











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La heterostilia es un polimorfismo floral que se caracteriza por poseer 2 (distílicas) o 3 (tristílicas) formas florales (morfos) que difieren en la longitud del estigma y de las anteras de sus flores (Barrett, 1992; Barrett y Shore, 2008; Ganders, 1979; Price y Barrett, 1984). En las plantas distílicas, las flores del morfo Pin poseen el estigma por encima de las anteras y las flores del morfo Thrum poseen las anteras por encima del estigma (Barrett, 1992; Darwin, 1877; Ganders, 1979; Hernández-Ramírez, 2012; Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004). Este polimorfismo floral usualmente es acompañado por un sistema de autoincompatibilidad intramorfos y una serie de rasgos florales accesorios que difieren entre morfos florales (tamaño de las flores y del lóbulo del estigma, así como número y tamaño de los granos de polen; Barrett, 1992; Barrett y Shore, 2008; Ganders, 1979; Ree, 1997). Debido al sistema de auto-incompatibilidad intramorfos, únicamente el polen legítimo proveniente del morfo opuesto es capaz de fertilizar los óvulos y producir semillas viables (Barrett, 1992; Barrett y Shore, 2008; Ganders, 1979).
Darwin (1877) sugirió que el significado adaptativo de este polimorfismo estigma-antera estaba estrechamente relacionado con la promoción del flujo de polen legítimo entre morfos a través de un ajuste entre la morfología floral y el cuerpo del polinizador (Barrett, 1992; Barrett y Shore, 2008; Ganders, 1979; Hernández y Ornelas, 2007a; Ornelas, Jiménez et al., 2004). Estudios posteriores mostraron que el comportamiento de los visitantes florales repercute en la transferencia de polen legítimo entre morfos a través de la frecuencia de visita, el patrón diurno de llegada a las flores y el movimiento de los polinizadores dentro y entre plantas diferentes (Hernández-Ramírez, 2012; Price y Barrett, 1984; Stone, 1996). A pesar de existir gran número de estudios en los cuales se explora el significado adaptativo del polimorfismo estigma-antera en plantas heterostílicas (Barrett, 1992; Barrett y Shore, 2008; Ganders, 1979; Ree, 1997; Stone, 1996), pocos estudios han abordado la dimensión temporal en el estudio de la interacción planta-animal y sus posibles efectos sobre el potencial reproductivo de los morfos, excepto Price y Barrett (1984) y Wolfe y Barrett (1988).
Palicourea padifolia (Roem. y Schult.) C. M. Taylor y Lorence (Rubiaceae) es una especie distílica que presenta flores nectaríferas tubulares que son visitadas principalmente por colibríes, seguidas de llegadas ocasionales de abejas, abejorros y mariposas (Ornelas, Jiménez, 2004; Ree, 1997). En este sistema, los colibríes han sido identificados como el grupo de “polinizadores eficientes” con base en una serie de estudios empíricos y experimentales (Hernández y Ornelas, 2007a; Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004). No obstante, se desconoce el papel que desempeña el grupo de invertebrados en la transferencia legítima de polen entre morfos en P. padifolia.
El objetivo de este estudio fue investigar la variación temporal en el gremio de visitantes florales y el patrón del flujo de polen en la especie distílica Palicourea padifolia (Rubiaceae) durante un ciclo anual de floración. La hipótesis a probar fue que los colibríes representan el grupo de “polinizadores eficientes”, los cuales promueven la transferencia de polen legítimo entre morfos. Aunado a lo anterior, se probó la hipótesis del papel marginal y no significativo en la transferencia de polen entre morfos mediado por insectos en la especie, de acuerdo con Hernández y Ornelas (2007a); Ornelas, González et al. (2004) y Ornelas, Jiménez et al. (2004). Para ello, se estudió el comportamiento de forrajeo de los visitantes florales y se contrastó con las cargas de polen legítimo depositados en el estigma de las flores.
Materiales y métodos
El área de estudio se localiza en el bosque mesófilo de montaña que se distribuye en la región central del estado de Veracruz, México (19˚30’ N, 96˚57’ O; 1,225 m snm). La temperatura promedio anual es de 20˚C, con una precipitación anual que oscila entre los 1,500 y 1,900 mm. El trabajo de campo se realizó en el 2014 durante el ciclo anual de la floración de la especie.
