











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En el estado de Sonora (NO de México) se presenta una heterogeneidad litológica que comprende edades desde el Proterozoico al Reciente. Particularmente, sobre el Paleozoico Superior de la sierra Agua Verde, Ochoa y Sosa (1993) realizaron estudios geológicos y definieron la formación La Joya. Stewart et al. (1999) contribuyeron con la investigación geológica de la región. Posteriormente, Buitrón-Sánchez et al. (2005, 2007a, b, 2012, 2015), Gómez-Espinosa et al. (2008) y Mendoza-Madera et al. (2004) publicaron trabajos sobre diversos aspectos, entre ellos, el contenido biótico, la tafonomía, la paleoecología y la paleogeografía de la región.
Para esta localidad, Ochoa y Sosa (1993) han propuesto una estratigrafía conformada informalmente por el grupo San José, que incluye a las formaciones Puerto Blanco y Cuarcita Proveedora, Buelna y Arrojos del Cámbrico, además del grupo Agua Verde, en el que se encuentran las formaciones El Boquinete (Ordovícico), El Pollo (Devónico), Santiago (Misisípico), La Joya (Pensilvánico) y Tuntunudé (Pérmico).
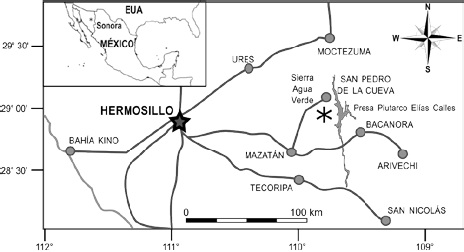
La sierra Agua Verde se lcaliza aproximadamente a 110 km al este de Hermosillo, Sonora, en el municipio de Villas Pesqueira, con coordenadas 29°12’10.50” N, 109°54’11.65” O (Fig. 1).
La formación La Joya está constituida por 300 m de caliza intercalada con lodolita y lentes arenosos. Los estratos de caliza contienen una biota abundante y diversa que consiste en algas, foraminíferos, corales, briozoos, braquiópodos y crinoides (Fig. 2). La cima de la formación La Joya presenta contacto por falla normal con la formación Tuntunudé y es correlacionable con la formación Picacho Colorado (Noll, 1981) y la caliza Aparejo (Vega y Araux, 1987) de ambientes marinos de plataforma con caliza y caliza ligeramente recristalizada, intercaladas con dolomía y caliza bioclástica que se presentan en el arroyo Agua Caliente.

Figura 2 Columna estratigráfica de la formación La Joya en el arroyo La Joya, sierra Agua Verde (modificada de Buitrón et al., 2015).
El objetivo principal de este trabajo es dar a conocer los braquiópodos del Paleozoico tardío (Pensilvánico temprano) en la región de sierra Agua Verde y contribuir al conocimiento de la estratigrafía y la paleogeografía de la región.
En este caso en particular, el material procede de la formación La Joya (unidad informal) en el arroyo Agua Caliente, municipio de Villas Pesqueira, Sonora. El material se recolectó en afloramientos de la formación y se describieron las características litológicas en campo. El material fue preparado con percutor neumático para eliminar el exceso de sedimento con el fin de permitir su identificación a especie. Las dimensiones de los ejemplares fueron tomadas con un calibrador vernier digital.
Materiales y métodos
La identificación de los ejemplares se hizo con la clasificación propuesta en el Treatise on Invertebrate Paleontology (Williams et al., 2000, 2002, 2006, 2007).
El material se encuentra depositado en la Colección de Paleontología “Dr. Emilio Almazán Vázquez” del Departamento de Geología de la Universidad de Sonora, con números de catálogo USON-DG-29 al 31, 33, 35, 37, 39, 42, 52 al 55, 57 al 60, 63, 65, 66, 68 al 78, 82, 83 y 147.
Paleontología sistemática
Phylum Brachiopoda Duméril, 1806
Clase Strophomenata Williams, Carlson, Brunton, Holmer y Popov, 1996
Orden Productida Sarycheva y Sokolskaya, 1959
Familia Productellidae Schuchert y Le Vene, 1929
Género Plicatifera Chao, 1927
Plicatifera sp.
(Fig. 3a)

Figura 3 a) Plicatifera sp.: vista externa de la valva ventral, ejemplar USON-DG-147. b) Kozlowskia capaci: vista externa de la valva ventral, ejemplar USON-DG-52; c) y f) Composita sp. cf. C. subtilita. c) vista anterior del ejemplar USON-DG-57; f) vista dorsal del ejemplar USON-DG-55 (molde interno); d) y e) Linoproductus sp. cf. L. eastoni, vistas externa e interna de la valva ventral del ejemplar USON-DG-50; g) y h) Anthracospirifer occiduus (Sadlick, 1960): g) vista externa de la valva ventral del ejemplar USON-DG-82, h) vista externa de la valva ventral del ejemplar USON-DG-83. Escala = 10 mm.
