











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Diversos estudios han mostrado que las especies presentan patrones de distribución discontinuos, debido a la variación espacial en las condiciones que determinan la calidad del hábitat que ocupan (Santos y Tellería, 2006). Otro aspecto que puede afectar la distribución de las especies, es la interacción con otros organismos, por lo que la repartición de recursos en una comunidad es un factor determinante para la supervivencia de los individuos y para mantener la riqueza de especies (Ortega et al., 1982; Pianka, 1974). Sin embargo, la alteración de los hábitats afecta su heterogeneidad y la disponibilidad de recursos, lo cual puede tener un efecto en el comportamiento individual, así como en las relaciones espaciales de las especies (Barbault y Maury, 1981). La heterogeneidad ambiental, también puede afectar el acceso de los individuos a recursos como presas y microhábitat (Lattanzio y Miles, 2014).
Los estudios realizados en las comunidades de lacertilios, han mostrado que la calidad del hábitat es un factor determinante en su distribución, debido a que éste debe cubrir las necesidades primarias de los organismos, tales como refugio, alimento, sitios de asoleo y disponibilidad de parejas (Newbold y MacMahon, 2014; Trzcinski et al., 1999). Se ha mencionado que la abundancia de individuos en un área determinada, está relacionada con el número de microhábitats disponibles y que la elección del microhábitat por parte de las especies, puede moldear la estructura dentro de una comunidad (Mata-Silva, 2003; Smith y Fox, 2017).
En hábitats heterogéneos, las posibilidades de que las especies encuentren sitios de refugio o percha son mayores que en hábitats con características homogéneas o con menor número de sustratos. De igual forma, el número de microhábitats disponibles se ha relacionado con el grado de conservación del hábitat, ya que a mayor perturbación, el número de microhábitats disponibles disminuye (Mata-Silva, 2003; Pianka, 1974; Triska et al., 2017).
En siglos recientes, el ser humano ha realizado diversas actividades que provocaron la alteración y pérdida del hábitat de varias especies (Harte, 2007). Por ello, es necesario conocer los factores que influyen en su distribución y los aspectos que puedan amenazar su persistencia (Laurance, 2008; Triska et al., 2017). Por lo anterior, el propósito de este estudio fue conocer la organización espacial de la comunidad de lacertilios diurnos del valle de Zapotitlán Salinas, Puebla, mediante un análisis de topología de redes.
En biología, el uso de los métodos de análisis de redes ha estado enfocado principalmente a estudios entre especies que presentan interacciones de mutualismo (Bascompte et al., 2003; Schlueuning et al., 2014; Tohu et al., 2015), depredación (Dunne et al., 2002; Villa-Galaviz et al., 2012) y parasitoidismo (Martínez-Falcón et al., 2010). Además, las redes se han utilizado para evaluar el efecto de la fragmentación del hábitat (Cagnolo y Valladares, 2011). Sin embargo, los estudios que tratan de explicar la preferencia y uso de recursos por parte de los animales, aún son escasos (Araújo et al., 2008; Marquitti et al., 2014).
Materiales y métodos
El estudio fue realizado en Zapotitlán Salinas, dentro de la Reserva de la Biosfera Tehuacán-Cuicatlán (Fig. 1). El área se ubica en la parte sudeste del estado de Puebla, entre los 18°18’ y 18°19’ N, 97°28’ y 97°30’ O con un área aproximada de 1,004 km2, la altitud varía de los 1,242 a los 2,800 m snm (Montoya-Ayala et al., 2004). El área se caracteriza por la presencia de mezquitales, en los que predomina el mezquite (Prosopis laevigata) y matorrales espinosos caracterizados por tetechos (Neobusbania tetetzo), yucas (Yucca carnerosana) y sotolines (Beucarnea gracilis), entre otras especies vegetales (Dávila et al., 2002; Dávila-Aranda y Lira-Saade, 2003; Valiente-Banuet et al., 2009). La temperatura media anual es de 21.2 °C y la precipitación anual, de 380 mm3 (Arias-Toledo et al., 2000).
