











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
En los últimos años el desarrollo urbano se ha incrementado rápidamente, lo que provoca una creciente demanda de energía. En la actualidad, esta es generada principalmente a través de combustibles fósiles, lo que conlleva la emisión de gases de efecto invernadero, causantes primordiales del calentamiento global (McKinney, 2002). Una de las alternativas es la obtención de energía renovable, como la eólica, la cual ha tenido auge en los últimos años (Atienza et al., 2011, Henestroza, 2008, Moryarty y Honnery, 2012). No obstante, se han registrado algunas consecuencias negativas, principalmente en ambientes templados, por el establecimiento y funcionamiento de parques eólicos, entre las que destacan la muerte de aves y murciélagos por colisiones, los ruidos (audible y ultrasónico) y el electromagnetismo (Cryan y Barclay, 2009, Henestroza, 2008, Kunz et al., 2007a).
Los estudios del impacto de los aerogeneradores sobre la fauna se han enfocado principalmente a las aves y recientemente a investigar las afectaciones a los murciélagos (Arnett et al., 2008). La metodología a nivel mundial se enfoca al conteo de cadáveres por aerogenerador por año o por megawatt generado (Hayes, 2013). La actividad de los murciélagos se ha evaluado a través de cámaras infrarrojas y detectores ultrasónicos; con estas técnicas se han determinado las diferencias de actividad por temporadas y su relación con la cantidad de cadáveres encontrados (Baerwald y Barclay, 2009, Horn et al., 2008, Johnson et al., 2004). Las investigaciones registran que las poblaciones de vespertiliónidos y molósidos que vuelan a grandes alturas tienen mayores impactos (Barros et al., 2015, Kunz et al., 2007b).
En regiones tropicales de Latinoamérica, sobre todo en México y Brasil, las evaluaciones en los impactos sobre murciélagos son escasas, a pesar del creciente desarrollo y aprobación de proyectos de parques eólicos (Barros et al., 2015, Bolívar-Cimé et al., 2016, Rodríguez et al., 2009, Rodríguez-Durán y Feliciano-Robles, 2015, Zolotoff-Pallais et al., 2012). Debido a las diferencias en diversidad de la región neotropical con la región neártica (mayor riqueza y menor abundancia; Jenkins, Pimm y Joppa, 2013), son necesarios estudios poblacionales, de diversidad, riqueza y patrones de actividad espacial y temporal para evaluar de manera más precisa el efecto que tienen los parques eólicos sobre los murciélagos en ambientes tropicales.
En este estudio se analizan las diferencias en la riqueza de murciélagos artropodívoros (Segura-Trujillo, Lidicker y Álvarez-Castañeda, 2016) y sus patrones de actividad en sitios dentro y fuera de parques eólicos por temporada. Se ha reportado que los murciélagos son sensibles a la perturbación y responden generalmente con la disminución en riqueza y abundancia (Jung y Kalko, 2010, Stahlschmidt y Brül, 2012, Williams y Perfecto, 2011). Por lo tanto, predecimos que la riqueza y la abundancia de los murciélagos artropodívoros serán menores en las áreas con presencia de aerogeneradores, mientras que la actividad será mayor en sitios sin aerogeneradores y estará acotada a las primeras horas del atardecer, debido a que en ese periodo hay una mayor disponibilidad de presas (Rydell, Entwistle y Racey, 1996). La información generada podría ser utilizada para crear estrategias que disminuyan colisiones futuras de estos mamíferos en parques eólicos del neotrópico y apoyar el establecimiento de una mejor política de manejo y conservación.