Descripción: la concha es subcuadrangular, convexa de umbo agudo; aurículas antecedidas por una deflexión ligera; arrugas irregulares más sinuosas en la región anterior, costelación recta; espinas huecas y dispersas, ligeramente inclinadas hacia la parte anterior. El ejemplar presenta 12.8 mm de longitud, 16.4 mm de anchura y 10 mm de altura.
Resumen taxonómico
Material: el ejemplar procede de un nivel de caliza de la parte media de la formación La Joya (unidad informal) en la localidad del arroyo Agua Caliente, en la sierra Agua Verde, municipio de Villas Pesqueira, Sonora. En este nivel se observa empaquetamiento variable y aloquímicos sin orientación preferencial dentro del sedimento.
Distribución: Muir-Wood y Cooper (1960, p. 203) mencionan a Plicatifera para el Carbonífero inferior (Viseano) de Irlanda, Bélgica y Rusia (Brunton, 1966; Brunton et al., 1993; Mundy, 1994; Ludford, 1951). Li et al. (1986) y Chen et al. (2003) la citan para el Pensilvánico de China. Otras menciones del género son para el Triásico de Japón (Nakazawa, 1971).
Número de catálogo: USON-DG-147.
Comentarios taxonómicos
Plicatifera sp., tiene semejanza con Plicatifera plicatilis (Sowerby, 1824), del Misisípico (Viseano) de Irlanda (Brunton, 1966: p. 204, lám. 6, fig. 6) y con Plicatifera lipoensis Grabau del Pérmico temprano de China (Grabau, 1934: p. 47, lám. 2, fig. 22), pero difieren en el patrón de ornamentación que es menos sinuoso. Plicatifera chaoi del Pérmico de China es también comparable por el número de arrugas, pero la valva dorsal es más cuadrangular (Grabau, 1936: p. 171, lám. 6, fig. 9).
Género Kozlowskia Fredericks, 1933
Kozlowskia capaci d’Orbigny 1842
(Fig. 3b)
Descripción: concha estrófica, cóncavo-convexa, con el umbo curvo; aurículas de tamaño variable con reticulación inconspícua; rugosidades variables en tamaño; valva ventral convexa, con costillas que se desarrollan desde el umbo y se ensanchan ligeramente hacia la comisura; espinas pequeñas en la parte anterior de la concha; canal medio en la valva ventral; proceso cardinal sésil, con crestas triangulares; valva dorsal cóncava con ornamentación similar a la de la valva ventral. Los ejemplares tienen 12.0 a 14.3 mm de longitud, 11.9 a 17.2 mm de anchura y 6.6 a 8.88 mm de altura.
Resumen taxonómico
Material: los ejemplares proceden de estratos de caliza y limolita bioclástica de la parte media y alta de la formación La Joya en la localidad Arroyo Agua Caliente, municipio de Villas Pesqueira, Sonora. En estos estratos se presentó de manera frecuente recristalización y/o reemplazamiento.
Distribución: la especie ha sido descrita del Pensilvánico medio (Desmoinesiano) y Carbonífero superior-Pérmico (Gzheliano-Kunguriano) de Bolivia, Perú y EUA (King, 1931; Newell et al., 1953; Pérez-Huerta y Sheldon, 2006).
Número de catálogo: USON-DG-39, 42, 52, 53, 60, 69, 70, 71.
Comentarios taxonómicos
Se determinó el material de Sonora como Kozlowskia capaci d’Orbigny, por la concha pequeña con umbo poco sobresaliente, moderadamente proyectado por detrás de la línea de la charnela y el tipo de costelación, características de la especie mencionada.
Familia Linoproductidae Stehli, 1954
Género Linoproductus Chao, 1927
Linoproductus sp. cf. L. eastoni Gordon 1975
Descripción: concha cóncavo-convexa, orbicular, con arrugas y líneas de crecimiento conspícuas; costillas de menor anchura que los espacios intercostales, algunas bifurcadas desde el primer tercio anterior; valva ventral con arrugas conspícuas y espinas dispersas, orientadas hacia la parte anterior; umbo grande y curvo; proceso cardinal trilobulado con lóbulos subcuadrados y divergentes; crestas cardinales inclinadas hacia la parte superior. Los ejemplares tienen 22.2 a 33.1 mm de longitud, 21.8 a 43.3 mm de anchura y 12.8 a 16 mm de altura.
Resumen taxonómico
Material: los ejemplares proceden de niveles de caliza arenosa con coloraciones gris claro, marrón-rojizo o crema de la parte media de la formación La Joya en la localidad Arroyo Agua Caliente, municipio de Villas Pesqueira, Sonora. En estos niveles se exhiben diferentes grados de empaquetamiento con bio- y litoclastos mal clasificados en los que se aprecia también diferente grado de redondeamiento, así como presencia de material fino limolítico con oxidación.
Distribución: Linoproductus eastoni se ha registrado en EUA para el Carbonífero de Nevada (Langenheim et al., 1985; Pérez-Huerta y Sheldon, 2006) y Pérmico de Wyoming (Gordon, 1975).