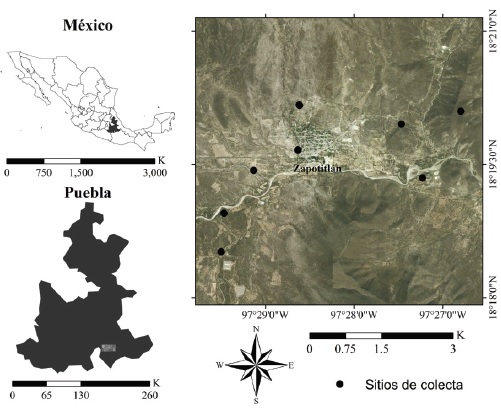
Los muestreos se realizaron mensualmente, de noviembre de 1997 a noviembre de 1998, en 8 sitios elegidos al azar en el valle de Zapotitlán Salinas, con una superficie de 2 ha cada uno (Fig. 1). La búsqueda de los organismos se llevó a cabo en los 8 sitios, con la participación de 3 personas, durante 2 horas, entre las 9:00 y 14:00 h. En cada sitio se registró el número de especies encontradas, el número de individuos por especie y el sustrato sobre el cual fueron observados por primera vez (Tabla 1).
Tabla 1 Sustratos utilizados por los lacertilios de Zapotitlán Salinas, Puebla.
| Sustrato | Descripción |
|---|---|
| Roca | Roca de cualquier material y tamaño |
| Arbusto | Arbusto de menos de 50 cm de alto |
| Mezquite | Mezquite de más de 50 cm de alto |
| Suelo | Suelo descubierto |
| Agave | Pencas de agave |
| Sotolin | Tronco de Beucarnea gracilis |
| Columnar | Tronco de cactácea columnar |
| Pared arenisca | Pared de arenisca de grano fino formando barrancas |
| Pencas nopal | Pencas de nopal acumuladas en el suelo |
| Yuca | Tronco de Yucca carnerosana |
| Pared | Pared de construcción |
| Pasto | Suelo cubierto con pasto |
| Palo verde | Arbusto de palo de verde |
| Suelo con vegetación | Herbáceas en suelo descubierto |
| Huizache | Arbusto de huizache |
| Paja | Paja acumulada en el suelo |
| Poste madera | Poste de cerca de madera |
| Suelo río | Cauce seco del río |
| Mamilaria | Cactáceas en forma de globo |
Con los datos obtenidos, se realizó un estudio de redes de tipo bipartita, que se caracterizan por presentar 2 grupos de datos (especies-sustratos) y su patrón de asociación (Jordano et al., 2009). Se construyó una matriz de adyacencia L (lacertilios) × S (sustratos), con valores de 0 y 1, donde 0 indica que no existe asociación y 1 que hay una asociación entre sustrato y especie. La red cualitativa obtenida se analizó con el programa ANINHADO (Guimarães y Guimarães, 2006) para obtener el valor de anidamiento (NODF), que muestra la forma en que están relacionados o conectados las especies y los sustratos en la red. Valores de anidamiento cercano a 1 indican que existe un patrón de anidamiento alto, mientras que valores cercanos a 0, indican que no existe anidamiento y que los datos pueden estar formando grupos o módulos (Jordano et al., 2009). Para establecer si los valores de anidamiento obtenidos son mayores que los esperados por azar, se llevó a cabo la comparación del grado de anidamiento obtenido con 1,000 iteraciones, utilizando el modelo nulo CE o modelo nulo 2 por medio del programa NODF (Almeida-Neto et al., 2008; Guimarães y Guimarães, 2006; Martínez-Falcón et al., 2010; Pagel et al., 2014).
