











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La biogeografía, entendida como el estudio de la distribución de los seres vivos en el espacio y a través del tiempo, constituye actualmente una de las disciplinas más activas en la biología evolutiva. Tiene como objetivos principales reconocer los patrones de distribución de las especies y taxones supraespecíficos, proponer hipótesis acerca de los procesos que los causaron y proporcionar un sistema de regionalización biótica del planeta (Morrone, 2004, 2005). En los últimos años, ha tomado un papel muy importante en la biología de la conservación, debido a que permite analizar a la biodiversidad desde una perspectiva histórica (Álvarez-Mondragón y Morrone, 2004; Escalante, 2009), lo cual resulta útil en la planeación ambiental para dar prioridad a los sitios de conservación, permitiendo proteger áreas que no serían consideradas con otros tipos de análisis (Arriaga-Cabrera et al., 1998).
El análisis de parsimonia de endemismos (PAE, por sus siglas en inglés) y el análisis de trazos, son 2 de los métodos más utilizados para identificar patrones de distribución con fines de conservación biológica. El PAE permite clasificar áreas en un cladograma a través de la solución más simple, basada en la presencia de taxones compartidos derivados (Nihei, 2006). Mediante este análisis se han podido proponer regionalizaciones que permiten categorizar a las áreas en términos de su biota (Escalante, 2009). Dichas áreas al presentar una combinación única de especies, resultan útiles en la planeación ambiental para priorizar sitios de conservación (Posadas y Miranda-Esquivel, 1999).
Por otra parte, un análisis panbiogeográfico parte de la construcción de trazos individuales de organismos filogenéticamente distantes, los cuales constituyen las coordenadas primarias de un taxón en el espacio, es decir, la unión de los puntos donde éste ha sido registrado (Crisci et al., 2000). A partir de la coincidencia de 2 o más trazos individuales, de taxones no relacionados filogenéticamente y con mecanismos de dispersión diferente, es posible visualizar patrones generales, conocidos como trazos generalizados o componentes bióticos (Humphries y Parenti, 1999), que representan biotas ancestrales fragmentadas por eventos geológicos o tectónicos (Craw, 1988). A todo este proceso se lo conoce como hipótesis de homología biogeográfica primaria (Morrone, 2001). La intersección de 2 o más de estos componentes da lugar a los nodos biogeográficos, áreas compuestas que permiten delinear límites biogeográficos (Craw, Grehan y Heads, 1999), derivadas de la convergencia de biotas con diferente origen espacial. Tal es el caso del nodo ubicado en América Central (Humphries y Parenti, 1999), y que constituye la zona de transición de las biotas neártica y neotropical. El análisis panbiogeográfico constituye una excelente herramienta para identificar áreas para su posible conservación (i.e., los nodos biogeográficos), al contener una alta riqueza de especies y una gran diversidad de relaciones filogenéticas y geográficas (Álvarez-Mondragón y Morrone, 2004).
Otros análisis que se incorporan en biogeografía para dar prioridad de conservación a las áreas son el análisis de complementariedad que permite identificar en un número reducido de áreas la máxima diversidad biológica posible y el análisis de diversidad filogenética que favorece el establecer prioridades entre los grupos de especies que se evalúan, de acuerdo con la máxima diversidad jerárquica representada, otorgando mayor importancia a los taxones con categorías superiores (Álvarez-Mondragón y Morrone, 2004; Torres-Miranda y Luna-Vega, 2006). Ambos criterios representan un requisito indispensable para la biología de la conservación.
Las macroalgas se dividen en 2 clases: macroalgas enanas (menores a 10 cm de longitud) y grandes macroalgas (mayores a 10 cm de longitud). Estas últimas, debido a su talla, dan lugar a hábitats biogénicos naturales y son refugio para una gran diversidad de organismos marinos (Connell, 2005). Además, representan materia prima sumamente importante en las industrias química (extracción de agar y carragenina), alimenticia, cosmética y papirogénica (Vilchis, 2015). A pesar de su importancia biológica y económica, dichos organismos no han sido objeto de estudios biogeográficos o de conservación en el país (Pedroche y Sentíes, 2003). Esto se debe en parte a que los criterios de conservación se han enfocado, principalmente, en la protección de áreas en las que se encuentran especies de vertebrados, invertebrados, pastos marinos y manglares; organismos conspicuos, de importancia económica y cultural, o por ser taxones catalogados en alguna categoría de riesgo (Escalante, 2003; Conabio-Conanp-TNC-Pronatura, 2007).