Número de catálogo: USON-DG-35, 54, 65.
Comentarios taxonómicos
Los ejemplares de Sonora se determinaron como Linoproductus sp. cf. L. eastoni por la forma subcuadrangular del margen y convexidad semejante de la concha, sin embargo los ejemplares presentan diferente altura de la valva dorsal y el umbo es menos prominente y recurvado. L. prattenianus es similar pero se distingue en la costelación y en la presencia de espinas en hileras cerca de la línea de charnela.
Clase Rhynchonellata Williams, Carlson, Brunton, Holmer y Popov, 1996
Orden Athyridida Boucot, Johnson y Staton, 1964
Familia Athyrididae Davidson, 1881
Género Composita Brown, 1845
Composita sp. cf. C. subtilita Hall, 1852
Descripción: la concha es lisa, ventribiconvexa y subtriangular; región umbonal alargada y angosta con el umbo redondeado y proyectado más allá de la línea de la charnela; foramen mesotírido, pequeño, ovalado y con un ligero reborde; valva dorsal con surco medio bien definido, desarrollado posteriormente y con el braquidio de tipo espiralio. La concha tiene líneas de crecimiento regularmente espaciadas. La longitud del ejemplar es de 27.4 mm, con una altura de 13.2 mm y una anchura de 24.4 mm.
Resumen taxonómico
Material: los ejemplares proceden de niveles de caliza mal clasificada con evidencia de oxidación y recristalización, recolectado en estratos de caliza de la parte media de la formación La Joya en la localidad Arroyo Agua Caliente, municipio de Villas Pesqueira, Sonora.
Distribución: Composita subtilita Hall, 1982 se citó para el Misisípico de Canadá (Dawson, 1873), Carbonífero de EUA (Coogan, 1960; Hoare, 1961) y Pérmico de Bolivia (Badyrka et al., 2013), Perú (Newell et al., 1953) e Indonesia (Roemer, 1880).
Número de catálogo: USON-DG-55, 57.
Comentarios taxonómicos
El material de Sonora presenta afinidad con las especies de Composita poposiensis (Gordon, 1975: p. 61, lám. 8, figs. 6-20; lám. P, figs. 1-15, 31-36) y C. ozarkana (Easton, 1962: p. 77, lám. 11, figs. 5-7), del Misisípico (Chesteriano) de Wyoming y Utah. C. poposiensis se diferencia por una mayor redondez y elevación del pliegue, mientras que C. ozarkana posee un pliegue dorsal más redondeado y una relación longitud-anchura menor. Por su parte, las especies C. affinis Girty (Cooper y Grant, 1976) y la variedad lisa de C. stalagmiumCooper y Grant (1976) registradas para el Pérmico del oeste de Texas, EUA, son semejantes al ejemplar de Sonora, pero tienen mayor altura y convexidad.
Orden Spiriferida Waagen, 1883
Familia Spiriferidae King, 1846
Género Anthracospirifer Lane, 1963
Anthracospirifer occiduus (Sadlick, 1960)
Anthracospirifer occiduus (Sadlick, 1960); sinonimia completa en Butts (2007, p. 58, figs. 5.34-5.36).
Descripción: la concha es ancha, deprimida en la parte anterior de la valva dorsal y con mayor profundidad del surco hacia la comisura; interárea ventral amplia y ligeramente cóncava, con estrías a partir de la región cardinal; palíntropo aserrado; surco de la valva ventral con 3 a 5 costillas subparalelas, más anchas que los espacios intercostales; costillas marginales y aurículares divididas hacia la tercera parte anterior; aurículas alargadas y precedidas por deflexiones poco marcadas; umbo sobresaliente de la línea de la charnela formando una prolongación subtriangular y subredondeada; lamelaciones concéntricas, subparalelas, más desarrolladas cerca de la mitad anterior; deltirio triangular, abierto, más largo que ancho, con placas deltidiales incipientes; interior de la valva dorsal con extremos cardinales ligeramente mucronados y redondeados.
Resumen taxonómico
Material: los ejemplares proceden de estratos de caliza gris claro mal clasificada con contenido variable de intraclastos y fragmentos dispersos de conchas de otros braquiópodos como estrofoménidos que presentan conservación moderada a buena y recristalización parcial. Estos ejemeplares se recolectaron en la parte media de la formación La Joya en la localidad Arroyo Agua Caliente, municipio de Villas Pesqueira, Sonora.
Distribución: Anthracospirifer occiduus (Sadlick, 1960) se ha descrito en México para el Carbonífero (Chesteriano-Desmoinesiano) de Nochixtlán, Oaxaca (Torres-Martínez y Sour-Tovar, 2012). En EUA, esta especie tiene una amplia distribución en Arizona (Brew y Beus, 1976; Gordon, 1982), Idaho (Butts, 2007), Kansas (Williams et al., 1937, 1965), Missouri (Hoare, 1961), Nevada (Lane, 1964), Nuevo México (Gehrig, 1958), Ohio (Sturgeon y Hoare, 1968), Utah (Hoare, 1961) y Wyoming (Sando et al., 1975).
