











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
En México los bosques de Pinus son frecuentes en zonas de montaña con clima templado y frío. Se encuentran constituidos como elementos dominantes o comparten la dominancia con especies de otros géneros como Quercus, Abies u otras latifoliadas. Cubren amplios rangos elevacionales, desde el nivel del mar como Pinus caribaea var. hondurensis, hasta las regiones de alta montaña con Pinus hartwegii a 4,200 m snm (Perry et al., 2000). Gernandt y Pérez-de la Rosa (2014) registran para México 49 de las 120 especies de pino que existen en el mundo, por lo cual se considera a este país como un centro secundario de diversificación. Algunas especies de Pinus llegan a formar comunidades adaptadas a crecer en condiciones que generalmente son desfavorables para otras especies; además se sabe que son resistentes a las heladas, periodos de sequías, incendios frecuentes y al pastoreo (Rzedowski, 2006). Algunas especies son consideradas como pioneras porque se establecen en las primeras etapas de la sucesión (Cayuela et al., 2006; Galindo-Jaimes et al., 2002). Sin embargo, en los procesos sucesionales más avanzados sucumben ante la intolerancia de condiciones de sombra, llegando a ser sustituidos por otras especies de mayor tolerancia, como la mayoría de las especies del género Quercus, así como especies del bosque mesófilo de montaña y algunas especies de Abies (Lara-González et al., 2009; Weber et al., 2008). Por su parte, Abies se distribuye de forma insular en zonas elevadas del Eje Neovolcánico Transversal, así como en las porciones más elevadas de la Sierra Madre del Sur (Rzedowski, 2006) y son considerados relictos de la extensión de los bosques boreales que avanzaron de norte a sur durante los periodos glaciares (Rzedowski y McVaugh, 1966), cuando los climas fríos alcanzaron latitudes tropicales. Se conocen 8 especies y 5 variedades en México (Gernandt y Pérez-de la Rosa, 2014). Su distribución está confinada a laderas protegidas del viento y de la incidencia solar elevada, y su gradiente altitudinal es desde los 2,000 a 3,600 m (Lara-González et al., 2009). Los bosques puros de Abies son considerados poco complejos estructuralmente con baja diversidad y riqueza de especies (Gernandt y Pérez-de la Rosa, 2014). Sin embargo, al occidente de México se ha encontrado que las fitoasociaciones que forman con el bosque mesófilo de montaña y las bajas altitudes a las que descienden los Abies presentan mayor diversidad y riqueza de especies que lo reportado en otras áreas de México (Cuevas-Guzmán et al., 2011; Guerrero-Hernández et al., 2014) por lo que esta asociación vegetal de Pinus-Abies resalta la importancia de las condiciones ambientales pasadas y sus adaptaciones a las condiciones actuales. Diversos estudios se han enfocado en la ecología, biogeografía y diversificación del bosque de Pinus y del bosque de Abies (Encina-Domínguez et al., 2008; Manzanilla, 1974; Richardson y Rundel, 1998). Sin embargo, son escasos los estudios encaminados a entender la contribución del cambio climático y las actividades humanas en su expansión y persistencia (Figueroa-Rangel et al., 2008).
Estudios paleoecológicos (e.g., Lozano-García et al., 2007; Ortega-Rosas et al., 2008) señalan que, durante el Holoceno temprano, en respuesta al calentamiento que caracterizó este periodo, se presentó un aumento en la expansión de los bosques de Pinus en diversas regiones de México. Sin embargo, durante el Holoceno tardío se presentaron condiciones variables en el clima que condujeron a 2 principales anomalías: la Anomalía Climática Medieval (ACM) (950-1400 d.C.), que fue un periodo más cálido al promedio normal y la Pequeña Edad de Hielo (PEH) (1400-1900 d.C.), un periodo frío en general pero de mayor variabilidad sobre todo en latitudes medias y altas del Hemisferio Norte (Díaz et al., 2011). Los registros de diversos sitios en México señalan que la respuesta ante la variabilidad climática es compleja y la duración en cada lugar puede ser diferente (Caballero et al., 2006; Lozano-García et al., 2007; Metcalfe et al., 2010; Rodríguez-Ramírez et al., 2015; Sosa-Nájera et al., 2010). Estos cambios ambientales se atribuyen a la variación en el balance de diferentes forzamientos globales, tal como la actividad solar (Bond et al., 2001), así como con los cambios en la posición media latitudinal de la zona intertropical de convergencia (ZITC) y El Niño-Oscilación del Sur (ENOS) (Metcalfe et al., 2000). También se ha documentado que algunos de los bosques de Pinus se han generado como consecuencia de impacto humano, derivados de las actividades del desmonte de tierras para la explotación forestal y la agricultura (Almeida-Leñero et al., 2005; Ludlow-Wiechers et al., 2005). Estudios arqueológicos aledaños al sitio de estudio en la Sierra de Cacoma del estado de Jalisco, indican que han sido ocupados desde ~1200 d.C., por poblaciones indígenas (Jardel, 1994; Jiménez, 1995; Kelly, 1945; Weigand, 1996). La evidencia arqueológica sugiere que la actividad humana ha estado presente en las partes bajas del sitio de estudio, y archivos históricos indican la existencia de diferentes localidades indígenas antes de la llegada de los españoles (Acuña, 1988). Excavaciones arqueológicas en Autlán, una localidad cercana ubicada al suroeste del sitio de la sierra de Cacoma, señalan la presencia de asentamientos humanos por lo menos desde 3,400 ± 200 años antes del presente (Kelly, 1980). Sin embargo, la cerámica prehispánica encontrada en dicha zona ubica la presencia de asentamientos humanos entre 800 y 500 años antes del presente (Kelly, 1945). Algunos documentos históricos del siglo XVI de la Nueva Galicia mencionan la llegada de los españoles en la región en 1524 d.C. y registran 22 asentamientos humanos en 1579 d.C. en tierras bajas alrededor del sitio de estudio (Acuña, 1988). Las poblaciones indígenas estaban comprendidas principalmente por grupos nahuas y otomíes asentadas en aldeas dispersas; se dedicaban a cultivar la tierra bajo la modalidad de roza, tumba y quema, producían maíz, frijol y calabaza, principalmente (Tetreault et al., 2011). Posterior a la llegada de los españoles se modificaron las actividades culturales de los indígenas nativos, se intensifica la explotación forestal, se introduce ganado bovino y se abren áreas por el aprovechamiento minero, la cual se convirtió en una actividad importante en la región durante esa época, por lo que las poblaciones humanas se establecieron cercanas a las minas (Jiménez, 1995).
Los bosques de Pinus-Quercus-Abies de alta montaña en el occidente de México, representan una fuente importante de recursos maderables y de servicios ambientales. Sin embargo, la sierra de Cacoma es una región que ha sido poco explorada y el conocimiento sobre la dinámica de la vegetación y la influencia de las actividades humanas a largo plazo es desconocido. Por lo tanto, es de interés conocer la historia en la distribución del bosque de Pinus-Quercus-Abies del sitio de estudio, la incidencia de las actividades humanas en el área, además de entender si la distribución actual del bosque de Pinus-Quercus-Abies es consecuencia de los procesos naturales como el cambio climático y los incendios forestales, e inclusive del incremento de los periodos de aridez del último milenio. Por lo que, el objetivo del trabajo es responder las preguntas: ¿Cuál ha sido la dinámica del bosque de Pinus-Quercus-Abies durante los últimos 1,580 años? ¿Cuál ha sido la influencia del cambio climático en su dinámica? ¿Es posible explicar la permanencia y expansión del bosque de Pinus-Quercus-Abies a partir de actividades antropogénicas durante los últimos 1,580 años?
