis, E = Pseudocraterellus calyculus, F = Russula austromontana, G = Russula pulverulenta, H = Russula versicatoria.
Considerando la abundancia registrada, las especies más representativas de la comunidad de hongos ectomicorrizógenos fueron: Laccaria amethystina con el 7.41% de abundancia relativa, Craterellus lutescens y Suillus decipiens con 5.78%, Craterellus ignicolor con 5.19%, Cantharellus minor y Coltricia cinnamomea con 3.7% (Fig. 3).

Figura 3 Hongos con valores altos de abundancia en las zonas de estudio: A = Laccaria amethystina, B = Craterellus lutescens, C = Suillus decipiens, D = Craterellus ignicolor, E = Cantharellus minor, F = Coltricia cinnamomea.
La completitud de los inventarios de hongos de las 9 lagunas fue del 55 al 91%. Así, el inventario de hongos más completo y con el menor déficit de muestreo correspondió con la laguna Montebello (Mon) con el 91%. Los inventarios con el porcentaje de completitud más baja fueron los sitios en Antelá (Ant) y Pomarrosa (Pom) con el 66 y 55%, respectivamente. La completitud del inventario regional del PNLM fue de 97% (Tabla 2).
Tabla 2 Valores obtenidos en el Análisis de Completitud, con base en incidencia de hongos ectomicorrizógenos en los bosques aledaños a las Lagunas de Montebello, Chiapas, PNLM.
| Laguna | Unidades de muestreo |
Especies únicas |
Especies duplicadas |
Riqueza específica |
Cm | Déficit de muestreo |
| Pom | 17 | 8 | 3 | 12 | 0.55 | 0.45 |
| Ant | 53 | 18 | 2 | 27 | 0.66 | 0.34 |
| Esc | 32 | 9 | 1 | 13 | 0.72 | 0.28 |
| Poj | 41 | 9 | 5 | 18 | 0.79 | 0.21 |
| SnR | 49 | 10 | 3 | 19 | 0.80 | 0.20 |
| Ens | 91 | 16 | 8 | 35 | 0.83 | 0.17 |
| Enc | 120 | 19 | 5 | 38 | 0.84 | 0.16 |
| BAz | 94 | 14 | 11 | 37 | 0.85 | 0.15 |
| Mon | 159 | 14 | 10 | 44 | 0.91 | 0.09 |
| PNLM | 656 | 23 | 11 | 90 | 0.97 | 0.03 |
Valores de completitud para cada inventario (Cm: cobertura de la muestra) con base en datos de incidencia (número de muestras), especies únicas (especies representadas por una sola muestra) y especies duplicadas (especies representadas por dos muestras), y los valores del déficit de muestreo para cada laguna y a nivel regional Parque Nacional Lagunas de Montebello. Ant = camino a Antelá; Pom = camino a Pomarosa; Esc = laguna Escondida; Poj = laguna Pojoj; SnR = grutas de San Rafael; Ens = laguna Ensueño; Enc = laguna Encantada; BAz = laguna Bosque Azul; Mon = laguna Montebello; PNLM= Parque Nacional Laguna de Montebello.
La mayor riqueza de hongos estimada fue para las lagunas Bosque Azul y Montebello (Baz = 22 y Mon = 21 especies, respectivamente), en comparación con las lagunas San Rafael (SnR = 16 especies) y laguna Escondida (Esc = 14 especies). No obstante, la riqueza estimada de hongos en los 9 sitios fue muy similar sin diferencias estadísticamente significativas con intervalos de confianza a 95% (Fig. 4).
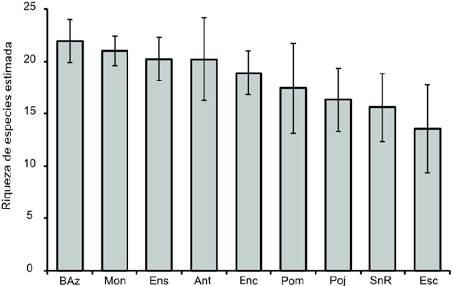
Figura 4 Riqueza de especies de hongos estandarizada a un mismo tamaño de muestra (n = 34) con intervalos de confianza de 95% a 100 aleatorizaciones. Ant = camino a Antelá; Pom = camino a Pomarosa; Esc = laguna Escondida; Poj = laguna Pojoj; SnR = grutas de San Rafael; Ens = laguna Ensueño; Enc = laguna Encantada; BAz = laguna Bosque Azul; Mon = laguna Montebello.