Materiales y métodos
El área de estudio se ubica al sureste de México, en el istmo de Tehuantepec, Oaxaca. Es una de las zonas de mayor potencial eólico del país y del mundo, con vientos promedio de 7.7-8.5 m/s a 50 m sobre el suelo, calificados como excelentes para la producción de energía eólica y presentes durante todo el año (Elliott et al., 2004; Fig. 1). El clima es cálido subhúmedo (Aw) con una temperatura media anual de 22-26 °C. La precipitación media anual va de 1,000 a 1,200 mm (Trejo, 2004). En esta región existen grandes extensiones de terrenos con alto grado de fragmentación, utilizados para agricultura de riego y temporal y cría de ganado bovino. Posee escasos corredores de vegetación arbórea con huizaches (Acacia sp.) y huamúchiles (Phithecellobium dulce), y con pequeños remanentes de bosque tropical caducifolio. El mayor afluente de la zona es el río Tehuantepec. Además, existen canales de riego para irrigar los pastizales y cultivos agrícolas, principalmente de maíz y sorgo (Inegi, 2013).
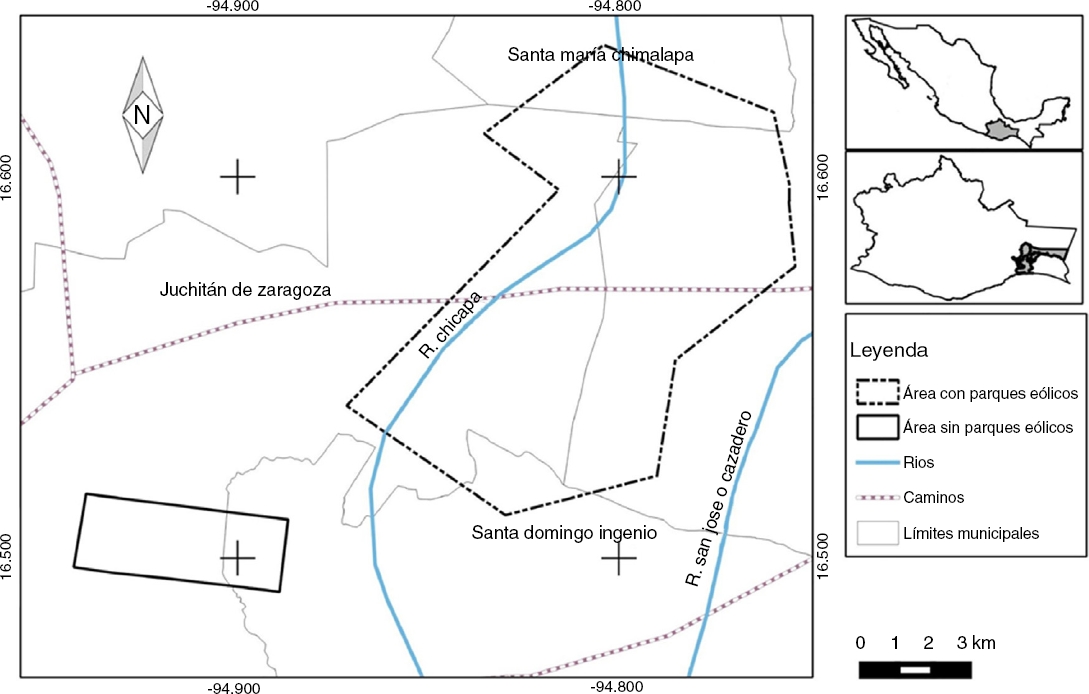
Figura 1 Localización de las áreas de estudio dentro de parques eólicos (DPE) y fuera de parques eólicos (FPE) en el istmo de Tehuantepec, Oaxaca, México.
Dentro de la zona de estudio se seleccionaron 2 áreas con diferente condición. La primera dentro de parques eólicos (DPE, 20 parques en la región, con un total de 1,882 aerogeneradores; Sener, 2016) y la segunda fuera de parques eólicos (FPE, Fig. 1).
Diseño de muestreo
Se establecieron 8 sitios de muestreo, 4 DPE (1 al 4) y 4 FPE (5 y 8). Desafortunadamente, durante el estudio 2 de los sitios FPE fueron invadidos por un nuevo parque eólico, razón por la cual solo se pudieron considerar datos de 2 sitios FPE (5 y 6). El trabajo se realizó en 2 ciclos de muestreo anuales durante la temporada seca (marzo a mayo) y la temporada lluviosa (junio a agosto) de 2013 y 2014, dando un total de 12 periodos de captura.