Palicourea padifolia (Roem. and Schult.) C. M. Taylor y Lorence (Rubiaceae) se distribuye de la sierra Madre Oriental en México hasta Panamá (Taylor, 1989). Es una especie distílica, auto-incompatible con una reciprocidad estigma-antera entre morfos imperfecta (Hernández y Ornelas, 2007a, b; Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004; Ree, 1997). El morfo Pin y Thrum producen un número similar de inflorescencias, botones florales y flores de un día de longevidad (Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004). Las flores del morfo Thrum producen una menor cantidad de granos de polen, pero de mayor tamaño con respecto al morfo Pin (media ± ee; número de granos de polen para Pin: 335.1 ± 10.4 y Thrum: 267.1 ± 11.2; tamaño de granos de polen para Pin: 99.9 ± 0.6 µm y Thrum: 65.4 ± 0.5 µm; Ornelas, Jiménez et al., 2004). Lo anterior hace posible diferenciar los granos de polen provenientes de cada uno de los morfos en el estigma de las flores. Hernández y Ornelas (2007a) demostraron a través de diversos experimentos con flores emasculadas e intectas que la carga polínica estigmática es el resultado de la actividad de forrajeo de los visitantes florales en la especie y población estudiada. Detalles específicos del área de estudio y de la población muestreada se pueden consultar en Ornelas, González et al. (2004); Ornelas, Jiménez et al. (2004), así como en Hernández y Ornelas (2007a, b).
Se realizaron 3 muestreos a lo largo del año los cuales aseguraron la independencia de las observaciones entre sí (margen de 30 días de diferencia entre muestreos). El muestreo abarcó el periodo completo de la floración de la especie en el área de estudio y el año de muestreo. Debido a que la estación de floración inicia en marzo y finaliza en septiembre, se muestrearon los meses de marzo (inicio del periodo de floración), junio (pico de floración) y septiembre (fin del periodo de floración). Se seleccionaron al azar y se marcaron 30 plantas del morfo Pin y 30 plantas del morfo Thrum. En cada una de las etapas del muestreo se repitieron los mismos procedimientos con las mismas plantas focales. Dos personas realizaron el registro de los visitantes florales de forma simultánea previa aleatorización en el orden de muestreo de las plantas y de los morfos.
Se estimó la frecuencia de la presencia de los morfos a través del muestreo de todas las plantas encontradas en floración a lo largo del transecto que atraviesa la población (Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004). Se contabilizó el número total de inflorescencias por planta al inicio, pico y final del periodo de floración de P. padifolia.
El periodo de observación y el registro de los visitantes florales inició a las 08:00 hrs y finalizó a las 18:00 hrs. Para cada planta focal se registró la especie de visitante floral, hora de visita y si colectaba néctar y/o polen (Hernández-Ramírez, 2012; Ornelas, González et al., 2004). La actividad de forrajeo de los visitantes florales dentro y entre plantas focales incluyó el siguiente registro: a) voló dentro de la planta focal, b) voló a otra planta del morfo opuesto, c) voló a otra planta del mismo morfo y d) voló fuera del rango de observación (Hernández-Ramírez, 2012). El mismo protocolo de muestreo se realizó al inicio, pico y final del periodo de floración de P. padifolia. Los visitantes florales se agruparon en gremios (himenópteros, lepidópteos y troquilinos) y se procedió al análisis de datos.
Se aplicaron análisis de supervivencia para calcular la probabilidad de que las flores del morfo Pin y Thrum fueran visitadas en cada una de las etapas de muestreo (Muenchow, 1986). El inicio del periodo de observación (08:00 hrs) se consideró como tiempo cero y se contabilizaron los minutos trascurridos desde el inicio de la observación hasta la llegada del visitante floral (Muenchow, 1986). Se aplicó la prueba de Mantel-Cox para detectar diferencias significativas en la probabilidad de que las flores del morfo Pin y Thrum fueran visitadas por el gremio de visitantes florales en cada etapa de muestreo (Muenchow, 1986). Se realizaron pruebas de chi-cuadrado (χ2) con la finalidad de probar la independencia del comportamiento de forrajeo del gremio de visitantes florales dentro y entre plantas focales (Hernández-Ramírez, 2012).