Además, se generó una red cuantitativa, que incluye datos sobre la intensidad de asociación, con la cual se calcularon los siguientes parámetros estructurales: la conectividad (C), que se refiere a la fracción de interacciones registradas relativa al total de las interacciones posibles, su valor va de 0 a 1; la interacción de la uniformidad (IE), que indica la equidad de las interacciones; la vulnerabilidad (V), que es el número efectivo de especies por sustrato; la generalidad (G), que indica la media efectiva del número de sustratos por especie de lacertilio; la modularidad (M), que es una medida de la estructura de las redes, el número de módulos (NM) y la selectividad de la red (H2), como una medida de especialización. Estos índices se calcularon utilizando el paquete BIPARTITE 2.08 (Dorman et al., 2008; Morris et al., 2014; Pires et al., 2014; Villa-Galaviz et al., 2012).
La modularidad se calculó mediante el índice M (intervalo: 0-1) con base en el algoritmo de Newman y Girvan en el programa MODULAR (Marquitti et al., 2014) y su nivel de significancia se calculó utilizando 1,000 iteraciones, con el método de Monte Carlo.
Resultados
Durante la realización del estudio, se observaron 8 de las 15 especies de lacertilios que han sido registradas en la zona de estudio (Canseco-Márquez y Gutiérrez-Mayén, 2006), repartidas en 19 sustratos (Tabla 2).
Tabla 2 Especies y sustratos analizados durante el presente estudio, en Zapotitlán Salinas.
| Especies | Número | Sustratos | Número |
|---|---|---|---|
| Sceloporus jalapae | 127 | Roca | 63 |
| Sceloporus gadoviae | 41 | Arbusto | 51 |
| Phyllodactylus bordai | 28 | Mezquite | 32 |
| Aspidoscelis parvisocius | 18 | Suelo | 18 |
| Aspidoscelis sackii | 16 | Agave | 18 |
| Sceloporus horridus | 8 | Sotolín | 15 |
| Anolis quercorum | 3 | Columnar | 9 |
| Urosaurus bicarinatus | 2 | Pared de arenisca | 9 |
| Pencas de nopal | 7 | ||
| Yucca | 5 | ||
| Pared de construcción | 3 | ||
| Pasto | 3 | ||
| Palo verde | 2 | ||
| Suelo con vegetación | 2 | ||
| Huizache | 2 | ||
| Paja | 1 | ||
| Poste de madera | 1 | ||
| Suelo de rio | 1 | ||
| Mamilaria | 1 |
El análisis de anidamiento mostró que existen 45 asociaciones entre especies y sustratos, con un promedio de 1.66 interacciones por especie (Tabla 3). El valor de anidamiento de la matriz cualitativa fue de NODF = 45.72 (p = 0.001), que es significativamente diferente del obtenido con las 1,000 iteraciones (NODFce = 36.6, p = 0.07). Por otra parte, el valor de anidamiento usando la matriz cuantitativa fue de NODF = 43.95 (p = 0.0001). El análisis mediante el programa BIPARTITE mostró que la estructura de la red obtenida, es alta (Fig. 2). Dentro de la red se observa que Sceloporus jalapae y Aspidoscelis parvisocius, son las especies que presentan un número mayor de asociaciones, con 14 y 7, respectivamente (Figs. 2, 3). El resto de las especies presentaron un número menor de sustratos ocupados (Tabla 3).
Tabla 3 Parámetros de la red. Valores significativos corresponden a p < 0.05.
| Valor | p | |
|---|---|---|
| Número de lacertilios | 8 | |
| Número de sustratos | 19 | |
| Número de asociaciones | 45 | |
| Media del número de interacciones | 1.66 | |
| NODF (cualitativo) | 45.72 | 0 |
| NODF (cuantitativo) | 43.95 | 0.0001 |
| NODF (ce) | 36.6 | 0.07 |

Figura 2 Presencia de las especies por tipo de sustrato. A = Número de interacciones entre especies y sustratos (negro: presencia de interacción, blanco: ausencia de interacción); B = interacciones entre especies y sustratos (la intensidad del color indica la intensidad de la asociación).