Número de catálogo: USON-DG-82, 83.
Comentarios taxonómicos
Anthracospirifer birdspringensis (Lane, 1963: p. 388, lám. 44, figs. 17 y 18; lám. 45, fig. 3) se asemeja a la especie de Sonora en el número de costillas situadas en el surco ventral y en el número de costillas laterales, aunque se diferencia porque A. birdspringensis es de menor tamaño, y de mayor longitud en la línea de la charnela, con el margen anterior más redondeado.
Los ejemplares de Spirifer opimus Hall var. occidentalis estudiados por Girty (1927, p. 433, lám. 27, figs. 28-31), cuentan con igual número de costillas y varían solamente en la forma del umbo, siendo ligeramente más triangular con respecto a la especie A. occiduus. También se considera que Anthracospirifer occiduus es diferente de A. birdspringensis porque las costillas laterales y del surco son menos numerosas. Además, Lane (1964, p. 783) diferencia a A. occiduus y A. opimus por el número de bifurcaciones en los flancos, siendo menores en A. opimus.
Familia Retziidae Waagen, 1883
Género Hustedia Hall y Clarke, 1893
Hustedia mormoni (Marcou, 1858)

Figura 4 Hustedia mormoni (Marcou, 1858) y Reticulariina sp. a) y b) Hustedia mormoni (Marcou, 1867), vistas externas de las valvas dorsal y ventral del ejemplar USON-DG-67, c)-f) Reticulariina sp., c) y f) vistas externas de las valvas ventral y dorsal del ejemplar USON-DG-73; d) y e) vistas externas de las valvas ventral y dorsal del ejemplar USON-DG-78. Escala = 5 mm.
Hustedia mormoni (Marcou, 1858) Mudge et al., 1962, p. 85, lám. 15, figs. 6-8 (sinonimia completa).
Descripción: concha biconvexa con contorno subpentagonal; pliegues radiales simples que se desarrollan a partir del umbo en número de 10 a 13 con surcos intercostales profundos y de flancos redondeados; línea de la charnela curva, con menor longitud que en la valva dorsal; ápice umbonal curvo y elevado. Los ejemplares tienen 5.2 a 8.5 mm de longitud, 4.0 a 6.9 mm de anchura y 3.1 a 5.8 mm de altura.
Resumen taxonómico
Material: los ejemplares proceden de niveles limolíticos y de caliza con coloración gris claro de la parte media de la formación La Joya en la localidad Arroyo Agua Caliente, municipio de Villas Pesqueira, Sonora. En algunos estratos de caliza se exhiben diferentes grados de empaquetamiento, oxidación y orientación de los aloquímicos incluidos en la matriz.
Distribución: para México, la especie ha sido citada del Pérmico de Hidalgo (Carrillo-Bravo, 1965). La distribución de Hustedia mormoni para EUA, incluye el Pensilvánico de Missouri, Nebraska, Ohio (Dunbar y Condra, 1932; Hoare, 1961) y el Pérmico de Iowa, Kansas, Nebraska, Oklahoma y Texas; Pensilvánico (Missouriano-Virgiliano) de Iowa, Kansas, Nuevo México y Pensilvania (Harper, 1989; Hoover, 1981; Stehli y Grant, 1971).
Número de catálogo: USON-DG-58, 59, 63, 66, 67, 68, 72.
Comentarios taxonómicos
Hustedia mormoni de Sonora concuerda en la descripción y en las dimensiones con los ejemplares estudiados por Dunbar y Condra (1932) y Mudge et al. (1962), presentando ligeras variaciones en la costelación. Entre las especies de Hustedia con las que se ha comparado el material de Sonora se encuentran, H. meekana y H. meekana var. trigonalis las cuales se diferencian en la terminación más amplia de las costillas en la parte anterior (Girty, 1908: lám. 14, fis. 22-26; lám. 21, fig. 5-8, lám. 24, fig. 14; lám. 21, fig. 9). H. bipartita (Girty, 1908: lám. 30, figs. 19, 20) se diferencia por presentar el margen anterior más redondeado y ondulado. Con relación a H. papilataShumard (1858) del Pérmico medio de Delaware, EUA es similar en forma, tamaño, número y grosor de costillas, pero la valva braquial es de menor convexidad y presenta costillas más t enues. H. grandicosta del Pérmico de China (Davidson, 1862) tiene costillas más gruesas y espaciadas. H. mormoni es particularmente semejante a H. rotunda (Lane, 1962: p. 905, p. 127, figs. 38-41) y H. miseri subsp. gibbosa (Lane ,1962: p. 906, lám. 127, figs. 42-45) del Pensilvánico de Nevada, EUA, pero se diferencian por un tamaño mayor, menor número de costillas, lamelación concéntrica más pronunciada hacia la primera mitad anterior de la valva dorsal, y el umbo con mayor grado de curvatura. Por su parte, H. rotunda es similar en la altura de la concha y en la relación longitud-anchura y longitud-altura, pero se diferencia por su mayor espaciamiento intercostal, mayor convexidad posterior en la valva dorsal, y por el ápice umbonal con ángulo semirecto.