Materiales y métodos
El sitio de estudio se localiza en la sierra de Cacoma al occidente de Jalisco, México en el paraje denominado La Cumbre de Guadalupe, una región de confluencia entre la cordillera del Eje Neovolcánico Transversal y la sierra Madre del Sur (Fig. 1). Corresponde a una elevación de 2,119 m , y se extiende hasta el litoral del Pacífico y la costa del occidente de México (Inegi, 2015). El área de estudio es una zona de gran complejidad geomorfológica y litológica que presenta afloramientos rocosos formados a partir de procesos tectónicos que dieron lugar al levantamiento de un batolito (rocas ígneas intrusivas) y pisos marinos (rocas calizas) durante el Cretácico (Conabio, 2010). El tipo de suelo es regosol dístrico, cambisol dístrico y litosol (Inegi, 1976). Presenta un clima templado subhúmedo, con una temperatura media anual de 14.2 °C, precipitación anual de 2,003 mm con lluvias en verano (estación 14,271 de la localidad La Cumbre de Guadalupe, altura 2,120 m snm, 20°10’20” N, 104°42’45” O, Servicio Meteorológico Nacional, http://smn.cna.gob.mx/). La temperatura media del mes más cálido (mayo y junio) es de 16.7 °C. La precipitación está concentrada de mayo a septiembre y durante los meses de junio a enero y la incidencia de neblina es frecuente (Fig. 2). En la actualidad corresponde a un tipo de vegetación dominado por diversas especies de Pinus y Quercus, así como individuos de Abies jaliscana. Los elementos florísticos comprenden a Alnus acuminata, A. jorullensis, Arbutus xalapensis, Carpinus tropicalis, Clethra fragrans, Pinus oocarpa, P. jaliscana, P. herrerae, P. pseudostrobus, Quercus castanea, Q. candicans, Q. scytophylla, Q. magnoliifolia, Q. obtusata y Ternstroemia lineata. El componente herbáceo es diverso, representado por Amaranthus palmeri, Chenopodium ambrosoides, Iresine diffusa, Eryngium alternatum, Archibaccharis serratifolia, Baccharis heterophylla, Bidens odorata, B. aurea, Dahlia coccinea, Ageratina choricephala, Melampodium perfoliatum, Piqueria triflora, Podachaenium eminens, Tagetes filifolia, T. lucida, Crotalaria filifolia, C. mollicula, Dalea obreniformis, Mimosa albida, Fuchsia fulgens, Fuchsia microphylla, Salvia iodantha, entre otras.


Figura 2 Precipitación y temperatura medias mensuales y anuales para la zona de estudio, (estación 14,271 Servicio Meteorológico Nacional, http://smn.cna.gob.mx/).
Procesamiento de muestras e indicadores paleoambientales. Se extrajo un núcleo de sedimento de 62 cm de profundidad con un taladro Eijelkamp en una hondonada forestal. De acuerdo con Calcote (1995, 1998), consiste en una depresión natural sobre la que se deposita el polen a lo largo del tiempo por procesos naturales de sedimentación. La extracción del núcleo de sedimento se efectuó en un claro de bosque dominado en la actualidad por diversas especies del género Pinus y Quercus y una del género Abies (del Castillo-Batista, 2017) en el paraje denominado La Cumbre de Guadalupe (Fig. 1), (20°10’ 17.82” N, 104°42’42.32” O, a una elevación de 2,119 m). El núcleo se cubrió por completo con papel film transparente y posteriormente con papel aluminio sobre el cual se anotó la información correspondiente. El núcleo de sedimento se trasladó en tubos de PVC y se refrigeró a 5 °C para conservar las condiciones anaeróbicas. En el laboratorio de Paleoecología y Cambio climático de la Universidad de Guadalajara, se realizó el análisis de susceptibilidad magnética y posteriormente se tomaron muestras con resolución de un centímetro a lo largo del núcleo de sedimento de donde se extrajo polen fósil, micropartículas de carbón y materia orgánica. Además, utilizando técnicas palinológicas estandarizadas (Faegri e Iversen, 1989) se extrajeron palinomorfos de 62 muestras utilizando un volumen de 0.5 cm3. Cada muestra se montó en gelatina glicerina y se contaron 400 granos de polen y esporas para obtener la suma palinológica. El conteo de palinomorfos se realizó en un microscopio óptico Primo Star, Carl Zeiss con una magnificación de 400 aumentos. La determinación de polen y esporas se realizó utilizando claves palinológicas y bibliografía especializada; así como la opinión de expertos en palinología, completando la consulta con la colección de referencia palinológica del Laboratorio de Paleoecología y Cambio Climático de la Universidad de Guadalajara. Se añadieron a cada muestra 2 tabletas de Lycopodium clavatum L., con el fin de estimar la concentración de polen (granos de polen cm-3) y la concentración de partículas de microcarbón fósil (microcarbón cm2cm-3) (Stockmarr, 1971). La suma de polen se utilizó para calcular la abundancia relativa de las especies expresada en porcentajes incluyendo árboles-arbustos, herbáceas y pteridofitas sensu lato, con lo cual se construyó el diagrama polínico utilizando el programa PSIMPOLL 4.25 (Bennett, 2005). Las partículas de microcarbón fósil fueron contadas siguiendo la metodología propuesta por Clark (1982) mediante lo cual se estimó la ocurrencia de incendios forestales pasados y su relación con los cambios en la composición taxonómica y ambiental del sitio (Whitlock y Larsen, 2001). Esta técnica consiste en el conteo del número de micropartículas de carbón proyectadas sobre puntos determinados en la laminilla palinológica. Los valores de concentración de éstas se presentan como el número de partículas de microcarbón cm-3.