Dado lo anterior y tomando en cuenta que las divisiones que conforman al grupo de las macroalgas (Chlorophyta, Rhodophyta y Ochrophyta) no son un grupo natural, el objetivo del presente trabajo es identificar los patrones de distribución de las grandes macroalgas en el golfo de México y Caribe mexicano, a través de un análisis de parsimonia de endemismos (PAE) y uno de trazos, integrando un análisis de complementariedad y otro de diversidad filogenética, con la finalidad de encontrar áreas de importancia para la conservación de este grupo de organismos y que sean coincidentes con los sitios prioritarios para la conservación de la biodiversidad marina (SPCBM), propuestos por la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio) y la Comisión Nacional de Áreas Naturales Protegidas (Conanp), y de este modo, contribuir al conocimiento de este grupo de organismos y al entendimiento de su historia evolutiva.
Materiales y métodos
Se conjuntó una base de datos compuesta por 2,076 registros de todas las especies de grandes macroalgas reportadas para el golfo de México y Caribe mexicano, se obtuvo información referente a las localidades y entidades federativas donde se registraron, así como las coordenadas geográficas para cada localidad, registrando un total de 139 localidades únicas. Para la construcción de esta base se utilizó información bibliográfica obtenida a partir de los principales catálogos taxonómicos de macroalgas (Dreckmann, 1998; Littler y Littler, 2000; Ortega et al., 2001; Wynne, 2011) y registros de especímenes de recolecta albergados en AlgaeBase (Guiry y Guiry, 2016), la cual es la base de datos digitales colaborativa más importante en línea.
Para el PAE se trazó sobre un mapa del área de estudio una cuadrícula de 2° de latitud por 2° de longitud, utilizando el programa QGis v. 2.2.0 (Quantum GIS Development Team, 2014). En el mismo mapa se identificaron y se nombraron los cuadrantes que contenían registros de grandes macroalgas (Fig. 1). Posteriormente, se elaboró una matriz de cuadrantes vs. especies, codificando con “1” la presencia y con “0” la ausencia de las especies en cada cuadrante, además se incorporó un área hipotética para el enraizamiento del árbol, consistente en ausencias para todos los taxones. Esta matriz se analizó con el algoritmo Nona (Goloboff, 1999) en la interface WinClada (Nixon, 2002) bajo una rutina de máxima parsimonia, usando la opción de búsqueda heurística y una estrategia TBR+TBR (Tree Bisection and Reconnection + Tree Bisection and Reconnection). Una vez obtenidos los cladogramas más parsimoniosos se generó un cladograma de consenso estricto.
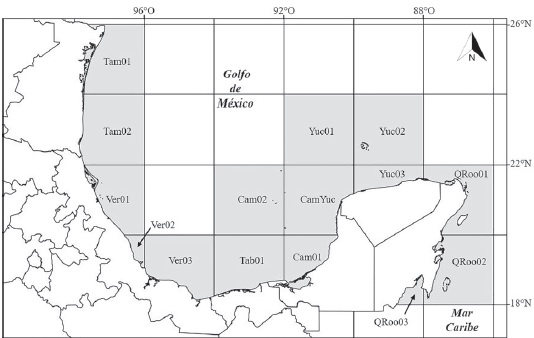
Figura 1 Mapa del golfo de México y Caribe mexicano que muestra, en color gris, los cuadrantes utilizados en los análisis.
Para el análisis de trazos se utilizaron las especies registradas en el área de estudio, exceptuando las que tuvieran una distribución amplia (i.e., que se distribuyan a lo largo de ambos litorales) o bien que están restringidas a un solo cuadrante. Se modificó la base de datos original utilizando OpenOffice 4 (hoja de cálculo), de tal manera que, sin eliminar información, solo quedara la entrada taxonómica y la georreferenciación. Esta información se exportó a formato dBaseIII, para posteriormente ser visualizadas en el sistema de información geográfica ArcView 3.2 (ESRI, 1999). Una vez obtenidas y cartografiadas las localidades puntuales de las 41 especies, se obtuvieron los trazos individuales, uniendo las localidades mediante el principio de distancia mínima, utilizando la extensión Trazos2004 (Rojas, 2004) en ArcView 3.2.; los archivos generados fueron transformados a formato Shapefile. Con la misma extensión, se construyeron los trazos generalizados, a partir de la congruencia de 2 o más trazos individuales, una vez delimitados se identificaron los nodos biogeo-gráficos.