Para la recolecta de murciélagos con redes de niebla se utilizaron 8 y 12 redes (12 × 2.6 m; Avinet® Inc) durante 2 noches por sitio por mes. Las redes se colocaron a partir del crepúsculo vespertino (18:00 h) y permanecieron abiertas alrededor de 7 h diarias en posibles rutas de vuelo y en fragmentos de vegetación. El número total de noches para los sitios DPE fue de 48, mientras que para los sitios FDP fue de 24.
Los murciélagos capturados fueron identificados utilizando claves especializadas (Álvarez et al., 1994, Medellín et al., 2007). Se obtuvieron datos de edad (joven o adulto de acuerdo al grado de fusión de las falanges; Anthony, 1988), sexo y condición reproductiva (testículos escrotados, testículos inguinales, inactivo/preñada, lactante, poslactante); posterior a ello, los organismos fueron liberados en el mismo sitio de su captura. Algunos especímenes fueron recolectados y preparados como ejemplares de museo, principalmente molósidos y vespertiliónidos, de los cuales no se tuvo certeza de identificación al 100% en campo y de los que se obtuvieron grabaciones vocales. Los ejemplares de museo se depositaron en la Colección Mastozoológica (OAX.MA.026.0497) del Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Unidad Oaxaca (CIIDIR-OAXACA, IPN). La captura fue realizada bajo el permiso de colecta (Semarnat, FAUT-0037), otorgado a los autores por las autoridades correspondientes (Secretaría de Medio Ambiente y Recursos Naturales).
La grabación de firmas vocales de murciélagos se llevó a cabo con el detector de ultrasonido Anabat SD2 (Titley Electronics, Ballina, Australia; intervalo de frecuencia 4-200 kHz, sistema de división de frecuencia). El detector funcionó aproximadamente 8 h al día, a partir del crepúsculo (18:00 h), y hasta la 01:59 am, con una noche de grabación por sitio por mes. En algunos sitios no se completaron las horas de grabación debido a condiciones ambientales, presencia de lluvia o vientos > 30 m/s. El método de grabación fue pasivo modificado del propuesto por O’Farrell, Miller y Gannon (1999) y Weller y Zabel (2002). El detector se colocó con una inclinación de 45° a una altura de 30 cm. Los archivos obtenidos con el Anabat SD2 fueron analizados con el programa AnalookW versión 3.8 (Titley Scientific, Ballina, Australia).
El análisis consistió en la separación de los archivos útiles. Se consideró como archivo útil (un pase) aquel que tuviera al menos 2 llamados sucesivos de ecolocalización; de esta forma se eliminaron archivos con secuencias fragmentadas o con ruido ambiental (Fenton, 1970, Thomas, 1988). Por otro lado, los eventos de caza (EC) fueron determinados a partir de los trenes de alimentación (definidos como un aumento en la tasa de repetición de llamados; Griffin, Webster y Michael, 1960), que indican la posible captura de presas en cada sitio. Se consideró cada tren de alimentación como un evento independiente (MacSwiney, Clarke y Racey, 2009). Finalmente, se determinaron los horarios y picos de actividad de las familias de murciélagos identificadas; los horarios de actividad se determinaron a través del número de pases por noche agrupados en intervalos de una hora.