El promedio de diversidad taxonómica de hongos esperado por azar en el PNLM fue de Δ+ = 49.9, sin diferencias significativas entre los 9 sitios. Sin embargo, con referencia a este promedio se pueden identificar 3 conjuntos de sitios con valores de diversidad taxonómica diferentes: laguna Escondida (Esc, Δ+ = 57.4), grutas de San Rafael (SnR, Δ+ = 57.5) y laguna Pojoj (Poj, Δ+ = 55), que presentaron los valores de diversidad taxonómica promedio más altos, contrario a camino a Pomarosa (Pom, Δ+ = 45.6) y camino Antelá (Ant, Δ+ = 45.5), cuyos valores fueron los más bajos. El resto de los sitios registraron valores de diversidad taxonómica cercanos al promedio entre Δ+ = 53 y 49.
Al comparar la riqueza de especies y la diversidad taxonómica de hongos del PNLM se observó que, a nivel de riqueza de especies, no hay una diferenciación marcada de la riqueza entre sitios, contrario con la diversidad taxonómica, donde se logran diferenciar conjuntos de sitios con valores diferentes en torno al promedio esperado por azar (Fig. 5).
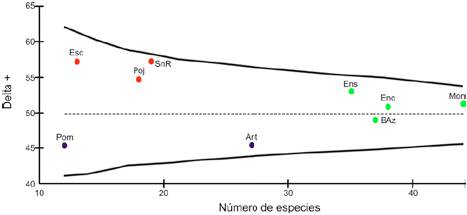
Figura 5 Diversidad taxonómica de hongos en El Parque Nacional Lagunas de Montebello, la línea discontinua es el valor de la diversidad taxonómica promedio esperado por azar. Los colores indican conjunto de sitios con diferentes valores de diversidad taxonómica. Ant = camino a Antelá; Pom = camino a Pomarosa; Esc = laguna Escondida; Poj = laguna Pojoj; SnR = grutas de San Rafael; Ens = laguna Ensueño; Enc = laguna Encantada; BAz = laguna Bosque Azul; Mon = laguna Montebello.
La disimilitud de especies promedio (1-βJ) entre pares de comunidades de hongos fue de 64%. Los sitios con mayor diferenciación en la composición de especies son las lagunas Camino a Pomarosa (Pom), laguna Pojoj (Poj) y camino a Antelá (Ant), con valores de disimilitud de 89 a 93%. Mientras que los sitios con la menor diferenciación de especies (62%) fueron: laguna Montebello (Mon), bosque Azul (Baz), laguna Ensueño (Ens) y laguna Encantada (Enc) (Fig. 6A).
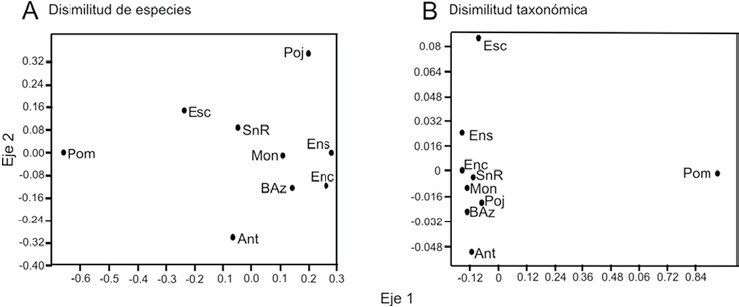
Figura 6 Ordenación no paramétrica multidimensional (NMDS) de las comunidades de hongos en El Parque Nacional Lagunas de Montebello. A, con base en la disimilitud de especies (1-βJ) y B, con base en la disimilitud taxonómica (1-ΔT).
La disimilitud taxonómica promedio de hongos en el área de estudio fue de 22.1%. Los sitios camino a Pomarosa (Pom), laguna Pojoj (Poj) y laguna Escondida (Esc) fueron los sitios con mayor diferenciación en composición taxonómica en un 72.8%. En el resto de los sitios la disimilitud taxonómica fue similar entre un 46% y un 65% (Fig. 6B).
Al comparar la disimilitud de especies y la disimilitud taxonómica, se puede observar que las lagunas camino a Pomarosa (Pom) y camino a Antelá (Ant) son sitios que se diferencian del resto tanto a nivel de especies como en estructura taxonómica, mientras que la comunidad de hongos de laguna Encantada (Enc) y laguna Ensueño (Ens) son comunidades muy similares a nivel de especies y a nivel de estructura taxonómica.