Se contabilizó el número total de especies contenidas en cada cuadrante del área de estudio, considerando también el número de órdenes, familias y géneros de grandes macroalgas (Tabla 4), con la finalidad de conocer una primera aproximación de la diversidad de estos grupos en la región. Con base en la información anterior, se llevó a cabo un análisis de complementariedad, el cual permite encontrar pares de áreas que en conjunto alberguen el mayor número de especies posible, comparándolas unas con otras. Se utilizó el índice propuesto por Colwell y Coddington (1994), que cuantifica el grado de disimilitud en la composición de especies de las áreas que se comparan, mediante la siguiente formula:
Tabla 1 Especies exclusivas (sinapomorfías) que definen a los 9 grupos del PAE.
| Grupo | Cuadrante | Especies |
|---|---|---|
| I | Todos exceptuando QRoo03 | Acanthophora spicifera, Gelidium pusillum e Hypnea musciformis |
| II | Tam01, Tam02, Ver01, Ver02, Ver03, Tab01, Cam01, Yuc02, Yuc03, QRoo01, QRoo02 | Gracilaria blodgettii, G. cervicornis, G. cylindrica, Grateloupia filicina e Hydropuntia caudata |
| III | Tam01, Tam02, Ver01, Ver02, Ver03, Cam01, Yuc02, Yuc03, QRoo01, QRoo02 | Bryopsis plumosa, Dictyota ciliolata, Digenea simplex, Hydropuntia cornea, Jania capillacea, Sargassum filipendula, Spyridia filamentosa, Ulva flexuosa y U. lactuca |
| IV | Tam01, Tam02, Ver01, Ver02, Ver03, Yuc02, Yuc03, QRoo01, QRoo02 | Padina pavonica y Spatoglossum schroederi |
| V | Ver01, Ver02, Ver03, Yuc02, Yuc03, QRoo01, QRoo02 | Caulerpa sertularioides |
| VI | Yuc02, Yuc03 | Sporochnus pedunculatus |
| VII | Ver01, Ver02, Ver03, QRoo01, QRoo02 | Chnoospora minima y Cymopolia barbata |
| VIII | Ver02, Ver03, QRoo01, QRoo02 | Amasia multifida |
| XI | QRoo01, QRoo02 | Laurencia filiformis, Palisada flagellifera y Yuzurua poiteaui |
Tabla 2 Cuadrantes con especies únicas (autapomorfías).
| Área (cuadrante) | Especies |
|---|---|
| Norte del Sistema Arrecifal Veracruzano (Ver02) | Trichogloea requienii |
| Sur del Sistema Arrecifal Veracruzano (Ver03) | Dudresnaya crassa y Rhodogorgon ramosissima |
| Sur de Quintana Roo (QRoo01) | Gracilaria cuneata, Hypoglossum tenuifolium, Udotea abbottiorum, U. dixonii, y Wrightiella tumanowiczii |
| Centro de Quintana Roo (QRoo02) | Dictyota crenulata |
Tabla 3 Prioridad de conservación de acuerdo a los 4 criterios establecidos en este trabajo y a las categorías de importancia de los sitios prioritarios para la conservación de la biodiversidad marina (SPCBM) propuestos por Conabio-Conanp-TNC-Pronatura, (2007).
| Cuadrante | Criterios establecidos en este estudio | SPCBM de acuerdo a su importancia | |||||
|---|---|---|---|---|---|---|---|
| Especies exclusivas (sinapomorfías) | Nodo biogeográfico | Mayor riqueza filogenética | Complementariedad de áreas |
Importante | Muy importante | Extrema importancia | |
| Yuc02 | • | • | • | • | • | ||
| QRoo01 | • | • | • | • | |||
| Ver02 | • | • | • | ||||
| Yuc03 | • | • | • | ||||
| QRoo02 | • | • | • | • | |||
| Tam02 | • | ||||||
| Ver03 | • | ||||||
| Tam01 | |||||||
| Ver01 | |||||||
| Tab01 | |||||||
| Cam01 | |||||||
| Cam02 | |||||||
| CamYuc | |||||||
| Yuc01 | |||||||
Tabla 4 Diversidad de cada uno de los cuadrantes analizados.