En cada etapa de muestreo se seleccionaron al azar 2 inflorescencias por planta focal. Una inflorescencia se embolsó como control y la otra se mantuvo expuesta a los visitantes florales (Hernández y Ornelas, 2007a, b). A las 19:00 hrs se recolectaron 10 flores por inflorescencia y se trasladaron al laboratorio en cajas Petri. Se seleccionó al azar una flor por inflorescencia, se removió el estigma de las flores y se fijó la muestra en un portaobjetos (Hernández y Ornelas, 2007a). Con la ayuda de un microscopio compuesto equipado con un micrómetro (Zeiss, Oberkochen, Alemania), se contabilizó el número de granos de polen de los morfos Pin y Thrum presentes en cada uno de los estigmas de las flores (Hernández y Ornelas, 2007a ; Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004). Este mismo protocolo se realizó al inicio, pico y final del periodo de floración de la especie.
Se aplicaron modelos lineales generalizados para examinar las diferencias en el número de granos de polen y en la proporción de granos de polen legítimos en el estigma de las flores entre morfos y entre estaciones de floración (Crawley, 2005). Se ajustó un error tipo Poisson (log) para el número de granos de polen y un error tipo binomial para la proporción de granos de polen legítimos en el estigma de las flores (variable compuesta conformada por el número total de granos de polen y el número de granos de polen legítimos; Crawley, 2005). Los análisis se realizaron en el programa R (R Development Core Team, 2017).
Resultados
La proporción de morfos no difirió de la relación 1:1 (156 Pin:160 Thrum). Ambos morfos produjeron un número similar de inflorescencias a lo largo del ciclo anual de floración (10-12 inicio, 56-66 pico y 5-7 final del periodo de floración).
Se registraron 441 visitas a las flores de P. padifolia en 1,800 horas de observación (232 visitas al morfo Pin y 209 visitas al morfo Thrum). En total, 15 especies de invertebrados y 5 de vertebrados visitaron las flores de P. padifolia, los cuales hicieron contacto con el estigma y/o las anteras de las flores (Tabla 1). Los himenópteros o abejas (sensu lato) Apis mellifera, Scaptotrigona mexicana y Bombus medius se observaron libando néctar y colectando polen de las anteras de las flores en cada una de sus visitas. Los lepidópteros (mariposas) y troquilinos (colibríes) únicamente libaron el néctar de las flores. Los lepidópteros fueron los visitantes florales más frecuentes (51%), seguidos de los troquilinos (36%) e himenópteros (13%).
Tabla 1 Proporción de visita y gremio de visitantes florales de Palicourea padifolia (Rubiaceae) por morfo y estación de floración.
| Visitantes florales | Inicio de la floración | Pico de la floración | Final de la floración | |||
| Pin | Thrum | Pin | Thrum | Pin | Thrum | |
| Apis mellifera | 2 | 2 | 3 | 1 | - | - |
| Bombus medius | 3 | 2 | - | - | - | - |
| Scaptotrigona mexicana | 6 | 5 | 3 | 4 | - | - |
| Visita relativa de himenópteros (%) | 20 | 11 | - | |||
| Aeria eurimedia pacifica | 3 | 3 | - | - | - | |
| Phoebis philea | 4 | 4 | - | - | - | |
| Chlosyne janais | 3 | 3 | 8 | 4 | - | - |
| Dione juno huascuma | 4 | 2 | - | - | - | - |
| Dione moneta poeyii | 3 | 3 | 3 | 4 | - | - |
| Enantia mazai mazai | 4 | 3 | - | - | - | - |
| Eresia phillyra | 4 | 4 | - | - | - | - |
| Heliconius charithonia | 3 | 3 | - | - | - | - |
| Anteos clorinde | 3 | 4 | 5 | 5 | - | - |
| Ithomia leila | 2 | 2 | 5 | 7 | - | - |
| Phoebis sennae marcellina | 3 | 4 | - | - | - | - |
| Phoebis neocypris virgo | 5 | 4 | - | - | - | - |
| Visita relativa de lepidópteros (%) | 80 | 41 | - | |||
| Atthis heloisa | - | - | - | - | 17 | 17 |
| Eugenes fulgens | - | - | - | - | 14 | 10 |
| Amazilia cyanocephala | - | - | 8 | 8 | - | - |
| Campylopterus curvipennis | - | - | 14 | 13 | 15 | 17 |
| Lampornis amethystinus | - | - | 2 | 3 | - | - |
| Vista relativa de colibríes (%) | - | 48 | 100 | |||
Los himenópteros y lepidópteros visitaron las flores de P. padifolia al inicio del periodo de floración, mientras que los troquilinos se integraron al ensamble de visitantes florales durante el pico de floración y fueron los únicos que permanecieron hasta el final del periodo de floración de la especie (Tabla 1).