Superfamilia Pennospiriferinoidea Dagys, 1972
Familia Reticulariinidae Waterhouse, 1978
Género Reticulariina Fredericks, 1916
Reticulariina sp.
Descripción: concha densamente punctuada, con la línea de la charnela ligeramente más pequeña que la anchura; márgenes laterales curvos con una concavidad ligera; comisura anterior uniplegada; valva dorsal delgada pero gruesa hacia la región apical; ápice del umbo curvo y cubre parcialmente al deltirio; área cardinal triangular y aplanada con estrías irregulares; costillas redondeadas, originadas a partir del umbo, con espacios intermedios de una menor longitud formando surcos con la parte baja suavizada. El interior de la valva dorsal presenta placas cardinales poco sobresalientes y divergentes con engrosamientos anteriores y terminaciones laterales redondeadas.
La valva ventral es mucho más alta que la dorsal, con el área cardinal triangular y aplanada y una ligera curvatura longitudinal que presenta un pliegue que se desarrolla desde el umbo y que se profundiza hacia la parte anterior. En el surco ventral se aprecian regularmente de 2 a 4 pliegues internos que inician en la parte media y se hacen más gruesos hacia el margen anterior. Ambas valvas presentan lamelación concéntrica muy marcada, espaciada en la parte anterior de la concha y con mayor grosor anterior. Los ejemplares tienen de 5.5 a 9.8 mm de longitud, de 8.0 a 13.9 mm de anchura y de 5.3 a 9.0 mm de altura en, con 11 a 16 costillas en la valva dorsal.
Resumen taxonómico
Material: los ejemplares se recolectaron en estratos de caliza gris claro-marrón rojiza y de limolita con diferentes grados de oxidación que se encuentran en la parte media de la formación La Joya en la localidad Arroyo Agua Caliente, municipio de Villas Pesqueira, Sonora. En esta parte de la sección se presentan estratos con matriz bio-lito-clástica, con elementos que exhiben orientación diferencial y variaciones en el grado de empaquetamiento.
Distribución: son escasos los registros de Reticulariina en México. Cooper et al. (1953) describieron a Spiriferellina sonorensis. Posteriormente, Cooper y Grant (1976) determinaron que esta especie es sinónima de Reticulariina sonorensis del Pérmico medio (Wordiano) de Cerros Los Monos y El Antimonio, Sonora. Para Arizona, EUA, la especie se ha mencionado de la región de Little Dragoon y de las Montañas Mule (Cooper, 1953; Cooper et al., 1965).
Número de catálogo: USON-DG-29, 30, 31, 33, 37, 73-78.
Comentarios taxonómicos
Las especies del género Reticulariina presentan similitudes con especies de Punctospirifer (Gehrig, 1958: p. 12) cuyas diferencias radican en 4 características importantes: 1) presencia de espinas a lo largo de la parte anterior del margen, 2) área cardinal de la valva ventral regularmente curvada, 3) presencia de lamelas menos espaciadas y 4) menor cantidad de pliegues en tamaños comparables. Reticulariina se diferencia además de Punctospirifer por el desarrollo de la inter-área dorsal con mayor elevación y por presentar un margen de variabilidad muy amplio en sus extremos cardinales que pueden ser desde subangulares a redondeados. Sour-Tovar et al. (2005) consideran que algunas especies de Reticulariina Fredericks, 1916, pueden ser confundidas con especies del género SpiriferellinaCooper y Grant, 1976, sin embargo, se diferencian por el tipo de espinas, por la mayor anchura de la concha y las espinas más cortas y gruesas. Además, Spiriferellina posee generalmente lamelas más espaciadas hacia la región anterior y las valvas son comúnmente más lisas, presentando pliegues de mayor profundidad y redondez.
De las especies de Reticulariina estudiadas por Cooper y Grant (1976) solamente están relacionadas Reticulariina pusilla y Reticulariina sp. 1 (Cooper y Grant, 1976: lám. 702, figs. 7 y 9, lám. 705, figs. 45-49). Sin embargo, se observa que poseen costillas más gruesas, redondeadas y en menor número, con surcos más amplios y crenulación más marcada para el caso de R. pusilla, siendo también más alta la inter-área dorsal y con lamelación imbricada muy marcada en Reticulariina sp. 1. De otras especies estudiadas por Cooper y Grant (1976) se diferencia por la lamelación más marcada, la altura de la inter-área dorsal, el grosor de la costelación, la profundidad de los interespacios, la inclinación de los bordes laterales posteriores y la forma general y longitud del margen, más amplio y de forma subtriangular. Sin embargo, Reticulariina pusila tiene una relación longitud-altura más pronunciada, con los tamaños comparables por debajo de los presentados en los ejemplares de sierra Agua Verde. Respecto de Reticulariina sp. 1, Cooper y Grant (1976) no incluyen una caracterización de la misma, pero mencionan que pudieran tratarse de ejemplares juveniles de R. cerina. Los ejemplares de Sonora tienen longitudes medias parecidas a las presentadas por las especies con que se compararon. De tratarse de alguna de estas especies es factible que sea un estadio juvenil.