La identificación de especies se efectuó, en el caso de las especies de la familia Mormoopidae, con grabaciones de referencia de obtención propia. Las grabaciones se obtuvieron de individuos capturados en refugios cercanos a la zona de estudio (< 10 km de distancia) a través del método de liberación en mano (hand release) en áreas abiertas (e.g., Frick, 2013, Williams y Perfecto, 2011). Las especies restantes fueron identificadas por comparación de espectrogramas de regiones o estados del país y de localidades de Centroamérica (Gannon et al., 2004, Gannon et al., 2001, Jung et al., 2007, O’Farrell, 1997, O’Farrel y Miller, 1997, Orozco-Lugo et al., 2013, Rydell et al., 2002). En algunos casos no fue posible llegar a nivel de especie, por lo que se analizaron a nivel de género, o también considerados como sonotipos (grupos de especies con llamados similares; Bader et al., 2015), particularmente para las familias Molossidae y Vespertilionidae, que poseen patrones de sonido variables o pulsos similares entre especies (Szewczak, 2000, Walters et al., 2012).
Para la determinación taxonómica de especies, o en su caso de los sonotipos, se midieron 6 parámetros de cada llamado (Dur: duración, Fmax: frecuencia máxima, Fmin: frecuencia mínima, fmean: frecuencia media, Tk: tiempo de inicio del llamado a la inflexión o rodilla, y Fk: frecuencia de la inclinación o rodilla; O’Farrell et al., 1999). La visualización de llamados se realizó en una división de frecuencia 16, con el eje de tiempo (x) en intervalos divididos cada 10 ms y eje de frecuencia (y) en escala logarítmica (kHz).
Análisis de datos
Se generaron curvas de acumulación de especies y se determinó la eficiencia del muestreo para la captura en redes de niebla y detección ultrasónica. Por cada método de recolecta se realizaron matrices con los datos de presencia-ausencia, DPE y FPE, y se ingresaron al programa EstimateS ver.9.1 (Colwell, 2009). Los datos se combinaron 1,000 veces para obtener una distribución aleatoria, eliminar el efecto de orden de ingreso y suavizar la curva. La riqueza esperada se calculó a través de los estimadores no paramétricos Bootstrap y Chao 2. La completitud del inventario se obtuvo calculando el porcentaje de especies observadas respecto a las calculadas por los estimadores (Moreno y Halffter, 2000).
El esfuerzo de muestreo en redes de niebla se calculó por cada periodo para ambas temporadas (lluvias y secas) de 2013 y 2014. El total fue utilizado para estimar la abundancia relativa (AR), que se refiere al número de individuos colectados de cada especie, dividido por el número de m.red/hora total (Medellín, 1993). Para determinar diferencias en la captura de individuos con redes DPE y FPE y por temporadas se realizó la prueba no paramétrica de Kruskall-Wallis, por no cumplirse los supuestos de normalidad de los datos, a través del programa Infostat (Di Rienzo et al., 2016).
Para el análisis de los pases grabados y EC se realizó un modelo lineal generalizado (MLG; programa SPSS ver. 22) con la función de enlace Poisson y el estadístico de Wald (Mangeaud y Videla, 2005); la variable explicativa fue el número de conteos por cada sitio con 12 repeticiones (meses), es decir, 48 repeticiones para la condición DPE y 24 para FPE. Se compararon estos 2 factores (DPE, FPE) y se realizó la selección de modelos mediante el criterio de información de Akaike (AIC) y de máxima verosimilitud.
Finalmente, se usó una prueba de Kruskall-Wallis (Di Rienzo et al., 2016) para determinar diferencias entre los horarios de actividad con el programa Infostat. Para todos los casos se utilizó un nivel de significancia de p = 0.05.
Resultados
La riqueza general de especies de murciélagos artropodívoros de la zona estuvo compuesta por 13 especies y 4 géneros, agrupados en 4 familias. Se recolectaron 8 especies con redes de niebla, y se registraron 9 especies y 4 géneros por medio de detección ultrasónica (Fig. 2, Tabla 1, Tabla 2). En el área DPE se registraron 12 especies y 4 géneros, mientras que en el área FPE resultaron 10 especies y 4 géneros. Pteronotus personatus y Eptesicus sp. se presentaron solo en DPE, mientras que Molossus rufus solo se registró en sitios FPE.