| Cuadrante | Núm. de especies (%) | Núm. de géneros (%) | Núm. de familias (%) | Núm. de órdenes (%) |
|---|---|---|---|---|
| QRoo1 | 105 (88) | 53 (88) | 27 (90) | 15 (88) |
| Yuc02 | 70 (58) | 42 (70) | 25 (83) | 14 (82) |
| QRoo02 | 68 (46) | 40 (53) | 20 (67) | 12 (82) |
| Ver02 | 55 (57) | 32 (67) | 20 (67) | 14 (71) |
| Ver03 | 54 (45) | 39 (65) | 23 (77) | 13 (76) |
| Ver01 | 51 (43) | 34 (57) | 19 (63) | 13 (76) |
| Tam02 | 48 (40) | 33 (55) | 19 (63) | 12 (71) |
| Yuc03 | 47 (39) | 33 (55) | 20 (67) | 12 (71) |
| Cam01 | 43 (36) | 28 (47) | 19 (63) | 11 (64) |
| Tam01 | 37 (31) | 26 (43) | 16 (53) | 11 (64) |
| CamYuc | 17 (14) | 15 (25) | 12 (40) | 9 (53) |
| Cam02 | 14 (12) | 10 (16) | 8 (27) | 7 (41) |
| Yuc01 | 14 (12) | 9 (15) | 8 (27) | 7 (42) |
| Tab01 | 14 (12) | 12 (20) | 8 (27) | 5 (29) |
| QRoo03 | 13 (11) | 12 (20) | 7 (23) | 5 (29) |
donde A: número de especies presentes en el área 1, B: número de especies presentes en el área 2, j: número de especies presentes en ambas áreas.
El valor del índice de complementariedad varía entre 0 y 1; si su valor es igual a 0, son áreas no complementarias, indicando que las áreas contienen las mismas especies. Cuando el valor es igual a 1 no tienen especies en común, es decir, son áreas que se complementarían entre sí por las especies que cada una presenta.
Con la intención de analizar si los esfuerzos actuales de conservación coinciden con la diversidad de grandes macroalgas, se superpusieron en un mapa los cuadrantes con mayor riqueza filogenética, los que presentaron especies exclusivas únicas, los nodos biogeográficos y las áreas con mayor complementariedad. Los cuadrantes donde 2 o más de estos criterios coincidieron, se compararon con el polígono de los sitios prioritarios para la conservación de la biodiversidad marina.
Resultados
Se registraron 118 especies de grandes macroalgas válidas taxonómicamente para el golfo de México y Caribe mexicano, de las cuales 59 pertenecen a la división Rhodophyta, 34 a Chlorophyta y 25 a Ochrophyta. Del total de especies, Quintana Roo presentó 108, siendo el estado con mayor riqueza específica, seguido por Veracruz y Yucatán con 85 y 73 especies respectivamente. Asimismo, Tabasco fue el estado con el menor número de especies reportadas (14).
El PAE produjo 2 cladogramas igualmente parsimoniosos, con una longitud de 309 pasos, un IC = 0.38 y un IR = 0.53. Se generó un consenso estricto, el cual tuvo una longitud de 311 pasos, un IC = 0.38 y IR = 0.53. En éste se reconocieron 9 grupos de áreas basados en la presencia de especies exclusivas (i.e., sinapomorfías) (Fig. 2, Tabla 1). Es posible observar un clado sustentado por 9 especies (clado III), el cual muestra una relación entre las siguientes áreas: centro de Campeche (Cam01), norte y sur de Tamaulipas (Tam01, Tam02), arrecife Alacranes (Yuc02), costa centro de Yucatán (Yuc03), norte de Veracruz (Ver01), Sistema Arrecifal Veracruzano (SAV), (Ver02, Ver03), norte y centro de Quintana Roo (QRoo01, QRoo02). De los 15 cuadrantes considerados en este análisis, 4 presentaron especies únicas restringidas a un solo cuadrante, es decir autapomorfías (Tabla 2). Tal es el caso del clado VI que agrupa la región norte de la península de Yucatán (Yuc02, Yuc03) por la presencia de una sola especie (Sporochnus pedunculatus).

Figura 2 Cladograma de consenso estricto mostrando los 9 grupos definidos por la presencia de especies exclusivas (puntos negros) (véase especies en las Tablas 1 y 2).