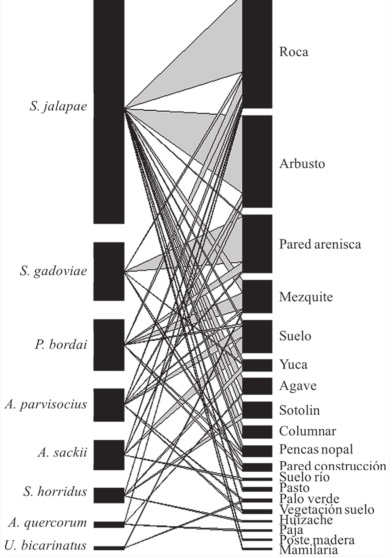
Figura 3 Especies de lacertilios y sustratos presentes en el área de estudio, las líneas representan las interacciones y el grosor la importancia de las mismas.
Por otra parte, el análisis reveló un valor de modularidad de 0.36, mientras que el valor de conectividad fue de 0.29. La interacción de la uniformidad fue de 0.60, la vulnerabilidad de 2.28, la generalidad de 4.98 y el solapamiento de nicho entre las especies fue de 0.24 (Tabla 4). Además, se detectó la existencia de 5 módulos (Fig. 4).
Tabla 4 Valores de los atributos de la red (valores significativos corresponden a p < 0.05).
| Valor | p | |
|---|---|---|
| Modularidad (M) | 0.36 | 0.65 |
| Número de módulos (NM) | 5 | |
| Conectividad (C) | 0.30 | 0.0001 |
| Interacción de la uniformidad (IE) | 0.60 | |
| Generalidad (G) | 4.98 | |
| Especialización (H2) | 0.44 | |
| Vulnerabilidad (V) | 2.28 | |
| Solapamiento de nicho de | 0.24 | |
| escamados | ||
| Solapamiento de nicho de | 0.34 | |
| sustratos | ||
| Robustez | 0.67 |

Figura 4 Módulos formados entre los elementos de la red (la intensidad del color indica la intensidad de la asociación).
En cuanto a las interacciones con mayor incidencia e importancia se encuentran las de S. jalapae y su asociación con las rocas y arbustos, seguida de la asociación mostrada por S. gadoviae con las paredes de arenisca (Fig. 3).
Discusión
Aunque se han registrado 15 especies de lacertilios en el valle de Zapotitlán Salinas, las 8 especies encontradas en el estudio (Tabla 1), son las especies diurnas más comunes y abundantes en la región (Canseco-Márquez y Gutiérrez-Mayen, 2006). El no haber localizado a todas las especies registradas para la zona, puede deberse a que la distribución de los lacertilios dentro del valle no es uniforme. Se ha mencionado que la distribución de las especies depende de la estructura en forma de parches del paisaje, así como de su grado de alteración (Mata-Silva, 2003). Además, existen especies como Xenosaurus rectocollaris que ocupan zonas muy restringidas dentro del área de estudio. Esta especie en particular, se localiza en las zonas altas de los cerros El Pajarito y Chacatecas, que no fueron incluidos en los sitios de muestreo (Canseco-Márquez y Gutiérrez-Mayén, 2006).
El valor de anidamiento obtenido sugiere que existe un patrón de anidamiento significativo entre los elementos de la red. En varios estudios se ha visto que el anidamiento depende de las interacciones entre especies generalistas y especialistas. En una red anidada, existe una jerarquización de las interacciones (Guimarães et al., 2016) y se observa un patrón en donde las especies generalistas interactúan con las especies especialistas y otras generalistas, pero las especialistas no interactúan entre sí (Britton et al., 2015). Además, el anidamiento depende de la riqueza de elementos en la matriz (Bascompte et al., 2003).