El análisis de supervivencia mostró diferencias significativas en la probabilidad de que las flores de los morfos Pin y Thrum fueran visitadas al inicio y al pico del periodo de floración (Fig. 1). Al inicio del periodo de floración, los lepidópteros visitaron primero las flores del morfo Pin y más tarde visitaron a las de Thrum (Tabla 2). Del mismo modo, los troquilinos visitaron primero a las flores del morfo Pin y posteriormente a las de Thrum durante el pico de floración de la especie (Tabla 2). Las diferencias antes mencionadas se desvanecieron al final del periodo de floración, resultando en una probabilidad similar de que las flores de ambos morfos fueran visitadas por los troquilinos (Tabla 2; Fig. 1).
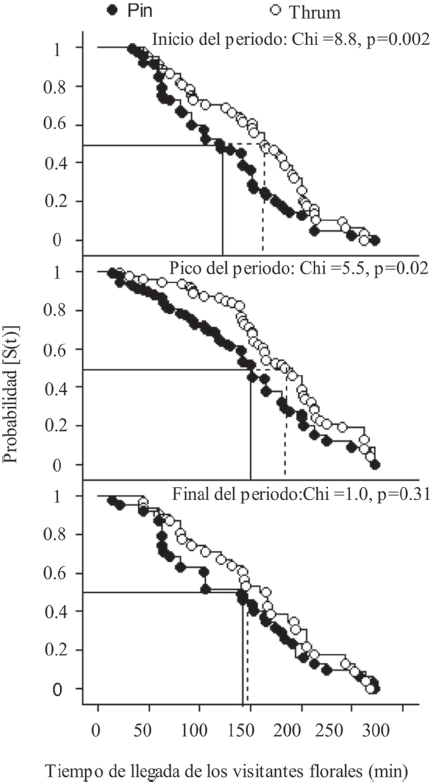
Figura 1 Comparación entre morfos de Palicourea padifolia (Rubiaceae) en la probabilidad de que las plantas sean visitadas por los visitantes florales durante los 300-min del periodo de observación por periodo de floración. La abreviación S(t) es la probabilidad de que un visitante floral aún no haya hecho la visita a la planta. El tiempo de llegada de los visitantes florales se refiere al tiempo trascurrido desde el inicio del periodo de observación.
Tabla 2 Tiempo de llegada del gremio de visitantes florales a las flores y cargas de polen en estigma por morfo y estación de floración en Palicourea padifolia (Rubiaceae). Los datos mostrados son media ± E. E. *p ≤ 0.01, **p ≤ 0.001, ***p ≤ 0.0001.
| Variables analizadas | Inicio de la floración | Pico de la floración | Final de la floración | |||
| Pin | Thrum | Pin | Thrum | Pin | Thrum | |
| A) Patrón diurno de visita (minutos) | ||||||
| Himenópteros (abejas) | 146.4 ± 11.4 | 154.8 ± 11.3 | 165.0 ± 53.5 | 180.9 ± 18.9 | - | - |
| Lepidópteros (mariposas) | 118.7 ± 7.3* | 156.5 ± 7.9 | 152.1 ± 10.2 | 160.6 ± 8.6 | - | - |
| Troquilinos (colibríes) | - | - | 147.2 ± 12.2* | 196.6 ± 10.6 | 134.6 ± 12.1 | 164.3 ± 12.5 |
| B) Cantidad de granos de polen depositados en el estigma de las flores | ||||||
| No. Total de granos de polen | 41.5 ± 2.3*** | 16.4 ± 1.8 | 62.6 ± 4.1*** | 48.5 ± 3.8 | 12.1 ± 1.6 | 8.0 ± 2.8 |
| Proporción de granos de polen legítimos |
0.19 ± 0.04 | 0.67 ± 0.05*** | 0.32 ± 0.02 | 0.55 ± 0.03*** | 0.29 ± 1.4 | 0.32 ± 2.0 |
Con respecto a la actividad de forrajeo, los lepidópteros realizaron un mayor número de movimientos dentro de las plantas al inicio del periodo de floración (χ2 = 6.43, p = 0.01; Fig. 2A). Cuando los lepidópteros se movieron entre plantas diferentes visitaron preferentemente a las plantas del mismo morfo (χ2= 7.22, p = 0.007; Fig. 2A). En contraste, los himenópteros se movieron entre plantas de morfos diferentes al inicio del periodo de floración (χ2= 8.34, p = 0.003; Fig. 2A).