En el análisis de trazos se obtuvieron 40 trazos individuales, 1 por cada especie, cuya superposición resultó en 2 trazos generalizados, 1 para el golfo de México y otro para el Caribe mexicano (Fig. 3a, b). La intersección de ambos resultó en un nodo biogeográfico ubicado en la región norte de la península de Yucatán (Fig. 3c). Las especies que sustentan a cada trazo generalizado se mencionan a continuación.

Figura 3 Trazos generalizados y nodo biogeográfico: a, trazo generalizado del golfo de México; b, trazo generalizado del Caribe mexicano; c, nodo biogeográfico.
Trazo generalizado del golfo de México: abarca desde el norte de Tamaulipas al noroeste de la península de Yucatán. Construido con los trazos individuales de Caulerpa ashmeadii, Chondria atropurpurea, Cladophora laetevirens, Cymopolia barbata, Eucheuma isiforme, Gracilaria tikvahie, Hypnea valentiae, Osmundaria obtusiloba, Rosenvingea intricata, Solieria filiformis, Sporochnus pedunculatus, Ulva flexuosa y U. prolifera.
Trazo generalizado del Caribe mexicano: desde el noroeste de la península de Yucatán hasta el sur de Quintana Roo y está construido con los trazos individuales de Amphiroa tribulus, Caulerpa microphysa, Cladophora longicaulis, Codium decorticatum, Chondria atropurpurea, C. cnicophylla, Dictyopteris jamaicensis, D. justii, D. plagiogramma, D. polypodioides, Dictyota pinnatifida, Gracilaria cuneata, Halimeda incrassata, H. monile, H. simulans, Halymenia floresii, Hydropuntia crassissima, Laurencia filiformis, Palisada flagellifera, Penicillus dumetosus, Rhipocephalus oblongus, Sargassum acinarium, S. ramifolium, Udotea occidentalis, U. flabellum, U. dixonii y Yuzurua poiteaui.
Las especies de grandes macroalgas reportadas en este trabajo se encuentran distribuidas en 60 géneros, 30 familias y 17 órdenes. El cuadrante con mayor riqueza específica fue QRoo01, que corresponde al norte del estado de Quintana Roo, mientras que los que tuvieron menor número fueron QRoo03 y Yuc01 (Tabla 4). De acuerdo con los porcentajes de las 4 jerarquías taxonómicas para cada cuadrante, los más altos corresponden al cuadrante QRoo01, seguido por Yuc02, QRoo02 y Ver02 (Tabla 4).
El análisis de complementariedad identificó que los cuadrantes Tam02 y Yuc02 son los que mejor se complementan, al presentar un ICC de 0.81, lo que indica que comparten pocas especies, permitiendo reunir en este par de áreas un total de 85 especies de grandes macroalgas de las 118 presentes en el golfo de México y Caribe mexicano.
Comparando los resultados obtenidos en los diferentes análisis realizados, es posible apreciar que el cuadrante Yuc02, que comprende al conjunto de islas de origen coralino conocidas como arrecife Alacranes al noroeste de la península de Yucatán, es la única región donde se superponen los 4 criterios establecidos (Tabla 3): mayor diversidad filogenética, especies exclusivas (sinapomorfías) presentes, presencia de un nodo biogeográfico y mayor complementariedad de áreas. En el norte y centro de Quintana Roo (QRoo01, QRoo02), el centro de Yucatán (Yuc03) y el norte del Sistema Arrecifal Veracruzano (SAV, Ver02), se superpusieron solo 2 de los 4 criterios utilizados (Tabla 3).
De acuerdo con las categorías de conservación propuestas por Conabio-Conanp-TNC-Pronatura (2007) para los sitios prioritarios para la conservación de la biodiversidad marina (SPCBM), arrecife Alacranes y el norte del SAV coinciden con 2 de “extrema importancia (EI)” (Fig. 4). Mientras que el norte de Quintana Roo coincide con un sitio de “EI” y 3 de categoría “muy importante (MI)”, el centro de Quintana Roo con 2 de “EI” y uno “MI”. Finalmente, el centro de Yucatán coincide con un sitio catalogado como “MI” para la conservación marina (Fig. 4).