La red obtenida muestra un patrón asimétrico en la especialización de las asociaciones de los lacertilios con los sustratos (Fig. 3). Los resultados indican que la especie que se presentó con mayor frecuencia en los sitios visitados (S. jalapae), mostró un comportamiento generalista con relación a los sustratos utilizados, mientras que otras especies tuvieron preferencias más específicas (Fig. 2).
Sceloporus jalapae es una especie de lacertilio terrestre que puede explotar diferentes tipos de sustrato para llevar a cabo sus actividades. Sin embargo, los resultados muestran que a pesar de utilizar hasta 14 tipos de sustrato (Fig. 2A), los individuos de la especie presentan una interacción mayor con los arbustos y las rocas, y en menor grado con el pasto o las mamilarias (Fig. 2B). Es común en regiones de pendiente baja y es abundante debajo de los arbustos y en la base de diferentes plantas de tronco alto (Canseco-Márquez y Gutiérrez-Mayén, 2006; González-Espinoza, 2006).
El análisis mostró que la mayor parte de las especies de lacertilios no tienen restricciones en la elección de los sustratos que utilizan para realizar sus actividades (Fig. 2). Sin embargo, se observa que existen asociaciones importantes entre ciertas especies y sustratos (Fig. 2B), como en el caso de S. gadoviae, que está asociada a paredes de arenisca. Los hábitos de esta lagartija la ubican en lugares rocosos o en paredes de tierra formadas por la erosión (Canseco-Márquez y Gutiérrez-Mayén, 2006; Mata-Silva, 2003; Woolrich-Piña et al., 2005). S. gadoviae es una especie trepadora, aunque durante el estudio se observó a varios individuos en las partes bajas, asoleándose o en busca de alimento (Fig. 2).
Por otra parte, los resultados obtenidos indican que no existe una modularidad significativa dentro de la matriz (Tabla 3). Lo anterior puede deberse a que en el valle de Zapotitlán Salinas existe un número amplio de microhábitats y a que la mayor parte de las especies de lacertilios elige varios tipos de sustratos (Fig. 2). Una modularidad alta significa que dentro de la red, existen subconjuntos de especies que de manera frecuente eligen ciertos sustratos, pero pocas veces utilizan sustratos que están fuera del módulo del que forman parte.
En lacertilios, esto podría estar influenciado por la filogenia, ya que se ha documentado que el uso de los sustratos está correlacionado con diversas características morfológicas que tienen relación con la filogenia. Aunque también existe la posibilidad de que especies con caracteres morfológicos conservados, utilicen una gama amplia de sustratos (Tulli et al., 2012).
El análisis de modularidad mostró la existencia de 5 grupos constituidos por las especies bajo estudio. Tres de ellos están conformados por una sola especie (Anolis quercorum, Sceloporus jalapae y S. horridus), lo que indica que a pesar de encontrase en un número amplio de sustratos, presentan preferencia por un número reducido de ellos. El resto de las especies, están formando módulos que muestran su preferencia por sustratos similares (Fig. 4). A pesar de lo anterior, la competencia no parece ser un factor determinante en la repartición de los sustratos, diversos estudios han mostrado que las condiciones microambientales, juegan un papel importante en la elección de los sustratos por parte de los lacertilios. Se ha visto que en las comunidades de lagartijas del género Anolis, la repartición de sustratos está relacionada con la fisiología térmica de las especies (Buckley y Roughgarden, 2005). Además, los lacertilios pueden compartir sustratos sin que haya competencia, al utilizar este recurso en diferentes momentos del día o en diferentes temporadas (ej. Sceloporus gadoviae y Phyllodactylus bordai; Canseco-Márquez y Gutiérrez-Mayén, 2006).
Existen especies que parecen tener mayor especialización, pero el número de asociaciones detectadas no permite ser concluyente en cuanto a sus preferencias de sustrato. Urosaurus bicarinatus fue la especie observada con menor frecuencia (Tabla 2), ya que solo se encontraron 2 individuos, asociados a arbustos de mezquite y al palo verde. Aunque un mayor número de registros permitiría precisar las preferencias de U. bicarinatus, otros estudios realizados en la zona, indican que se trata de una especie escaza (Canseco-Márquez y Gutiérrez-Mayén, 2006; Mata-Silva, 2003).