Consideraciones paleoecológicas
La asociación biótica de la sierra Agua Verde está constituida por esponjas (Chaetetes), corales (Syringopora), briozoos (Prismopora), vermes, braquiópodos (Plicatifera sp., Kozlowskia capaci, Linoproductus sp. cf. L. eastoni, Composita sp cf. C. sp. cf. subtilita, Anthracospirifer occiduus (Sadlick, 1960), Hustedia mormoni (Marcou, 1858) y Reticulariina sp.), gasterópodos (Donaldina) y crinoides (Cyclocaudex, Cyclocrista, Heterostelechus, Lamprosterigma, Preptopremnum, Pentaridica), que permiten caracterizar condiciones de depósito en una plataforma somera con aporte de terrígenos de salinidad normal en condiciones tropicales y de profundidad somera a moderada (Cocks, 1967; Fürsich y Hurst, 1974; Stevens, 1966).
La distribución de los braquiópodos en el pasado geológico está determinada por una serie de factores ambientales tales como la profundidad, el tipo de sustrato y la disponibilidad de alimento (Ager, 1967; Rudwick, 1962). Fürsich y Hurst (1974, p. 889) consideran que en algunos braquiópodos de aguas profundas (órthidos, estrofoménidos, rinconélidos) se observa una tendencia a un menor tamaño y que esta tendencia es inversa con respecto a lo que ocurre con espiriféridos y pentaméridos; otras adaptaciones relacionadas con la profundidad del agua son el desarrollo de aletillas y surcos para la separación de corrientes en planos horizontales y verticales, y la formación de fuertes deflexiones medias para facilitar la separación de las corrientes inhalantes y exhalantes. Ager (1967), Rudwick, (1962), Fürsich y Hurst (1974) proponen también una serie de adaptaciones morfológicas para los braquiópodos que habitan zonas con turbulencia como son el engrosamiento de la concha, el desarrollo de costillas gruesas, la formación de aberturas pedunculares fuertes y la generación de aletillas y surcos más marcados; para el caso de conchas resupinadas de braquiópodos de aguas de fondos suaves mencionan la generación de amplias superficies de contacto con el sustrato.
Por otra parte, es común encontrar incrustaciones, que corresponden a crecimientos de organismos epibiontes o depósitos de minerales, y que comúnmente se encuentran sobre las partes duras de los organismos hospederos, como puede ser en las conchas de braquiópodos que habitan principalmente aguas someras. Estas incrustaciones pueden formarse mediante el crecimiento de otros braquiópodos o por el desarrollo de algas, foraminíferos, serpúlidos y briozoos, los cuales pudieron haberse establecido durante el tiempo de vida del organismo hospedero o una vez que están enterrados parcialmente o que sus restos “post-mortem” son depositados en el sustrato.
La asociación simbiótica con otros organismos puede proporcionar información. Si se observa que los organismos incrustantes se desarrollan hacia la región anterior de la línea de charnela proporciona información sobre la posición y orientación que debieron presentar los braquiópodos durante su desarrollo.
Además, en los braquiópodos y otros invertebrados se pueden evidenciar condiciones de depredación a partir de la reparación de las partes duras. En el caso de los braquiópodos, la reparación de las valvas puede notarse con un cambio en la ornamentación ya sea porque ésta se encuentra interrumpida o disminuida, o porque se pierde una parte importante de la concha debido a la actividad de organismos durófagos, tales como peces y artrópodos. Así, una concha que es reparada se ve también con cambios en el arreglo mineralógico de alguna de las capas que la conforman. Por su parte, las perforaciones son debidas a organismos que producen sustancias que disuelven o que van raspando las valvas para llegar a la parte blanda del organismo. Dos estrategias que disuaden tanto a los depredadores como a los organismos que forman perforaciones en ellos, son el desarrollo de hileras de espinas muy finas en el interior de la concha (rinconélidos, terebratúlidos) y la formación de espinas marginales en rinconélidos (Álvarez y Martínez-Chacón, 2009, p. 415).
La mayoría de los taxones aquí descritos se han registrado en ambientes de plataforma somera, con circulación abierta y buena oxigenación. De los taxones estudiados, Reticulariina, Linoproductus y Composita son géneros que se encuentran comúnmente asociados a rocas tipo wackestone-packstone, que representan facies de baja profundidad y con amplia disponibilidad de nutrientes.
Beresi et al. (2012) coinciden en que la abundancia de fauna bentónica evidencia condiciones ambientales en las que se presentan niveles normales de oxígeno y adecuados niveles de nutrientes. La fauna estudiada se caracteriza por la presencia de braquiópodos y de diversos grupos, los cuales son en su mayoría epibentónicos. Considerando las asociaciones fosilíferas y el tipo de litología, se infiere un ambiente postarrecifal con buenos niveles de oxigenación y disponibilidad de nutrientes. Se considera que la disponibilidad de nutrientes cambió a lo largo del tiempo, tomando en cuenta la marcada variabilidad de las tallas y de la presencia de diferentes tipos de lofóforo.