Figura 2 Patrón de visita del gremio de visitantes florales de Palicourea padifolia (Rubiaceae) por morfo y estación de floración. Solo los datos estadísticamente significativos son mostrados (en porcentaje). La imagen identifica al gremio de visitante floral (mariposa: lepidópteros, abeja: himenópteros y colibrí: troquilinos). La flecha continua hace referencia a la dirección del movimiento de forrajeo del visitante floral dentro y entre plantas diferentes. La flecha punteada hace referencia al movimiento de forrajeo entre plantas diferentes, pero del mismo morfo.
Durante el pico de floración de la especie se registró la mayor diversidad taxonómica en el gremio de visitantes florales. El movimiento de forrajeo al interior de las plantas fue similar entre los lepidópteros, himenópteros y troquilinos (χ2= 0.58, p = 0.74; Fig. 2B). Por otro lado, los colibríes se movieron preferentemente del morfo Pin hacia Thrum, mientras que los lepidópteros se movieron entre plantas distintas pero del morfo Pin (χ2= 10.47, p = 0.005 y χ2= 9.63, p = 0.008; respectivamente, Fig. 2B). Al final del periodo de floración sólo los troquilinos visitaron las flores de la especie y su comportamiento de forrajeo no difirió dentro y entre plantas focales (χ2= 0.39, p = 0.82; Fig. 2C).
Durante el año de muestreo fue relativamente raro observar que los visitantes florales volaran fuera del rango de observación (< 5% de los eventos de visita) y en ninguno de los casos hubo diferencias estadísticas entre los visitantes florales (inicio: χ2= 0.53, p = 0.46, pico: χ2= 2.23, p = 0.32 y final: χ2= 0.07, p = 0.78).
Las flores del morfo Pin recibieron un mayor número de granos de polen (Pin: 31.15 ± 2.89, n = 90) que Thrum (27.00 ± 2.09, n = 90; χ2= 0.94, p = 0.036). No obstante, la mayor proporción de granos de polen legítimos (intermorfo) se observó en los estigmas de las flores de Thrum (Pin: 0.30 ± 0.03, n = 90 y Thrum: 0.51 ± 0.02, n = 90; χ2= 29.40, p < 0.0001). La mayor carga de polen se observó durante el pico de floración de la especie (χ2= 26.48, p < 0.0001 para el número de granos de polen y χ2= 30.94, p < 0.0001 para la proporción de granos de polen legítimos; Tabla 2). Las flores de Pin recibieron una mayor carga de polen (número de granos de polen) al inicio y en el pico de la floración de la especie (Tabla 2). En contraste, las flores de Thrum recibieron una mayor proporción de granos de polen legítimos (intermorfo) al inicio del periodo de floración, seguido del pico de floración de la especie (Interacción Morfo × periodo de floración; χ2= 20.3, p < 0.0001 para el número de granos de polen y χ2= 19.15, p < 0.0001 para la proporción de granos de polen legítimos; Tabla 2).
La carga de polen observada en el estigma de las flores se atribuyó a la actividad de forrajeo de los visitantes florales. Lo anterior es debido a la mínima cantidad de granos de polen depositados en el estigma de las flores excluidas (control; auto-deposición de polen) con respecto al tratamiento abierto a los visitantes florales (número de granos de polen para el tratamiento de exclusión 0.1 ± 0.001, n = 90 y para el tratamiento abierto 31.00 ± 2.19, n =90 abierto, χ2= 90.51, p < 0.0001; proporción de granos de polen legítimos para el tratamiento de exclusión 0.0 ± 0.0, n = 90 y para el tratamiento abierto 0.41 ± 0.003, n = 90 abierto; χ2= 74.51, p < 0.0001).
Discusión
P. padifolia exhibe un sistema de polinización generalizado, el cual está compuesto por un grupo taxonómicamente diverso de invertebrados y vertebrados que consumen el néctar y el polen de sus flores. Casos similares de polinización generalizada se han reportado para un gran número de especies distílicas de la familia Rubiaceae, incluyendo a Psychotria suerrensis (Stone, 1996) y Psychotria nervosa (Hernández-Ramírez, 2012).