Los sitios laguna Encantada y laguna Ensueño comparten 20 taxones, es decir, el 54.1 y 57.1%, respectivamente, del total por localidad. Los sitios laguna Bosque Azul y laguna Montebello comparten 22 taxones, que representan el 59.5 y 50%, respectivamente, de los taxones por localidad. Estas 4 localidades en conjunto forman el grupo con menor diferenciación, compartiendo entre ellas 9 especies: Boletellus chrysenteroides, Craterellus ignicolor, Craterellus lutescens, Cantharellus minor, Coltricia cinnamomea, Helvella crispa, Lactarius chrysorrheus, Russula mexicana y Suillus decipiens. De éstas, Boletellus chrysenteroides no se encontró en ningún otro sitio. La riqueza es similar en las 9 lagunas; sin embargo, tienen una composición propia (60%). Esto es importante para la conservación porque a nivel beta, se observa una riqueza diferente en las Lagunas Pojoj, Pomarrosa, Antelá y Escondida. Es decir, a nivel de paisaje tienen una riqueza similar, sin embargo, cada laguna aporta diferentes especies al ensamble total. Esta información muestra la importancia de conservar cada una de las lagunas del parque.
Los sitios laguna Encantada y laguna Ensueño se ubican muy cerca una de otra, a menos de 200 m y comparten el mismo tipo de vegetación, con dominancia de Quercus. Ambas localidades compartieron 4 especies no presentes en los sitios laguna Bosque Azul y laguna Montebello: Boletellus jalapensis, Lactarius costaricensis, Lactarius indigo y Strobilomyces confusus. Las 2 especies de Lactarius fueron exclusivas para los sitios laguna Encantada y laguna Ensueño.
Laguna Bosque Azul y laguna Montebello compartieron 7 especies no presentes en las lagunas Encantada y Ensueño: Craterellus fallax, Hydnum umbilicatum, Inocybe dulcamara, Russula cf. amoenolens, Russula vesicatoria, Scleroderma cepa y Suillus hirtellus. De éstas, Russula vesicatoria y Scleroderma cepa fueron exclusivas para estas 2 localidades. Antelá comparte 21 taxones con las 4 localidades antes mencionadas, es decir, el 77.8% de su total y los sitios grutas de San Rafael y laguna Escondida compartieron 7 especies: Craterellus ignicolor, Craterellus lutescens, Cantharellus minor, Craterellus fallax, Turbinellus floccosus, Helvella macropus y Lactarius chrysorrheus.
Cinco Lagos y laguna San Rafael (no incluidas en el análisis previo) fueron las 2 localidades con mayor diferencia de las anteriores localidades muestreadas. Ambas tuvieron el menor número de ejemplares recolectados, 5 y 4 respectivamente, y compartieron únicamente a Coltricia cinnamomea.
Discusión
Debido a la naturaleza biológica de los hongos, un problema importante ha sido el reconocimiento de los individuos para estimar su abundancia y otros parámetros ecológicos. Se han realizado esfuerzos para proponer estrategias de análisis y muestreo en casos particulares de hongos, incluyendo los que forman ectomicorrizas (Rossman et al., 1998; Mueller et al., 2004). En México, la mayoría de los estudios ecológicos de hongos se han enfocado en conocer aspectos como la abundancia, biomasa, disponibilidad, diversidad, frecuencia, entre otros; sin embargo, en todos ellos la metodología empelada para estimar la abundancia de los individuos ha sido diferente y de igual forma la mayoría solo se enfocan en especies comestibles, útiles o de importancia biocultural (Garibay- Orijel et al., 2009; Montoya et al., 2014; Ruan-Soto, 2014) y por lo tanto, se presenta un sesgo si se pretende evaluar los hongos que forman ectomicorrizas.
En un estudio reciente llevado a cabo en diferentes localidades tanto de los Altos de Chiapas, como de las Tierras Bajas, Ruan-Soto (2014) observó que las especies con valores más altos de abundancia pertenecen a los géneros Clavulina, Ramaria y Turbinellus, los 2 últimos también están presentes en el PNLM; sin embargo, en este caso, no fueron de los más abundantes, incluso para Ramaria se obtuvieron valores muy bajos. En ambos estudios se registran valores altos de abundancia para especies de los géneros Cantharellus, Craterellus, Laccaria y Suillus, lo que puede resultar importante ya que incluyen especies con importancia alimentaria, aunque desaprovechadas en el PNLM. Con respecto a la riqueza obtenida, el autor antes mencionado, reporta 38 especies de las que 18 taxones son ectomicorrizógenos y en el presente trabajo se incluyen 93 especies todas ellas formado asociaciones mutualistas. En el inventario del PNLM, no se registraron Amanita vaginata ni Lyophyllum decastes. Se recolectaron taxones de los géneros Clavariadelphus y Clavulina, pero no se incluyeron en el listado debido que en la Tabla 1 únicamente se citan materiales con determinaciones a nivel de especie.