En el caso de los lacertilios del género Aspidoscelis, ambas especies (A. parvisocius y A. sackii) presentan territorios de actividad extensos, con solapamiento amplio entre ellos. El sustrato principal de estas especies fue el suelo, ya que normalmente se les encuentra desplazándose en busca de alimento, debido a que son especies de forrajeo activo (Woolrich-Piña et al., 2011). Sin embargo, se ha observado que las especies de Aspidoscelis pueden utilizar microhábitats diferentes, para llevar a cabo sus distintas actividades (Anderson, 1993).
La conectividad de la red obtenida fue baja, lo cual indica una estabilidad alta dentro de la red y evita que se produzcan efectos negativos o colapso entre los nichos por posible extinción de las especies (Cagnolo y Valladares, 2011; Dunne et al., 2002; Villa-Galaviz et al., 2012). Se ha mencionado que las redes que presentan baja conectividad se producen cuando existe competencia intensa entre las especies por los recursos (Villa-Galaviz et al., 2012). Sin embargo, el valor de solapamiento de nicho de los lacertilios de Zapotitlán Salinas es bajo (Tabla 3), lo cual indica que la competencia por los sustratos es baja.
En las comunidades de lacertilios, la repartición de recursos está mediada por la conducta, temporalidad y espacialidad de los taxones, evitando así, la competencia entre ellos (Sillero y Gomes, 2016). En la zona de estudio, pueden observarse individuos de las distintas especies en áreas pequeñas, ocupando los diferentes sustratos. Además, se han observado individuos de especies como Sceloporus jalapae y S. horridus compartiendo el mismo sustrato (yuca o sotolín), sin embargo, la primera se encontró en la base y la segunda en la parte más alta de las plantas. Otro caso similar es el que se presenta entre S. gadoviae y Phyllodactylus bordai, ya que ambos están asociados a las paredes de arenisca, pero la actividad de S. gadoviae es diurna, mientras P. bordai tiene actividad nocturna (Canseco-Márquez y Gutiérrez-Mayén, 2006).
El valor de la interacción de la uniformidad (IE), obtenido para la red del presente estudio es bajo (Tabla 4), lo cual indica que los lacertilios de Zapotitlán muestran niveles heterogéneos de asociación con los sustratos (Blüthgen et al., 2008; Schlueuning et al., 2014).
Por otra parte, el valor de la especialización (H2) es bajo, lo cual puede deberse a que existen especies que se asocian con pocos sustratos, ya que valores más bajos de este parámetro, indican una especialización global de las asociaciones representadas en la red (Jordano et al., 2009). Dentro de la red generada, existen sustratos como las mamilarias y los postes de madera, que solo fueron elegidos por una especie. La selección de sustratos es importante para la estructura de la comunidad, ya que especies generalistas, coexisten con aquéllas que tienden a ser más especialistas, utilizando sustratos que no usan las que tienen preferencias más restringidas (Smith y Fox, 2017).
Los valores de especialización, son congruentes con los valores de generalidad (4.98) y de vulnerabilidad (2.28), que representan la media de asociaciones efectivas por lacertilio y por sustrato, respectivamente (Blüthgen et al., 2008; Morris et al., 2014). La generalidad indica que los lacertilios pueden elegir varios sustratos, mientras la vulnerabilidad sugiere que los sustratos, son seleccionados por pocas especies.
El análisis realizado indica que en la comunidad de lacertilios diurnos de Zapotitlán Salinas, las especies utilizan los sustratos disponibles de forma diferenciada. La heterogeneidad del paisaje proporciona una variedad de sustratos que son importantes para la presencia y distribución de los lacertilios en el área.