Los lepidópteros fueron los visitantes florales más frecuentes, seguidos de los troquilinos e himenópteros. Esta relación de dominancia de invertebrados sobre vertebrados polinizadores había sido subestimada en la especie, dado que la mayoría de los muestreos se realizaron durante el pico de floración de P. padifolia; es decir, cuando la frecuencia de visita de los colibríes es relativamente mayor (Hernández y Ornelas, 2007a; Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004).
Durante el ciclo anual de floración de la especie se observó un sistema de polinización no-especializado y dinámico en el tiempo, el cual inició con la presencia de mariposas y abejas (lepidópteros e himenópteros); a los cuales se unió el grupo de colibríes (troquilinos) en el pico de floración de P. padifolia. En concordancia con estos resultados, Wolfe y Barrett (1988) reportaron variaciones en el gremio de visitantes florales a lo largo de la estación de floración de Pontederia cordata.Wolfe y Barrett (1988) observaron que la mayor diversidad de visitantes florales coincidió con en el pico de la floración de P. cordata. En P. padifolia, la mayor carga de polen depositado sobre el estigma de las flores coincidió con la mayor actividad y con el grupo más diverso de visitantes flores (durante el pico de floración). Price y Barrett (1984) también observaron un mayor número de granos de polen en el estigma de las flores asociado a la mayor actividad de los visitantes florales en Pontederia cordata.
Aunado a los cambios en la cantidad de granos de polen depositados en los estigmas de las flores, el cambio en el gremio de visitantes florales modificó el potencial reproductivo de los morfos a lo largo del periodo de floración de P. padifolia. Específicamente, el morfo Pin recibió una mayor cantidad de granos de polen ilegítimos (intramorfo) en el estigma de sus flores al inicio del periodo de floración. Lo anterior basado en la observación de la visita temprana de los lepidópteros a las flores y a su comportamiento de forrajeo dentro y entre plantas del morfo Pin. Por ello, los lepidópteros se consideraron como visitantes florales poco eficientes al momento de transferir polen legítimo entre morfos. En contraste, el morfo Thrum recibió una mayor proporción de granos de polen legítimo (intermorfo) en el estigma de sus flores como consecuencia de que los himenópteros se movieron entre morfos diferentes. Aunado a lo anterior, el ajuste entre el cuerpo de los himenópteros y la morfología floral de P. padifolia pudo favorecer la dirección en la transferencia de polen de Pin hacia Thrum (apertura de la corola y separación estigma-antera; Hernández y Ornelas, 2007a). Un caso similar fue reportado por Stone (1996) para Psychotria suerrensis. En ese estudio, Stone (1996) reportó que las abejas y mariposas transfirieron cantidades similares de polen a ambos morfos. No obstante, las abejas fueron capaces de transferir una mayor proporción de granos de polen legítimos (intermorfo) a las flores del morfo Pin como consecuencia de un mejor ajuste entre el cuerpo del polinizador y la morfología floral de Pin en P. suerrensis (Stone, 1996).
Durante el pico de floración de P. padifolia se observó la mayor carga de granos de polen en el estigma de las flores de Pin, pero Thrum presentó una mayor proporción de granos de polen legítimos (intermorfo) en el estigma de sus flores. La carga polínica estigmática en Pin fue claramente atribuida al movimiento de forrajeo entre plantas diferentes pero del mismo morfo por parte de los lepidópteros, mientras que el comportamiento de forrajeo de los troquilinos explicó la proporción de granos de polen legítimos en Thrum. Específicamente, los colibríes iniciaron su actividad de forrajeo visitando las flores de Pin y subsecuentemente visitaron a las plantas de Thrum, lo que promovió el flujo de polen de Pin hacia Thrum. El papel de los himenópteros como agentes que promueven cargas de polen ilegítimos (intramorfo) en el estigma de las flores de Pin ha sido previamente descrito para Psychotria suerrensis (Stone, 1996) y Psychotria nervosa (Hernández-Ramírez, 2012). No obstante, en este estudio se muestra el papel de los lepidópteros como agentes que promueven cargas de polen ilegítimo en Pin, mientras que los himenópteros promovieron un flujo de polen legítimo en el estigma de las flores de Thrum al inicio del periodo de floración en ausencia de colibríes.