Es importante notar que en algunos niveles de caliza se encontró material fragmentario disperso con abundantes productélidos, los cuales se hallan por lo general en posición de vida o con ligeras inclinaciones. También se encuentran placas columnares de crinoides y briozoos (?Polypora sp., Archimedes sp.) del Misisípico-Pensilvánico y de ambientes de plataforma somera en el noroccidente de México (Ochoa y Sosa, 1993).
Entre los braquiópodos que se presentan en la región, los estrofoménidos (schuchertélidos) son escasos. Los braquiópodos de este tipo presentan conchas planoconvexas con costillas delgadas mejor adaptadas a aguas poco profundas de circulación baja a moderada, prefiriendo zonas de poca perturbación por acción del oleaje. Se infiere que la presencia de espinas de equinodermos evidencie zonas de quietud o menor energía que se encontraban cercanas a zonas de mayor energía. Ambientes de este tipo pueden tratarse de rampas de fondo irregular con asociaciones bióticas dispersas establecidas inicialmente en zonas de mayor estabilidad. De los briozoos encontrados son más comunes los fenestélidos; su relativa mayor abundancia en la parte alta de la sección indica que las rocas en las que están incluidos debieron formarse en ambientes de baja profundidad con remoción de material y un grado de turbidez bajo con niveles de salinidad que posiblemente variaron debido a que son tolerantes a estos cambios. La asociación de braquiópodos en esta parte de la sección estratigráfica denota un ambiente intermareal a submareal somero con la presencia de restos de gasterópodos acumulados en posibles ambientes mareales de circulación abierta bajo la acción de corrientes, por encima de la base del oleaje (Kidwell et al., 1986). Otra explicación posible a esta acumulación de conchas se debió a la disminución en la tasa de sedimentación con la cual los elementos esqueletales acumulados podrían tener algunos miles de años de diferencia (Flessa y Kowalewski, 1994; Kidwell et al., 1986; Kowalewski et al., 1998).
Pérez-Huerta y Sheldon (2006) mencionan que los ejemplares con más de 20 mm de la especie Kozlowskia splendens se desarrollaron en facies profundas, mientras que los de tallas menores se asocian a facies de agua somera, esto sugiere una disposición diferencial de nutrientes y variaciones en la eficiencia de aprovechamiento del alimento. Por su parte, Olszewski y Patzkowsky (2001a, b) mencionan a Kozlowskia del Pensilvánico-Pérmico para facies de aguas profundas en asociación con braquiópodos de los géneros Wellerella, Lissochonetes, Retaria, Kozlowskia, Hystriculina, Rhipidomella, Crurithyris y Hustedia.
Kues (2009) infiere que Composita es un organismo filtrador de agua poco profunda y con alta tasa de productividad primaria, ya que posee un braquidio de tipo espiralium que le confiere una mayor eficiencia para capturar el alimento. Composita y otros athíridos presentan rangos de tallas que dependen de la profundidad donde vivieron, no obstante, en algunas especies se observa una ocurrencia preferencial en ambientes de baja profundidad, como sucede con Composita subquadrata (Pérez-Huerta y Sheldon, 2006). La variabilidad morfológica de algunos taxones está directamente influenciada por factores del medio, por ejemplo, Schumann (1991) demuestra que las conchas de Terebratalia transversa presentan charnela amplia, con costillas radiales bajo una morfología “tipo espiriférida” a medida que las corrientes son menos fuertes. El autor indica también que la presencia de formas con pedúnculos pequeños está asociada a condiciones de mayor influencia hidrodinámica (mayor intensidad de corriente), pese a que la propia variabilidad de T. transversa, en ambientes de mayor energía, se vuelve también mayor, pero con un predominio de formas con costelación tenue o que incluso la pierden.
Las especies Anthracospirifer occiduus y A. opimus se han encontrado en formaciones caracterizadas por litologías contrastantes. Lane (1964, p. 783) indica que la asociación de A. occiduus con Orthotetes, Linoproductus y Antiquatonia, se encuentra mayormente en caliza arcillosa mientras que A. opimus, Orthotetes, Reticulariina, Millerella, Endothyra, Tetrataxis, corales rugosos, briozoos y trilobites son predominantes en caliza pura. La heterogeneidad de formas puede asociarse a una amplia variedad de condiciones ecológicas y geográficas en la que los tipos de sustratos estarían asociados a diversas configuraciones de la cuenca. Además, la presencia de sustratos para el desarrollo de comunidades marinas con braquiópodos y otras formas de vida bentónicas, sésiles y epifaunales, requiere de rutas marinas para la dispersión durante sus estadios larvarios.
Los géneros de braquiópodos que se estudiaron en este trabajo son relativamente frecuentes en Norteamérica y Sudamérica para edades comprendidas entre el Pensilvánico medio-Pérmico (Cisuraliano). Esta tendencia se hace menos marcada para los géneros distribuidos en Europa y Asia, pero se sugiere la existencia de cierta migración de los braquiópodos hacia zonas de mayor estabilidad geográfica y ecológica.