En bosques de Pinus-Quercus de Ixtlán de Juárez, Oaxaca, Garibay-Orijel et al. (2009) obtuvieron una riqueza de 81 especies de hongos silvestres comestibles, de los cuales 61 especies son ectomicorrizógenas. Laccaria y Cantharellus están entre las especies con valores de abundancia altos. Se cita al género Laccaria como uno de los 5 más abundantes en los bosques templados de México en los que se han realizado estudios ecológicos, lo que ocurre también en el PNLM, así como en los Altos de Chiapas. Del mismo modo, Montoya et al. (2014), en un bosque de coníferas en el Parque Nacional La Malinche, registraron una riqueza de 61 especies de hongos comestibles silvestres, de los que 44 son ectomicorrizógenas. Las especies más abundantes fueron Laccaria trichodermophora, Hebeloma aff. mesophaeum, Helvella lacunosa, Suillus pseudobrevipes y Helvella crispa. Por lo tanto, es probable que las especies del género Laccaria en México sean de las más abundantes en los bosques, incluyendo los bosques mesófilos, pero se requieren más estudios para confirmar estas observaciones. Lo mismo se observó para el caso de los taxones pertenecientes al género Cantharellus en Ixtlán de Juárez, pero al tratarse de zonas con diferente tipo de vegetación y clima que en el PNLM, las especies que se presentan en ambos lugares son diferentes. Al comparar los valores de riqueza, se observa que el bosque mesófilo de montaña del PNLM presenta un mayor número de especies en comparación con los bosques de Ixtlán de Juárez y con los bosques de coníferas en el Parque Nacional la Malinche; sin embargo, son necesarios más estudios para conocer mejor la diversidad de estos y otros tipos de vegetación, para poder determinar con mayor precisión los patrones de diversidad de los ecosistemas en donde los hongos ectomicorrizógenos desempeñan un papel fundamental. También es necesario incrementar los estudios enfocados en el conocimiento de la identidad taxonómica de las especies de hongos de México ya que esto permitirá una mejor comparación entre los diferentes estudios.
Sólo 6 especies de los hongos ectomicorrizógenos recolectados son consumidas y conocidas regionalmente: Turbinellus floccosus (bocina o trompeta), Hydnum albidum (lengua de toro), Hydnum umbilicatum (lengua de toro), Lactarius indigo (kanchayita azul), Lactarius miniatosporus (kanchayita roja) y Ramaria cystidiophora var. citronella (barbas de chivo). Los nombres tradicionales asignados a las especies están en su mayoría en español y pocos en tojolabal, como en el caso de kanchayita (k’an, amarillo, chay, pescado) y hacen referencia a la apariencia de los hongos, ya sea a la forma y/o al color. Estas especies son preparadas de diferentes formas para su consumo: asados a las brasas, en caldo, mole, quesadillas, recado (el hongo se pone a hervir en suficiente agua, se adiciona saborizante, especias y epazote; con la finalidad de que adquiera una consistencia espesa y se le agrega masa de maíz) o fritos con huevo. Turbinellus floccosus es un hongo que debe limpiarse raspando las escamas y la superficie del estípite; se pone a hervir y se deshecha el agua, después se fríe o se prepara de alguna de las formas antes indicadas. La recomendación con este hongo es comer poca cantidad, ya que el exceso conduce a tener trastornos estomacales. Las 2 especies de Hydnum son asados en las brasas y con sal, R. cystidiophora var. citronella es preparada en recado (con verduras) y Lactarius spp., se preparan con cualquiera de las recetas generales, o en tamales.