Los colibríes pertenecientes al género Amazilia se han considerado como territoriales debido a la defensa activa y al monopolio de áreas específicas de alimentación de grupos de plantas de P. padifolia en Costa Rica y México (Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004; Ree, 1997). Las aves al establecer su territorio incrementan la probabilidad de mover el polen al interior de la planta, debido a la visita consecutiva de las flores en su parche de alimentación (geitonogamia; Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004; Ree, 1997). Aunado a lo anterior, el establecimiento de territorios puede propiciar cambios en la transferencia de polen de Pin hacia Thrum o viceversa, lo anterior en respuesta al sitio en el cual el ave territorial se percha (Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004). Contrario a lo reportado previamente, Amazilia cyanocephala no presentó un comportamiento territorial en el área y año de estudio, visitó ambos morfos indistintamente.
Al final del periodo de floración, Atthis heloisa y Eugenes fulgens visitaron las plantas Pin y Thrum en una proporción similar. Ambos colibríes presentaron un comportamiento de forrajeo siguiendo una ruta y secuencia de visita, lo cual asociado a una distribución espacial no agregada de los morfos en la población (Ornelas, González et al., 2004), contribuyó a explicar las cargas similares de polen legítimo al final del periodo de floración de la especie.
Estudios previos han enfatizado en el papel que lo colibríes tienen como polinizadores de P. padifolia (Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004; Hernández y Ornelas, 2007a). Lo anterior se ha basado en la frecuencia de visita observada de los troquilinos durante el pico de la floración de la especie, así como de la carga de polen legítimo sobre los estigmas de Thrum (Ornelas, Jiménez et al., 2004; Hernández y Ornelas, 2007a). En este estudio se evidenció el papel dominante que tienen los invertebrados en términos de frecuencia de visita cuando se consideró el ciclo anual reproductivo de P. padifolila. Los himenópteros jugaron un papel importante en la transferencia de polen legítimo entre morfos y al igual que los troquilinos, depositaron una mayor proporción de polen legítimo en los estigmas de Thrum en ausencia de colibríes. Por ello, himenópteros y troquilinos parecen jugar un papel determinante en el éxito reproductivo de la especie, el cual requiere ser estudiado a mayor detalle.
Se ha sugerido que la falla en el flujo de polen entre morfos diferentes (polinización legítima, intermorfo) propiciaría el rompimiento del sistema distílico (Darwin, 1877; Ganders, 1979). Para el caso de P. padifolia, se ha propuesto como ruta de evolución la especialización del género (Pin como donador de polen o función masculina, Thrum como receptor de polen o función femenina; Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004). Lo anterior se debe a que la especie exhibe una imprecisión en la reciprocidad estigma-antera entre morfos y una asimetría en la transferencia de polen legítimo entre morfos (Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004). Los resultados obtenidos en este estudio muestran que la presencia de polen legítimo en el estigma de las flores en ambos morfos es variable y suficiente para fertilizar los óvulos de las flores durante el ciclo anual reproductivo de la especie (solo se requieren 2 granos de polen legítimos; Hernández y Ornelas, 2007a). La transferencia de polen legítimo en el estigma de las flores llega a ser similar para ambos morfos al final del periodo de floración, lo que permitiría mantener el sistema distílico. Es decir, la condición variable de carga de polen estigmática observada durante el ciclo anual de floración de P. padifolia permite mantener la imprecisión en el polimorfismo estigma-antera previamente observado en la especie a nivel poblacional (Hernández y Ornelas, 2007a; Ornelas, González et al., 2004; Ornelas, Jiménez et al., 2004). Price y Barrett (1984) discutieron que la condición variable en el flujo de polen observado entre morfos durante el ciclo anual de floración de Pontederia cordata permitiría el mantenimiento de su sistema heterostílico. Lo anterior, considerando que el flujo asimétrico de polen entre morfos se traduce a nivel poblacional y no individual (Price y Barrett, 1984).
Los resultados obtenidos en este estudio enfatizan en la importancia de reconocer los atributos de Pin y Thrum desde una perspectiva poblacional (complementaria), independientemente de las trayectorias divergentes que potencialmente los morfos pudieran experimentar (especialización del género a través de procesos de asignación sexual; Hernández y Ornelas, 2007b). La búsqueda de un “polinizador eficiente” como agente de selección que moldeé el ajuste preciso en los rasgos florales entre morfos en especies heterostílicas que por lo general están expuestas a cambios en la identidad y comportamiento de sus visitantes florales requiere ser re-evaluado a nivel conceptual y metodológico.