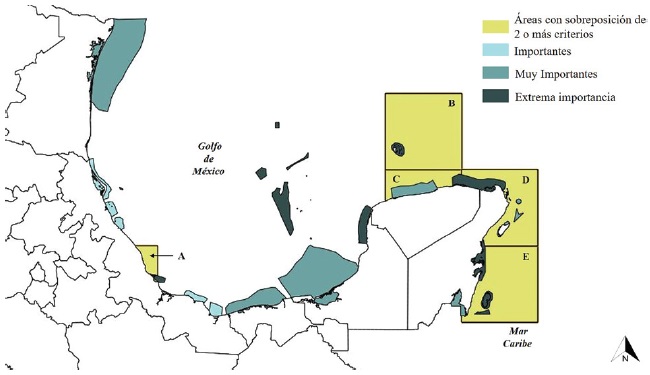
Figura 4 Coincidencia de los sitios prioritarios para la conservación de la biodiversidad marina (SPCBM) propuestos por Conabio-Conanp-TNC-Pronatura, (2007), con los cuadrantes que presentaron la sobreposición de 2 o más de los criterios establecidos en el presente estudio. A: norte del Sistema Arrecifal Veracruzano; B: arrecife Alacranes; C: centro de Yucatán, D: norte de Quintana Roo; E: centro de Quintana Roo. En un gradiente de colores se muestran los SPCBM de acuerdo a su importancia.
Discusión
Los resultados obtenidos a través del PAE muestran una relación entre los cuadrantes que integran el clado III del cladograma de consenso estricto, que puede ser explicada a partir de evidencia oceanográfica y geológica. Por un lado, la conectividad existente entre las 2 grandes áreas (golfo de México y mar Caribe), debido a la entrada de la corriente del Caribe proveniente del sur a través del estrecho de Yucatán. Dicha corriente termina en el noroeste de la península de Yucatán, para dar lugar a la corriente de Lazo, la cual genera giros al interior del golfo de México que fluyen hacia la parte central del mismo (Fig. 5). Generando una trayectoria hacia el oeste, que impacta con el litoral del norte de Veracruz y el sur de Tamaulipas, dividiéndola en 2 nuevos flujos, uno hacia el norte con dirección al litoral de EUA y el segundo con dirección sur, directamente hacia el Sistema Arrecifal Veracruzano (Fig. 5; González et al., 1991).
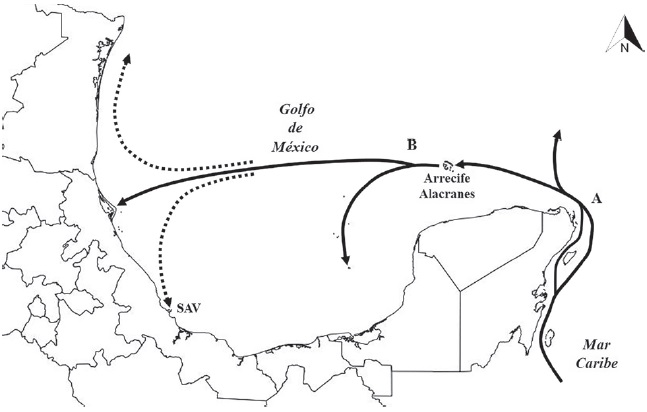
Figura 5 Circulación de las corrientes dentro del golfo de México y Caribe mexicano (modificada de Chávez-Hidalgo, 2009). A: corriente de Lazo; B: corriente del Caribe.
Bird et al. (2011) atribuyen una estrecha relación biótica entre estas 2 áreas, considerando la historia geológica de la apertura del golfo de México, iniciada hace aproximadamente 180 ma (Jurásico medio), con el desprendimiento de la península de Yucatán, aún sumergida del margen noroccidental del golfo en este periodo, desplazándose con un giro contrario a las manecillas del reloj, hasta su posición actual donde más recientemente emergió (Fig. 6). De acuerdo con esta hipótesis, en algún momento de la historia geológica del golfo, el norte de la península de Yucatán y la porción noroeste del mismo se encontraban unidas formando una probable área ancestral, este proceso pudo haber tenido gran influencia en la composición de la biota actual del golfo de México.

Figura 6 Desplazamiento de la península de Yucatán, aún sumergida, desde Texas dando un giro en sentido contrario de las manecillas del reloj, hasta su posición actual (modificada de Pindell et al., 1988).