Para estas 6 especies, las personas señalaron como lugar de crecimiento la tierra, es decir, que crecen sobre el suelo, a diferencia de otros hongos no ectomicorrizógenos, que pueden desarrollarse sobre palos (troncos), hojarasca o estiércol, varios de ellos son conocidos y utilizados como alimento por los habitantes de las diferentes comunidades del municipio La Trinitaria. La información sobre la época de crecimiento de los hongos es conocida de manera precisa y las personas señalaron que hongos como Lactarius miniatosporus se puede encontrar de abril a septiembre, Turbinellus floccosus de mayo a octubre, las 2 especies de Hydnum de junio a septiembre, Lactarius indigo de julio a octubre y Ramaria cystidiophora var. citronella durante todo el año. Varios de estos hongos son importantes debido a que se utilizan para preparar guisos que son ofrecidos en quesadillas a los turistas que acuden al parque durante la temporada de lluvias; por lo tanto representan un aporte al ingreso económico de las familias que participan en las actividades de turismo. Hay especies de hongos recolectados alrededor de las lagunas que están reportados en la literatura como comestibles, pero no son utilizados como tal por las personas entrevistadas, por ejemplo: Craterellus lutescens, Craterellus ignicolor, Craterellus fallax, Helvella crispa, Hygrophorus russula, Laccaria amethystina, Laccaria laccata, Laccaria trichodermophora, Lactifluus corrugis, Lactifluus volemus, Russula modesta, Russula delica var. centroamericana, Russula cf. cyanoxantha y Strobilomyces confusus. Es muy probable que el incrementar el número de personas entrevistadas y las localidades visitadas, podría generar registros de especies comestibles más completos, lo que se sugiere en estudios posteriores. La información obtenida confirma que Cantharellus lateritius (chikintaj) es uno de los 2 hongos más importantes desde el punto de vista cultural, entonces se sugiere como una especie que debe incluirse en la lista de especies prioritarias para su conservación en el parque, ya que además es un hongo ectomicorrizógeno, lo que junto con otros hongos de la lista obtenida, constituyen ejemplos de especies con importancia biocultural de los cuales debe priorizarse su conservación.
Concluyendo, los bosques del Parque Nacional Lagunas de Montebello, Chiapas, presentan una gran diversidad de hongos ectomicorrizógenos. Las especies más representativas con base en los valores de abundancia y distribución son: Laccaria amethystina, Craterellus lutescens, Suillus decipiens, Craterellus ignicolor, Cantharellus minor y Coltricia cinnamomea con 7.41%, 5.78%, 5.78%, 5.19%, 3.70% y 3.70%, respectivamente.
Los análisis de completitud realizados mediante el estimador de la cobertura de la muestra indican que las comunidades de hongos potencialmente ectomicorrizógenos están, en general, bien representadas en el muestreo, pero hacer recolecciones adicionales permitirían incrementar el número de especies de aquéllos taxones muy poco frecuentes. En la mayoría de las localidades se requiere de un mayor esfuerzo de recolección para completar las listas locales de hongos ectomicorrizógenos de las comunidades estudiadas en el PNLM, Chiapas. Haber estandarizado la cobertura de la muestra fue una buena estrategia de análisis porqué evitó el sesgo dado por las diferencias en la intensidad de muestreo para cada laguna y es, por lo tanto, una buena herramienta para comparar datos de riqueza en sitios con diferente esfuerzo de muestreo. También los resultados obtenidos, muestran que es una herramienta útil para priorizar los sitios de muestreo en futuros inventarios de la misma zona y en otras zonas en las que se hagan estudios de hongos y que presenten estas características. En este caso particular, se sugiere en el futuro enfatizar la recolección de hongos en las zonas aledañas a las lagunas Pomarosa, Antelá, Escondida y Pojoj, hasta lograr obtener un porcentaje del 80% al menos, lo que permitirá tener mejor representada la diversidad de hongos epigeos ectomicorrizógenos en el PNLM. El resultado del análisis realizado con incidencias corroboran los valores obtenidos con las abundancias, ya que ambos mostraron la misma tendencia.
Se considera de gran importancia la realización de inventarios más exhaustivos en el parque con un mayor esfuerzo de muestreo, ya que es muy probable que se amplíe el número de especies de hongos de manera considerable en los sitios que tuvieron porcentajes menores al 80%. Se recomienda hacer recolecciones un periodo más largo (más años) y semanales durante la temporada de fructificación de los hongos, evaluando las distintas condiciones climáticas de cada época y de cada año de muestreo, y extender los muestreos a otros grupos tróficos de hongos. Los hongos registrados en este trabajo tienen el potencial de ser utilizados para la reforestación, ya que se asocian con plantas existentes en los sitios, pero su aplicación práctica debe de ser antecedida por otros estudios relacionados con el aislamiento, caracterización fisiológica y elaboración de inoculantes para realizar ensayos de micorrización controlada en laboratorio, vivero y campo, entre otros, con el objeto de seleccionar aquellas especies que tengan potencial para promover la producción exitosa de plantas con fines de reforestación.
En general, los resultados que se obtuvieron en este trabajo muestran la importancia de llevar a cabo investigaciones para conocer la diversidad alfa y beta que no solo incluyan la riqueza absoluta, sino también el análisis de la diversidad taxonómica de hongos, porque así se entenderán más acertadamente aquellos procesos ecológicos y evolutivos que afectan los patrones de diversidad de las especies a diferentes escalas espacio-temporales.