Como indicador de cambio paleoambiental se utilizó la susceptibilidad magnética, ésta mide el aporte de minerales magnéticos en el sedimento (Thompson y Oldfield, 1986) y se considera como un indicador de procesos de erosión. El análisis de susceptibilidad magnética se realizó cada centímetro a lo largo del núcleo de sedimento de 62 cm utilizando un dispositivo Bartington MS2 (Bartington Instruments, Witney Oxford, Reino Unido). Se determinó la pérdida progresiva por ignición (LOI) con el fin de estimar el contenido de materia orgánica y carbonatos del núcleo de sedimento utilizando las relaciones lineales entre los valores LOI y el contenido de materia orgánica y los carbonatos expresados en porcentaje (Dean, 1974). Este análisis se realizó a cada centímetro en todo el núcleo de sedimento, tomando muestras de 0.5 cm3 y mediante la pérdida de peso en las muestras al ser sometidos a altas temperaturas secuenciales (550 °C y 910 °C). La interpretación en los cambios de porcentaje de LOI está relacionada con eventos de mayor productividad y estabilización de la vegetación. El análisis de la composición de elementos químicos (e.g., As, Ca, Fe, Mo, Pb, Rb, Sr, Ti, Zn y Zr) se realizó por medio de fluorescencia de rayos X (FRX) a cada 3 centímetros en todo el núcleo de sedimento con un espectrómetro manual Thermo Scientific Niton XL3t. Los resultados del análisis de elementos se expresan en la concentración de partes por millón (ppm) los cuales se utilizan como indicadores de cambios ambientales. En particular el titanio es considerado indicador del registro de aporte de humedad (registro clástico), mientras que el Ca puede indicar momentos de menor sedimentación, por lo que valores altos de Ti y bajos de Ca indican un incremento en el aporte de sedimentos terrígenos
El modelo cronológico de edad-profundidad fue calculado por interpolación lineal basado en 4 dataciones radiométricas (14C; Beta Analytic de Florida, USA) a lo largo del núcleo de sedimento mediante la técnica de espectrometría de aceleración de masas (AMS, por sus siglas en inglés). Las fechas de radiocarbono fueron calibradas con el programa CALIB v5.02 (Stuiver y Reimer, 1993) e INTCAL09.14 (Reimer et al., 2009). Las zonas palinológicas se determinaron por el método de conglomerados CONISS (Constrained Cluster Analysis by Sum of Squares) (Grimm, 1987). Se aplicó un análisis de rarefacción [E(Tn)] para estimar la riqueza palinológica en cada muestra, esta técnica permite estimar la riqueza polínica que se esperaría obtener si los conteos polínicos tuvieran el mismo tamaño de muestra (Birks et al., 2012). Así mismo, se realizó el análisis de la tasa de cambio para identificar los cambios sincrónicos de la vegetación entre las muestras de polen, este análisis mide la disimilitud entre pares de muestras adyacentes y la diferencia temporal entre las muestras (Bennett, 2005). Todos los análisis se efectuaron con el programa PSIMPOLL 4.25 (Bennett, 2005). Se realizó un análisis de correlación cruzada entre 6 taxones leñosos (Pinus, Alnus, Parathesis, Quercus, Ternstroemia y Cupressaceae) y las partículas de microcarbón fósil, así como una correlación cruzada entre las partículas de microcarbón fósil y 2 taxones herbáceos (Cyperaceae y Asteraceae) con el programa estadístico R (R Core Team, 2013) para estimar el grado de correlación entre 2 muestras (taxón/microcarbón fósil) y determinar su sincronía.
Se realizó un análisis de redundancia (RDA) con el fin de identificar las variables ambientales que mejor explican los patrones de ordenación observados en la composición de los taxones (Ter Braak, 1986). El RDA es un método directo de ordenación apropiado cuando se asume una relación monotónica entre los datos de ambas matrices (Ter Braak, 1986). Las variables ambientales incluidas en el análisis RDA fueron la susceptibilidad magnética, el contenido de materia orgánica, carbonatos y las partículas de microcarbón fósil. El análisis de ordenación se realizó con el programa CANOCO 5.0 (Ter Braak y Smilauer, 2002).
Resultados
La datación radiocarbono en la base de la secuencia sedimentaria del bosque de Pinus-Quercus-Abies de La Cumbre de Guadalupe (CUM-N2) indica que la acumulación del sedimento comenzó aproximadamente hace 1,580 años sin hiato aparente. Sin embargo, la tasa de acumulación de sedimento no fue constante. El modelo de edad-profundidad (Fig. 3) se obtuvo mediante interpolación lineal a partir de las dataciones obtenidas en 4 fechados 14C (Tabla 1), lo que permitió estimar la tasa de sedimentación promedio de 25 años/cm.

Figura 3 Modelo de edad-profundidad del núcleo de sedimento CUM-N2. La edad calibrada se presenta en años calendáricos (d.C.).
Tabla 1 Fechas de radiocarbono del núcleo de sedimento CUM-N2 calibradas en edades antes del presente (AP) y en años calendáricos (d.C.).
| Código de laboratorio | Profundidad (cm) | Edad 14 C (años AP) | Edad calibrada años d.C. 2σ |
|---|---|---|---|
| Beta - 343626 | 6 | 101.3±30 pMC | 57-73 |
| Beta - 343627 | 16 | 740±30 AP | 1220-1280 d.C. |
| Beta - 343628 | 36 | 1280±30 AP | 660-730 d.C. |
| Beta - 339533 | 62 | 1580±30 AP | 390-540 d.C. |
El registro palinológico del bosque de Pinus-Quercus-Abies reveló 67 taxones con 3 zonas palinológicas de acuerdo al análisis de conglomerados (Fig. 4). Pinus se presentó en toda la secuencia estratigráfica dominando el estrato arbóreo, principalmente durante la ACM y la PEH, con fluctuaciones constantes en su porcentaje a lo largo de toda la secuencia. En el estrato herbáceo las familias Asteraceae, Cyperaceae, Poaceae, Polypodiaceae, Aspleniaceae y Adiantaceae fueron los taxones más dominantes y al igual que Pinus estas familias estuvieron presentes en toda la secuencia estratigráfica. En el grupo de pteridofitas sensu lato Hymenophyllaceae fue la familia más dominante. Algunos taxones de bosque mesófilo de montaña (Ostrya-Carpinus, Parathesis y Fraxinus) también estuvieron presentes, aunque poco representados en toda la secuencia estratigráfica. La zonación de los datos palinológicos se describe a continuación expresada en años de la era común.

Figura 4 Diagrama polínico del núcleo de sedimento CUM-N2 representando ~1,580 años de tasas de acumulación de especies del bosque de Pinus-Quercus-Abies. Se muestra la división de 3 zonas palinológicas mediante CONISS: zona uno (C-1); zona 2 (C-2); zona 3 (la cual se divide en C-3a y C-3b). La franja horizontal gris oscura representa la Anomalía Climática Medieval (ACM) y la franja en gris claro representa la Pequeña Edad de Hielo (PEH).
Zona C-1 (62-59 cm, 370 d.C. - 405 d.C.). Los cambios más evidentes para esta zona se establecen con la dominancia de taxones arbóreos como Alnus, Pinus y Quercus con valores entre 10-25% (Fig. 4); otros taxones arbóreos presentes en esta zona, aunque con bajos porcentajes son Abies, Fraxinus, Ostrya-Carpinus, Parathesis y Xylosma, así como las epífitas representadas por las familias Hymenophyllaceae y Bromeliaceae, además de Sphagnum, un tipo de musgo de hábitats húmedos como las turberas. Los taxones herbáceos son poco abundantes, con porcentajes menores al 10%, representado principalmente por las familias Poaceae, Cyperaceae, Asteraceae, Lamiaceae, Aspleniaceae, Polypodiaceae y Adiantaceae (Fig. 4). Otros taxones herbáceos con porcentajes menores al 5% son Chenopodiaceae, Apiaceae, Onagraceae y Plantago. Por su parte, la riqueza palinológica registra valores altos para esta zona con el mayor pico a los 393 d.C., al igual que la tasa de cambio (Fig. 5).