Los sedimentos arcillosos o lodosos formaron barreras para la distribución de los organismos que se desarrollaban en sustratos firmes y fondos rocosos como Plicatifera, Kozlowskia, Linoproductus y Hustedia; Composita habría habitado en la cercanía de barreras arrecifales o zonas de playa y fondos lodosos. Finalmente, especies de Anthracospirifer y Reticulariina estarían relacionadas a zonas con aportes de sedimentos arenosos por acción de corrientes.
Otros factores ambientales se infieren por la biota asociada y por el tipo de sedimento. Buitrón-Sánchez, et al. (2007b) consideran que las calizas ricas en crinoides evidencian procesos de flujos de gravedad y tempestitas como lo consideran también Cook y Mullins (1983), Martin (1999) y Flügel (2010).
Consideraciones paleogeográficas
Durante el Paleozoico tardío, el Cratón Norteamericano presentaba fluctuaciones en la circulación oceánica debido al cierre de las aguas ecuatoriales que formaban mares epicontinentales. En el área de estudio existen también elementos estratigráficos, paleontológicos y paleogeográficos relacionados con cambios en las condiciones de sedimentación, tales como la presencia de estratos de caliza con gasterópodos y los de caliza arenosa formadas en aguas poco profundas y calizas rosáceas de mayor profundidad con fauna de braquiópodos en posición de vida y escasos restos de briozoos y placas desarticuladas de crinoides. De las variaciones en la fauna y la presencia de diferentes litologías a lo largo de la columna estratigráfica, se podrían inferir diversas causas pero que requerirían de su evaluación: 1) El aporte de material continental a través de canales de aguas superficiales con formación de deltas o planicies mareales sobre las que se desarrollaban comunidades bentónicas de aguas someras; 2) la afectación de la zona de estudio debido a control tectónico y/o magmatismo con deformación por esfuerzos; 3) el levantamiento de la zona por basculamiento asociado a fallas y la formación de relieves topográficos positivos, y 4) la disminución del nivel del mar asociado a cambios climáticos de nivel local, regional y/o global.
Aunque gran parte de la fauna estudiada en este trabajo refleja ambientes someros con energía moderada, es importante tomar en cuenta que el contexto paleogeográfico durante el Carbonífero-Pérmico del noroccidente de México estaba dominado por plataformas de agua somera y baja energía, con dominancia de masas de agua que formaban el océano Rhéico y que se encontraban condicionadas a bloques y/o terrenos sobre los que se desarrollaron este tipo de ambientes.
Las reconstrucciones paleogeográficas del Misisípico-Pensilvánico para el suroccidente de Estados Unidos de Norteamérica y el noroccidente de México, sugieren que esta región se encontraba en una latitud ecuatorial donde se formaban plataformas incididas por cañones submarinos, con influencia tectónica debido a la Orogenia Antler de principios del Carbonífero la cual propició el levantamiento continental en suroeste de EUA y noreste de Sonora (Ross, 1991; Ross y Ross, 1987).
Modelos paleogeográficos para el Pensilvánico temprano muestran que en la región de Norteamérica existían amplias plataformas marinas con circulación abierta y presencia de zonas con aporte diferencial de terrígenos así como zonas de circulación restringida en las que se producía dolomita asociados a incrementos en la sedimentación más que a disminuciones del nivel eustático (Haq y Schutter, 2008; Ross y Ross, 1987; Stevens y Stone, 2007; Stone et al., 2013).
Concluyendo, se describieron 7 especies de braquiópodos correspondientes a Plicatifera sp., Kozlowskia capaci., Linoproductus sp. cf. L. eastoni, Composita sp. cf. C. subtilita, Anthracospirifer occiduus, Hustedia mormoni y Reticulariina sp. que se incluyen en las familias Productellidae Schuchert y Le Vene, 1929, Linoproductidae Stehli, 1954, Athyrididae Davidson, 1881, Spiriferidae Carter, 1994), Retziidae Waagen, 1883 y Reticulariidae Waterhouse, 1978, con un registro estratigráfico del Misisípico tardío (Chesteriano) al Pérmico medio (Wordiano).
La asociación biótica registrada para la región de sierra Agua Verde es indicativa de un ambiente de plataforma somera, con desarrollo de un arrecife y del frente arrecifal con aguas bien oxigenadas y con niveles de energía diferenciados, pero mayormente moderada.
Para las especies identificadas se establecieron relaciones paleogeográficas para el Pensilvánico con otras regiones de México (Oaxaca, Hidalgo), Estados Unidos de América (Idaho, Wyoming, Nebraska, Nevada, Ohio, Kansas, Utah, Missouri, Pensilvania, Arizona, Nuevo México y Texas), Canadá, América del Sur (Bolivia, Perú), Europa (Irlanda, Bélgica) y Asia (Rusia, China, Japón, Indonesia).