Los componentes bióticos identificados en el análisis de trazos son resultado de 2 eventos geológicos subsecuentes a este desprendimiento, 1) el surgimiento del Istmo de Tehuantepec hace aproximadamente 50 ma, a principios del Eoceno y 2) la emersión de la península de Yucatán hace aproximadamente 19 ma, en el Mioceno medio (Bird et al., 2011). El primero podría explicar la fragmentación de las poblaciones compartidas entre el Pacífico mexicano y el golfo de México, mientras que el segundo da cuenta de la presencia de una barrera entre poblaciones de la cuenca del golfo y aquellas del mar Caribe. Dicha evolución geológica con la consecuente fragmentación de hábitat, sería el detonante para los procesos de radiación adaptativa y especiación, de tal modo que, justamente en el norte de la península de Yucatán se puede identificar una zona de transición o nodo biogeográfico, entre 2 biotas marinas o componentes bióticos diferentes: una proveniente del Caribe y otra propiamente del golfo, lo cual podría explicar la gran diversidad filogenética de macroalgas presente en esta área.
En la literatura se reporta que para el Atlántico mexicano, la zona del nodal presenta los valores más altos de riqueza y diversidad de otros grupos de algas como el género Laurencia (Tapia-Silva et al., 2015) y otros organismos como anélidos (Granados-Barba et al., 2003), moluscos (Castillo-Rodríguez, 2014; Escobar, 2004) y peces (Floeter et al., 2008), además de presentar los valores más altos de diversidad haplotípica de las especies de algas rojas Hydropuntia cornea e H. usneoides (Núñez et al., 2016). Este nodo se suma a los 2 reportados previamente para el territorio mexicano, un nodo continental centroamericano (Humphries y Parenti, 1999), cuya influencia se extiende hasta la Faja Volcánica Transmexicana, y un nodo marino identificado en el golfo de California (Aguilar-Aguilar y Contreras-Medina, 2001).
Dichos patrones de distribución han permitido, junto con los análisis de complementariedad y diversidad filogenética, establecer 4 criterios para la identificación de áreas de conservación marinas considerando la distribución de las grandes macroalgas (Tabla 3). A partir de éstos se señalan 5 áreas prioritarias para la conservación de estos grupos, principalmente el arrecife Alacranes en Yucatán, ya que representa un área de complejidad biótica y geológica, reflejada en la sobreposición de los 4 criterios. Aunque el análisis de complementariedad indique que el arrecife Alacranes y el sur de Tamaulipas son las áreas que en conjunto albergan el mayor número de especies (85), si a su vez se complementan con el norte del estado de Quintana Roo, se estarían conservando 110 de las 118 especies analizadas en este trabajo.
Aunque hasta el momento no hay información con respecto a especies de macroalgas en alguna categoría de riesgo nacional, se sabe que las grandes macroalgas constituyen un elemento fundamental en la estructura de las comunidades inframareales, al establecer ambientes biogénicos que contribuyen a la heterogeneidad de hábitats, permitiendo así el establecimiento, sobre ellas, de una biota rica en representantes de varios phyla, tanto procariontes como eucariontes (uni o multicelulares). Además, sirven como refugio para muchos animales (Cancino y Santelices, 1984; Connell, 2005), lo que permite la coexistencia de un número elevado de especies que da lugar a comunidades altamente diversas, con composiciones bióticas complejas. Esto se refleja de manera indirecta en la coincidencia de los resultados obtenidos en este trabajo, tanto con los SPCBM propuestos por Conabio-Conanp-TNC-Pronatura (2007), pero también con los resultados de riqueza y diversidad de otros organismos como los mencionados previamente.
Si bien es cierto que en este estudio no se consideró el intervalo de distribución completo de las especies de grandes macroalgas, es decir su distribución fuera de México, fue posible definir patrones de distribución y ofrecer algunas hipótesis sobre los procesos que los originaron, además de reconocer congruencias con los patrones de distribución de otros organismos. Sin embargo, dichas hipótesis deberán ser corroboradas continuando con el estudio de la distribución de las macroalgas en diversos contextos. En particular, es necesario llevar a cabo estudios histórico-evolutivos, filogenéticos y ecológicos con estos grupos para generar más información que permita confirmar hipótesis de homología biogeográfica primaria desde el punto de vista de la biogeografía evolutiva, entendiendo por homología biogeográfica primaria ese primer esbozo de relaciones históricas entre áreas, y que representa solo el primer paso dentro de un estudio de esta índole (Morrone, 2009).