Figura 5 Síntesis de datos palinológicos con la suma de todos los taxones: árboles-arbustos en color negro, herbáceas en color gris oscuro y pteridofitas en color gris claro. Además, en esta misma figura se presenta la concentración total de polen, la concentración de micropartículas de carbón, la tasa de cambio y la riqueza palinológica del núcleo CUM-N2. La franja horizontal gris oscura representa la Anomalía Climática Medieval (ACM) y la franja gris claro representa la Pequeña Edad de Hielo (PEH).
Zona C-2 (59-17 cm, 405-1183 d.C.). Al inicio de esta zona Pinus presenta porcentajes bajos (10%), con un aumento considerable a los 1048 d.C. y valores de hasta 35%. Por el contrario, Alnus se presenta como el taxón más dominante del estrato arbóreo al inicio de esta zona, pero rápidamente disminuye con menos del 10% alrededor de 520 d.C. Quercus se observa en toda la secuencia con valores fluctuantes menores al 10%, presentándose como codominante con Pinus y Alnus (Fig. 4). Otros taxones arbóreos presentes en esta zona con porcentajes bajos son Abies, Cyathea, Fraxinus, Ostrya-Carpinus, Parathesis, Salix, Ternstroemia y Xylosma. Las familias presentes en esta zona son Cupressaceae, Lauraceae, Leguminosae, Myrtaceae y Rosaceae. En el estrato herbáceo las familias más abundantes son Asteraceae, Cyperaceae y Poaceae con valores de 10 a 25%. Zea y Phaseolus aparecen por primera vez en el registro estratigráfico en esta zona (Fig. 4). Otros taxones culturales como Amaranthaceae están presentes con valores bajos. Para el grupo de las pteridofitas y herbáceas terrestres, la familia Hymenophyllaceae es la más abundante, seguida de Polypodiaceae. Otras familias como Adiantaceae, Aspleniaceae y Bromeliaceae están presentes en esta zona con porcentajes bajos. Tres eventos en el recambio de taxones tienen lugar en esta zona, mientras que la riqueza palinológica presenta valores fluctuantes a lo largo de esta zona (Fig. 5).
Zona C-3 (17-0 cm, 1183 d.C.-Presente). Esta zona está dividida en 2 subzonas, la primera de ellas (C-3a) ocurre entre ~1183-1280 d.C., representa la transición entre la ACM y la PEH los taxones leñosos y las epífitas presentan porcentajes más elevados, 45% y 30% respectivamente, en comparación con la vegetación herbácea con 20%. Pinus tiene el mayor porcentaje de los taxones leñosos con el 25%, seguido de Alnus con el 15%, después se encuentra Ternstroemia, Xylosma y Quercus con el 5%, mientras que Hymenophyllaceae presenta 15% seguida de Sphagnum con 10% y Polypodiaceae con 5% (Fig. 4). Con respecto a los taxones herbáceos en esta zona, el polen de la familia Asteraceae se mantiene abundante con el 15% en comparación con Cyperaceae y Poaceae que presentan menos del 5%. Por su parte, el registro palinológico de la zona C-3b muestra los porcentajes de Pinus como los más altos con fluctuaciones constantes a lo largo de la secuencia; Alnus y Quercus se mantienen en el estrato arbóreo con porcentajes variables. Otros taxones con bajos porcentajes (menores de 5%) son Abies, Fraxinus, Parathesis, Salix y Xylosma. La comunidad de herbáceas está representada principalmente por Asteraceae, Cyperaceae y Poaceae, aunque con porcentajes irregulares, Zea y Phaseolus continúan su registro en esta zona hasta el presente (Fig. 4). En el conjunto de las pteridofitas, las esporas de Hymenophyllaceae son abundantes al inicio de la zona, decreciendo sus porcentajes al final de la secuencia, los helechos como Polypodiaceae, Adiantaceae y Aspleniaceae presentan porcentajes muy bajos. En cambio, Sphagnum presenta un ligero aumento al inicio de esta zona y disminuye al final de la misma. Esta zona muestra un incremento de la riqueza palinológica a los ~1350 d.C., registrando una disminución a los ~1491 d.C. con los valores más bajos de toda la secuencia, manteniendo fluctuaciones hasta el presente. La tasa de cambio manifiesta periodos de recambio de taxones a los ~1210 d.C. y, a partir de ~1912 d.C., se registra un incremento considerable hasta el presente (Fig. 5).
El registro de partículas de microcarbón fósil indica la ocurrencia de incendios durante los ca. 1,580 años que comprende la secuencia sedimentaria (Fig. 5). Sin embargo, los incendios más intensos se registran en los periodos de ~647, 967, 1048, 1210 y 1491 d.C., a mediados de la zona C-2 y en la zona C-3a y C-3b. Los resultados de la susceptibilidad magnética indican valores altos en la base de la secuencia, pero se caracteriza por presentar grandes variaciones dentro de la misma, al igual que los valores de la materia orgánica, que registra valores bajos en la base de la secuencia en la zona C-1, con aumentos irregulares hasta el final de la secuencia en la zona C-3b (Fig. 6). La mayor concentración de materia orgánica se registra a los ~1491 d.C. en la zona C-3. El contenido de carbonatos es irregular a lo largo de la secuencia con valores altos en la base de la zona C-1 y una disminución al inicio de la zona C-2. Sin embargo, se observan 3 picos de mayor concentración: el primero a los 393 d.C., el segundo a los 1280 d.C. y el tercero que corresponde a la zona C-3b. Con relación a la composición química (Fig. 7), observamos que As, Zn y Fe muestran valores bajos al inicio de la secuencia, posteriormente presentan valores irregulares y aumentan gradualmente hasta el presente, mientras que Pb, Sr y Ca muestran valores altos al inicio de la secuencia y disminuyen en el presente; finalmente el Ti presenta valores menos irregulares a lo largo de la secuencia (Fig. 7).
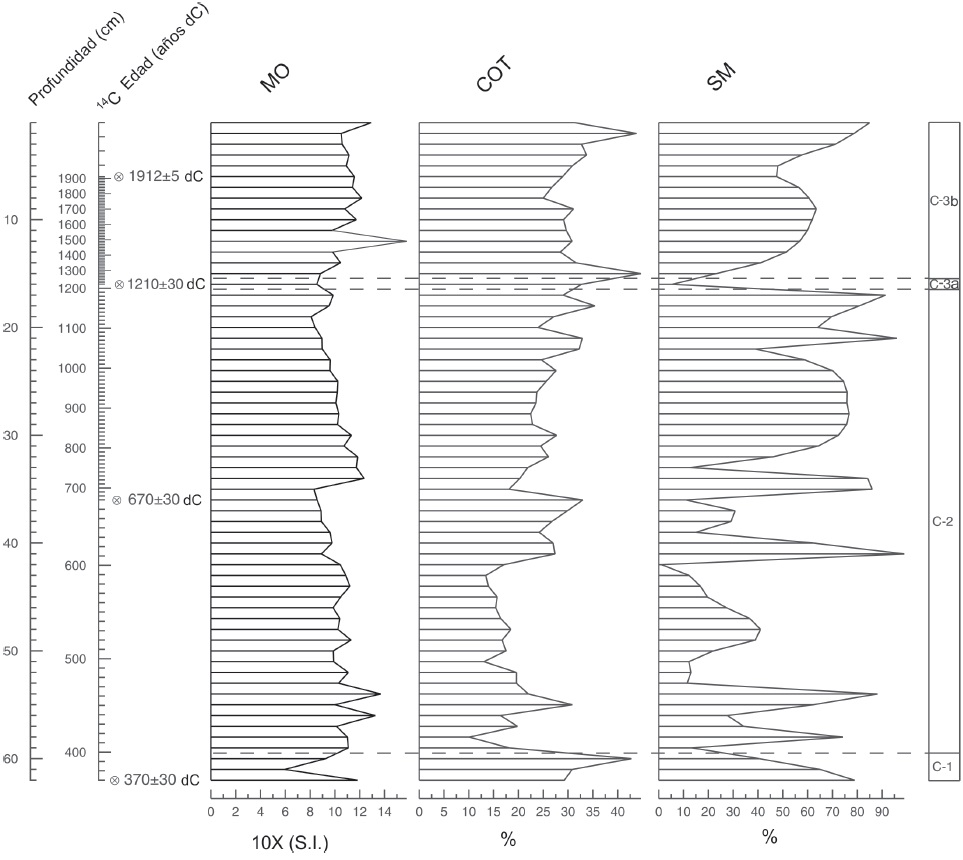
Figura 6 Materia orgánica (MO), carbonatos (COT) y susceptibilidad magnética (SM) del núcleo CUM-N2. Las zonas palinológicas están delimitadas de acuerdo a la opción de división óptima (CONISS) disponible en PSIMPOL 4.25.
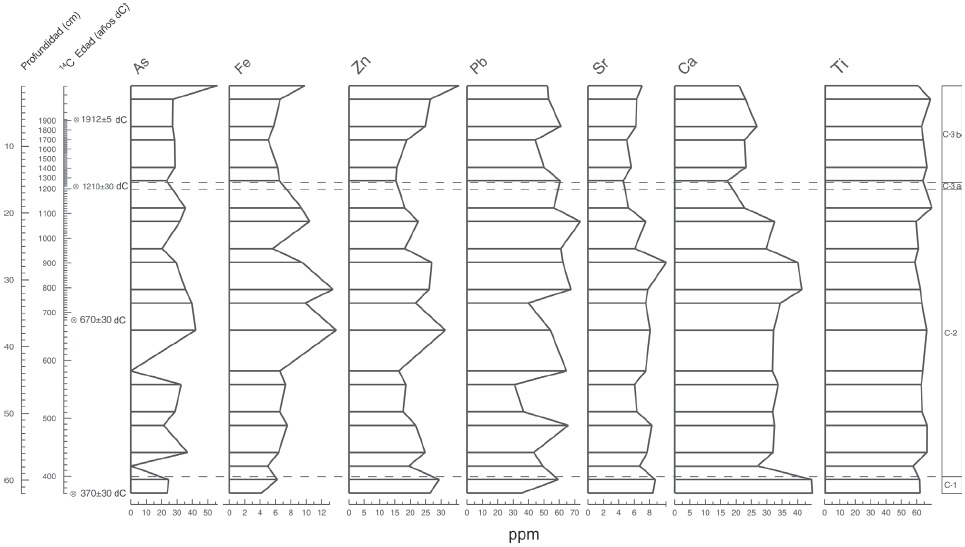
Figura 7 Análisis de elementos químicos del núcleo CUM-N2; se muestran los principales elementos registrados en orden de izquierda a derecha: arsénico (As), hierro (Fe), zinc (Zn), plomo (Pb), estroncio (Sr), calcio (Ca) y titanio (Ti).
El RDA extrajo el 69.64% del total de la varianza en los 2 primeros ejes; el primero explicó el 43.39% de la varianza; las variables con mayor correlación con este eje fueron las partículas de microcarbón fósil (MC), los carbonatos (COT) y la materia orgánica (MO) (Tabla 2); los principales taxones asociados a estos gradientes ambientales fueron Pinus, Adiantaceae e Hymenophyllaceae (Fig. 8). En el eje 1 los taxones estrechamente asociados con MC fueron Cyathea (Cyat), Salix (Sali), Ternstroemia (Tern) y Polypodiaceae (Poly). En el eje 2, las variables ambientales que resultaron significativas fueron COT y SM, mientras que los taxones estrechamente relacionados con COT y SM fueron Phaseolus (Phas), Celastraceae (Cela), Juncaceae (Junc), Fraxinus (Frax), Arbutus (Arbu), Onagraceae (Onag) y Zea (Fig. 8).
Tabla 2 Análisis de Redundancia (RDA) para las profundidades de núcleo de sedimento CUM-N2. Coeficientes de correlación (CC) de variables ambientales y valores de t (t); los números en negritas representan el valor crítico de (df ≥ α = 0.05) = 2.1.
| Eje 1 | Eje 2 | |||
|---|---|---|---|---|
| Eigenvalores | 0.1161 | 0.0665 | ||
| Variables ambientales | CC | t | CC | t |
| MC | 0.6671 | 5.1836 | -0.6772 | -0.8512 |
| MO | 0.3131 | 2.4172 | 0.1500 | 0.8473 |
| COT | 0.5367 | 3.9428 | 0.6710 | 3.6080 |
| SM | 0.0680 | 0.5227 | 0.3863 | 2.1744 |
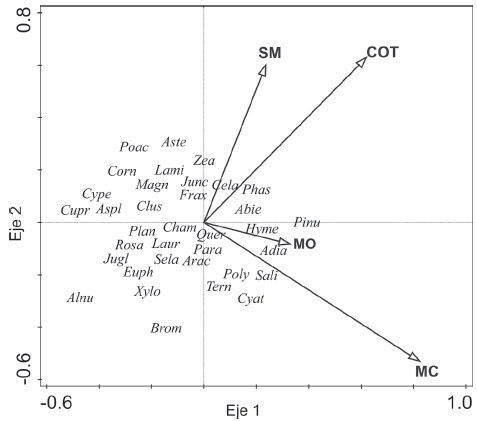
Figura 8 Análisis de redundancia (RDA) de los taxones de árboles-arbustos, herbáceas y epífitas en relación con las variables ambientales: susceptibilidad magnética (SM), carbonatos (COT), concentración de materia orgánica (MO) y las micropartículas de carbón (MC). Los taxones representados por su abreviación son los siguientes: Abies (Abie), Alnus (Alnu), Arbutus (Arbu), Adiantaceae (Adia), Apiaceae (Apia), Aspleniaceae (Aspl), Asteraceae (Aste), Bromeliaceae (Brom), Clethra (Clet), Clusia (Clus), Cornus (Conr), Cyathea (Cyat), Caryophyllaceae (Cary), Celastraceae (Cela), Cloranthaceae (Clor), Cupressaceae (Cupr), Cyperaceae (Cype), Euphorbiaceae (Euph), Fraxinus (Frax), Chenopodiaceae (Chen), Hedera (Hede), Equisetum (Equi), Hymenophyllaceae (Hyme), Ilex (Ilex), Juglans (Jugl), Juncaceae (Junc), Lamiaceae (Lami), Lauraceae (Laur), Magnolia (Mang), Malvaceae (Malv), Myrtaceae (Myrt), Ostrya -Carpinus (Osca), Onagraceae (Onag), Ophioglossaceae (Ophi), Parathesis (Para), Phaseolus (Phas), Pinus (Pinu), Plantago (Plan), Poaceae (Poac), Polypodiaceae (Poly), Quercus (Quer), Rosaceae (Rosa), Salix (Sali), Selaginella (Sela), Solanaceae (Sola), Ternstroemia (Tern), Tilia (Tili) y Zea (Zea).
La ordenación de las edades mediante RDA, agrupó en el eje uno a 439, 647, 967, 1048,1210 y 1491 d.C. cerca del vector MC, que corresponden a las edades con las mayores concentraciones de las partículas de microcarbón fósil en la secuencia. En el eje 2 se agruparon las edades de 393, 1156,1280, 1701, 1935, 1942 y 2012 d.C. cerca de los vectores COT y SM (Fig. 9).
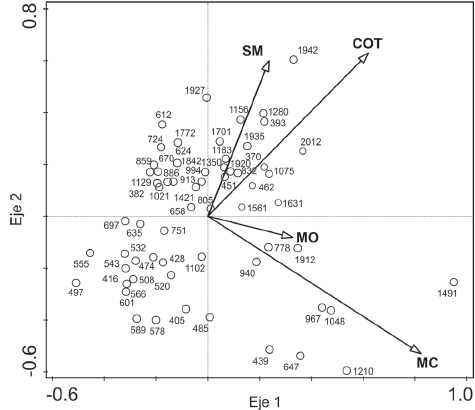
Figura 9 Análisis de redundancia (RDA) de las edades (años d.C.) con relación a las variables ambientales del núcleo CUM-N2. Las variables ambientales corresponden a susceptibilidad magnética (SM), carbonatos (COT), concentración de materia orgánica (MO) y las micropartículas de carbón (MC).
El análisis de correlación cruzada indicó que Cupressaceae, Cyperaceae y Asteraceae se correlacionan negativamente (p < 0.05) con la abundancia de las partículas de microcarbón fósil (lag 0), esto implica que sus abundancias son inversamente proporcionales, es decir cuando las partículas de microcarbón fósil aumentan, los taxones de Cupressaceae, Cyperaceae y Asteraceae disminuyen en su abundancia, más o menos en los mismos intervalos de tiempo. En contraste, se correlacionan positivamente con Pinus y Ternstroemia, ambos en sincronía con las partículas de microcarbón fósil (lag 0), correspondiendo a un incremento con las partículas de microcarbón fósil que coinciden con el incremento en Pinus y Ternstroemia en los mismos intervalos. Por el contrario, Alnus y Parathesis se correlacionan negativamente (p < 0.05) con las partículas de microcarbón fósil (lag 4) y con Quercus (lag 7), lo que significa que el aumento en la abundancia de Alnus y Parathesis ocurre 100 años después de un pico de las partículas de microcarbón fósil y 200 años para Quercus, respectivamente (Fig. 10).
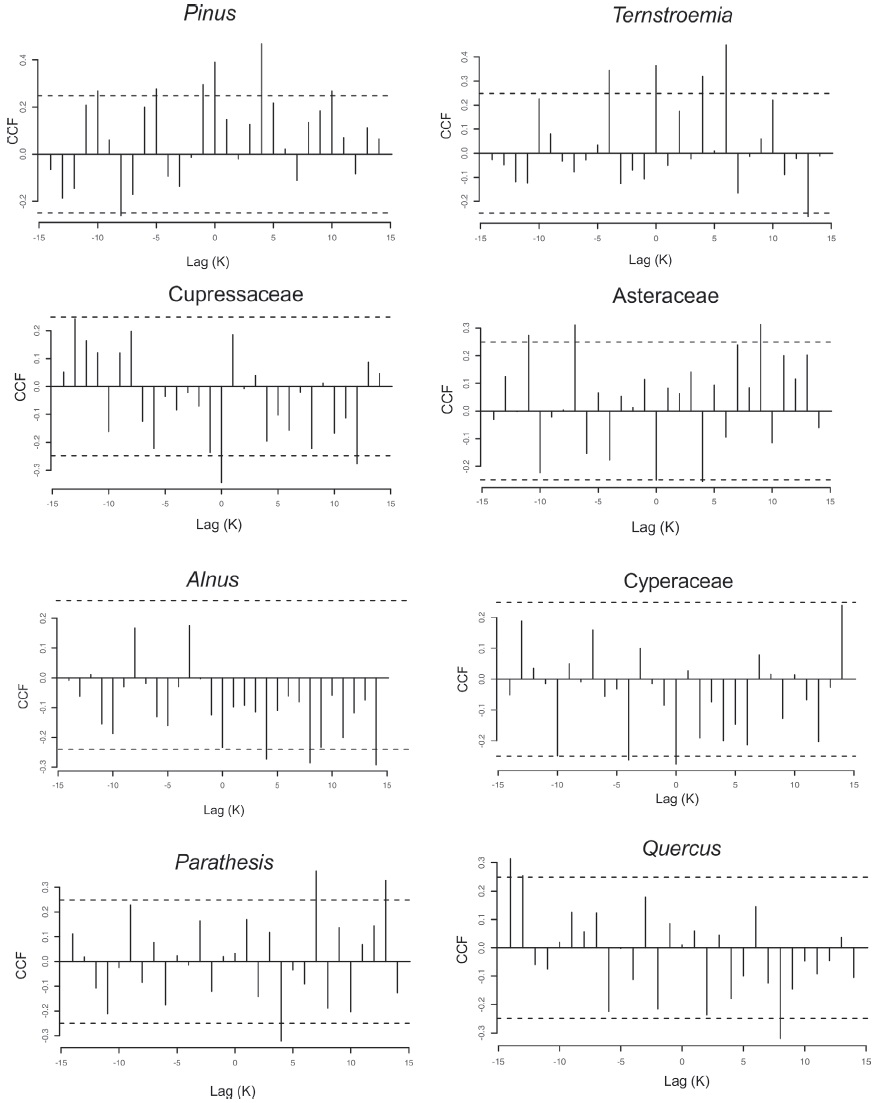
Figura 10 Diagramas de correlación cruzada entre la concentración de las micropartículas de carbón y la abundancia de polen de Pinus, Ternstroemia, Cupressaceae, Asteraceae, Alnus, Cyperaceae, Parathesis y Quercus. Los ejes verticales muestran los coeficientes de correlación, las líneas oscuras fuera de la línea punteada son significativas (p < 0.05). El eje horizontal muestra retrasos y avances en años. Los rezagos (lag positivos) corresponden con un retraso en una serie de tiempo. Un desfase negativo corresponde con los avances en una serie de tiempo. Cada lag tiene un intervalo de 25 años.
Discusión
Los resultados de este estudio proveen información a partir del registro de polen fósil y los indicadores paleoambientales, sobre la permanencia y expansión del bosque de Pinus-Quercus-Abies. A partir de estas evidencias se puede inferir cuales han sido los factores que conducen la dinámica natural del bosque. Los resultados de nuestro estudio sugieren que los intervalos de aridez y los incendios forestales han sido un factor clave en la dinámica del bosque del sitio de estudio, probablemente inducidos por actividades culturales como la agricultura (Caldararo, 2002) e intensificado por el cambio climático, principalmente relacionado con el aumento de intervalos de aridez (Cerano-Paredes et al., 2015; Figueroa-Rangel et al., 2008; Nelson et al., 2006). Los incendios forestales han demostrado ser un componente ecológico importante en la prevalencia de los bosques de pino que favorecen su amplia distribución en el hemisferio Norte (Agee, 2000; Rodríguez-Trejo y Fulé, 2003). La mayoría de las especies de Pinus están adaptadas a regímenes particulares de fuego por lo que presentan diferentes síndromes de adaptación a este factor (Keeley, 2012). Sin embargo, regímenes de incendios forestales frecuentes suelen mermar sus poblaciones al ceder el paso a otro grupo tolerante a los incendios como los pastos (Rzedowski, 2006). Lo que es evidente a partir de este estudio es que los incendios recurrentes no permitieron el establecimiento de un bosque dominado por latifoliadas; los factores responsables probablemente estén relacionados con la heterogeneidad fisiográfica del sitio de estudio, al encontrarse en las partes altas de las montañas donde las condiciones son propicias para el mantenimiento de las poblaciones de Pinus en áreas abiertas con pastos y matorrales secundarios (Quintana-Asencio y González-Espinosa, 1993).
Las evidencias encontradas en el registro polínico y los resultados de la ordenación directa RDA, muestran relación entre los taxones asociados a los incendios forestales através de las partículas de microcarbón fósil y la susceptibilidad magnética. Estos taxones son Pinus, Asteraceae, Poaceae, Phaseolus y Zea. En cambio, otros taxones como Abies y el grupo de pteridofitas sensu lato y epífitas requieren condiciones de mayor humedad y menor radiación solar (Martínez-Salas y Ramos, 2014) y se encuentran relacionadas con la presencia de mayor concentración de materia orgánica. En este sentido, la concentración de las partículas de microcarbón fósil que son indicadoras de incendios forestales parecen incidir en la abundancia de Pinus. Por otro lado, es evidente que los incendios ocurridos en el área de estudio han sido recurrentes durante los últimos 1,580 años. Sin embargo, cuando se presentan periodos de mayor humedad, Pinus es reemplazado por Alnus y en menor medida por Quercus, Abies, Parathesis, Fraxinus, Cupressaseae, Rosaceae y Myrtaceae indicando la presencia de un bosque mixto con algunos elementos arbóreos de bosque mesófilo de montaña. Resultados similares fueron registrados en el centro de México (Almeida-Leñero et al., 2005) donde algunos intervalos de mayor humedad favorecieron el florecimiento de la vegetación mesófila. También en la Sierra de Manantlán los periodos de mayor humedad registrados en esta región favorecieron la reducción del bosque de pino (Figueroa-Rangel et al., 2008), pero al retornar los periodos de aridez el bosque de pino se recuperó. Asimismo, Cerano-Paredes et al. (2015) encontraron una relación significativa en este mismo sitio entre las sequías y los periodos de incendios, por lo que el fuego se considera un factor clave para la dominancia del bosque de Pinus, relacionado con los intervalos de aridez.
Los resultados de este estudio indican que la abundancia de Pinus ha fluctuado durante los últimos ca. 1,580 años en el sitio de estudio como respuesta a los periodos de sequía y frecuencia de incendios. A través del tiempo Pinus ha sido el taxón dominante en gran parte de la secuencia, acompañado de taxones codominantes como Quercus, Abies y Alnus. Durante el periodo de ~370-405 d.C. (Zona C-1) se muestran condiciones relativamente secas inferido con base en el registro de Ti, carbonatos, Ca y susceptibilidad magnética. Estos indicadores señalan que el bosque transitaba por un recambio de taxones impulsado principalmente como respuesta a los cambios ambientales.
El periodo de ~405-1183 d.C. (Zona C-2) correspondió con la ACM que, de acuerdo con la mayoría de las evidencias provenientes de diversos registros paleoclimáticos, este periodo fue generalmente más cálido que durante la PEH (Díaz et al., 2011; PAGES-2K-Consortium, 2013). Por ejemplo, la secuencia sedimentaria de la Laguna de Juanacatlán (Metcalfe et al., 2010) y el Lago de Santa María del Oro (Sosa-Nájera et al., 2010) ambos al occidente de México, muestran un patrón de condiciones secas durante la ACM con alternancia de condiciones húmedas al final de la misma. Este patrón también está presente en el bosque del área de estudio. El registro de Ca muestra condiciones secas en el periodo de ~800 a ~1075 d.C., que coincide con la primera parte de la ACM, donde se observa la expansión del bosque de pino. Este periodo de sequía ha sido identificado en varios registros de México (Hodell et al., 2001, 2005; Lozano-García et al., 2010) al final del periodo Clásico (850 a 950 d.C.), con implicaciones en el colapso de la cultura Maya (Gill et al., 2007). También hay evidencia de que las diferentes culturas mesoamericanas podrían haber enfrentado diferentes periodos de cambios abruptos en el clima (Bhattacharya et al., 2015; Rodríguez-Ramírez et al., 2015), siendo en la mayoría de ellas los intervalos de sequías el factor más importante en las principales transiciones culturales. En esta región del occidente de México los patrones normales del clima pueden ser modificados por el desplazamiento estacional de la ZITC. En el verano del Hemisferio Norte se desplaza hacia el norte, este movimiento de sur a norte puede resultar en periodos de intensas lluvias o sequías extremas (Magaña et al., 2003; Sosa-Nájera et al., 2010). Otros mecanismos que podrían crear condiciones más secas o húmedas en esta región son los cambios en la frecuencia e intensidad de ENOS (Magaña et al., 2003; Metcalfe et al., 2010). La frecuencia de los huracanes y tormentas tropicales del Pacífico también son condiciones que pueden modificar el equilibrio de humedad en esta área (Vargas-Rodríguez y Platt, 2012). En algunos registros, las sequías muestran un patrón recurrente o cíclico (Figueroa-Rangel et al., 2008) lo que podría explicar los valores fluctuantes de los registros geoquímicos en el sitio de estudio. Estas condiciones parecen incidir en la vegetación de forma diferencial en Pinus, Quercus y Abies.
La correlación positiva entre Pinus y Ternstroemia con las partículas de microcarbón fósil, que cambian en sincronía sugiere que los incendios forestales en el área de estudio están relacionados con algunos periodos de expansión en el bosque. Estudios actuales sobre la dinámica del bosque de Pinus (González-Espinosa et al., 1991; Richardson y Bond, 1991) indican que este tipo de vegetación se establece en claros abiertos posteriores a los incendios forestales de baja intensidad, desmontes agrícolas o la explotación forestal. Mientras que Alnus y Parathesis se establecieron 100 años después de un aumento en la concentración de las partículas de microcarbón fósil, lo que implica que son elementos arbóreos de etapas tardías en la sucesión en bosque de alta montaña.
Es posible que los cambios ambientales estén relacionados con la dinámica del bosque de Pinus-Quercus-Abies. En nuestro registro Abies presenta los mayores porcentajes de la secuencia durante ~1048-1075 d.C. Posteriormente, se infieren condiciones de mayor humedad en el periodo de ~1102 a ~1156, el cual se asemejan con las condiciones en el registro de Ti del lago de Juanacatlán y el lago de Santa María del Oro, ambos sitios ubicados al occidente de México (Metcalfe et al., 2010; Rodríguez-Ramírez et al., 2015; Sosa-Nájera et al., 2010). Con la disminución de Pinus ocurrió un aumento de otros taxones leñosos como Parathesis, Ostrya-Carpinus, Xylosma y las familias Cupressaceae y Leguminosae, mientras que los taxones herbáceos como Cyperaceae también aumentaron, sobre todo Polypodiaceae y Aspleniaceae, grupos taxonómicos característicos del bosque mesófilo de montaña (Rzedowski, 1996).
El útimo periodo de ~1183 al presente, la zona C-3a refleja un periodo de transición de la ACM a la PEH con mayor variabilidad ambiental y recambio de taxones con la disminución en el componente herbáceo y el incremento en algunos taxones arbóreos como Pinus, Alnus, Ternstroemia y Xylosma. Sin embargo, muestra un patrón con 2 principales eventos de enfriamiento en la zona C-3b (~1421-1561 y ~1701-1842 d.C.) que se infieren por el incremento en los porcentajes de Pinus y Abies, así como la disminución de Quercus. Sin embargo, las condiciones ambientales son distintas en ambos periodos. En nuestro registro muestra condiciones más húmedas al principio de la PEH asociado con la reducción en los valores de Ca y el aumento en las concentraciones de Ti. En cambio, al final de la PEH muestra condiciones más secas y frías entre ~1701-1842 d.C. inferido con base en los mismos indicadores. Algunos registros históricos de México señalan condiciones extremas de sequías entre los siglos XVI y XIX, teniendo consecuencias severas en la agricultura, con la pérdida de cosechas, crisis sociales y la propagación de epidemias mortales en la población del centro de México (Contreras-Servín; 2005; Florescano et al., 1980; Therrell et al., 2004). Esta variabilidad ambiental puede estar relacionada con los ciclos de actividad solar (Spörer y Maunder) (Haug et al., 2003; Hodell et al., 2001). Los registros de Lago Verde en los Tuxtlas sugieren que los niveles del lago y la vegetación respondieron a este forzamiento solar y que además se puede tratar de un elemento importante que controla la variabilidad climática a escala de décadas y centurias en la región tropical del Atlántico norte (Lozano-García et al., 2007).
De acuerdo con nuestros datos, a partir de secuencias de sedimentos dentro del bosque, se proporcionan evidencias de la respuesta local de los bosques, por lo que nuestro trabajo sugiere que los distintos tipos de vegetación no responden unidireccionalmente ante el cambio global incluso durante los mismos periodos de tiempo. En su lugar, la evidencia señala patrones complejos en las respuestas de la vegetación durante los periodos de cambio ambiental, por lo que pueden ofrecer nuevos escenarios para evaluar los impactos del cambio climático a nivel local.
La evidencia palinológica como el registro de taxones indicadores de actividades culturales (Amaranthaceae y Asteraceae), así como taxones específicos de cultivares como Zea a partir de ~451 d.C. y Phaseolus a los ~1021 d.C., indica la existencia de impacto humano en el área de estudio. Los datos arqueológicos de la época prehispánica de esta región señalan la existencia de numerosos asentamientos indígenas dispersos en las partes bajas del área de estudio. Hacia el noroeste del valle de Mascota, Jalisco, se tienen indicios de poblaciones indígenas posiblemente Huicholes desde aproximadamente 100 a.C. (Cupul-Magaña y Mounjoy, 2012; Kelly, 1980), hacia el sureste en Colima aproximadamente entre 1200-800 a.C. (Mounjoy, 1994). Además, se destaca el hallazgo de Zea mexicana subsp. mexicana (teocintle de Chalco) y Zea mexicana subsp. parviglumis (teocintle del Balsas) cerca del volcán de Colima y la cuenca del Balsas, que posiblemente fue una zona de domesticación del maíz del occidente de México (Benz y Benz, 1990; Pollard, 1997). También hay evidencia de erosión del suelo por el aumento y marcada inestabilidad de la susceptibilidad magnética, además de la disminución en la materia orgánica, así como un régimen de incendios forestales frecuentes. Estos indicadores sugieren cambios importantes en la vegetación, probablemente asociados con disturbios humanos por el aclareo en los bosques para la agricultura (Noble y Dirzo, 1997), por lo que se infiere que la ocupación humana en el sitio de estudio ha estado presente durante algunos intervalos de tiempo, principalmente cuando las condiciones ambientales eran favorables.
La disminución de Pinus ocurrida al final del periodo ~1100-1200 d.C. registrado en nuestra secuencia, así como el aumento de otros taxones característicos de la vegetación herbácea de terrenos abiertos como Poaceae, Asteraceae, Cyperaceae y la presencia de polen de Zea, coincide con valores elevados de Ca y susceptibilidad magnética los cuales son considerados como indicadores de periodos secos y erosión respectivamente. Tales indicadores probablemente sugieren la presencia de actividades antropogénicas llevadas a cabo en el área de estudio. La historia de las diferentes culturas registradas para el occidente de México indica que las poblaciones indígenas eran afectadas por diversos procesos sociales que operaban en el centro de México, por lo que sus sociedades eran dinámicas con patrones complejos de interacción entre las poblaciones de la costa, la sierra del occidente y las poblaciones de las tierras altas del centro de México (Flannery y Marcus, 1994). Tal vez la complejidad de estos patrones se vean reflejados en los diferentes procesos ecológicos que muestra el registro palinológico del sitio de La Cumbre de Guadalupe. Sin embargo, el componente ambiental debió ser decisivo en la movilidad de las poblaciones de las partes altas de la sierra hacia las partes bajas cuando las condiciones del clima se tornaban desfavorables. Por ejemplo, la transición del ACM a la PEH pudo ser un momento difícil para los pobladores de las partes altas de la sierra debido a que en este periodo se presenta un aumento de algunos taxones leñosos y pteridofitas, lo que indica la recuperación del bosque.
Durante algunos periodos de la última zona de nuestro registro palinológico, está presente el polen de Phaseolus y Zea, por lo que sugiere el establecimiento de los pobladores hacia las partes altas en el área. Seguramente enfrentaban condiciones de sequía en las partes bajas de la sierra, debido a que éstas fueron generalizadas en diferentes zonas de nuestro país (Endfield y O´Hara, 1997; Florescano et al., 1980; Stahle et al., 2011).
Con la llegada de los españoles y la actividad minera, que caracterizó la actividad económica de las localidades aledañas al sitio de estudio en los últimos 2 siglos (Jiménez, 1995), el desarrollo de las actividades agrícolas en las partes altas de la sierra debieron implicar el aumento en el desmonte de las masas forestales; la evidencia de ello se observa en el registro palinológico con la disminución en la riqueza palinológica a partir de ~1927 d.C. y la alta concentración de micropartículas de carbón en ~1912 d.C. Posteriormente, se observa el incremento de los elementos como Ti, Fe, Zn y Pb, además del aumento en la susceptibilidad magnética que pueden estar asociados a mayores tasas de erosión y probablemente relacionadas con estas actividades.