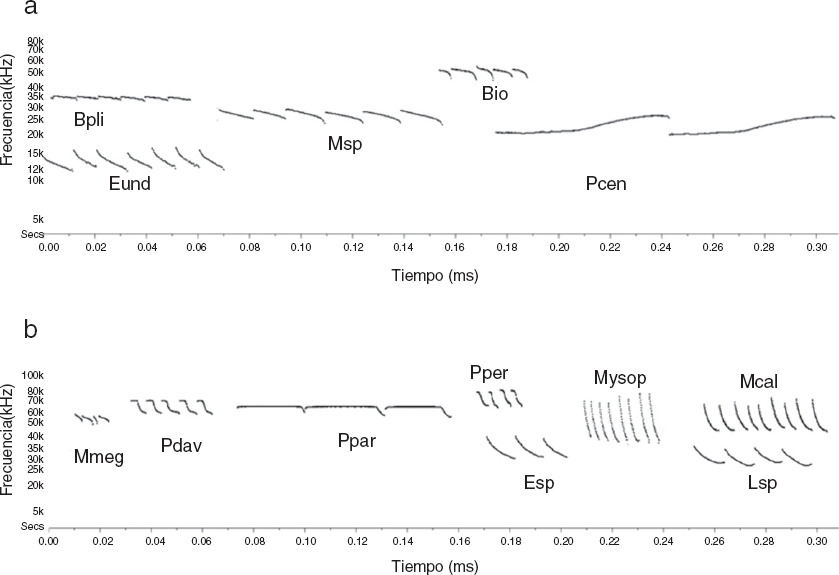
a) Familia Emballonuridae, Balantiopteryx io: Bio, Balantipteryx plicata: Bpli; familia Molossidae, Eumopsunderwoodi: Eund, Molossus sp: Msp, Promops centralis: Pcen.
b) familia Mormoopidae, Mormoops megalophylla: Mmeg, Pteronotus davyi: Pdav, Pteronotus parnellii: Ppar, Pteronotus personatus: Pper; familia Vespertilionidae, Eptesicus sp: Esp, Lasiurus sp.: Lsp, Myotis sp: Myosp, Myotis califonicus: Mcal.
Figura 2 Espectrogramas de las especies registradas a través del detector ultrasónico Anabat SD2 en el área DPE y FPE en el istmo de Tehuantepec, Oaxaca, México. Visualización de espectrogramas, eje y en kiloHertz (kHz), eje x en tiempo (10 ms).
Tabla 1 Registro de especies de murciélagos artropodívoros mediante detección acústica (A; número de archivos) y redes de niebla (RN; número de individuos), en sitios con presencia y ausencia de parques eólicos en el istmo de Tehuantepec, Oaxaca, México.
| Especie | Dentro de parques eólicos (DPE) | Fuera de parques eólicos (FPE) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Método | Ll-2013 | S-2013 | Ll-2014 | S-2014 | Ll-2013 | S-2013 | Ll-2014 | S-2014 | Total | ||
| Familia Emballonuridae | |||||||||||
| Balantiopteryx io | A | 0 | 2 (2) | 1 | 0 | 0 | 0 | 0 | 0 | 3 (2) | |
| Balantipteryx plicata | A | 7 | 7 | 4 | 14 | 2 | 8 | 2 | 6 | 50 | |
| Familia Molossidae | |||||||||||
| Eumops underwoodi | A | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 3 | |
| Molossus sp. | A | 193 (3) | 151 (88) | 218 (2) | 102 (1) | 52 (4) | 142 (1) | 215 (4) | 146 | 1,219 (103) | |
| Molossus molossus | RN | 0 | 1 | 1 | 2 | 0 | 2 | 0 | 1 | 7 | |
| Molossus rufus | RN | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | |
| Promops centralis | A | 2 | 0 | 0 | 3 | 0 | 0 | 0 | 2 | 7 | |
| Familia Mormoopidae | |||||||||||
| Mormoops megalophylla | A | 18 | 16 | 0 | 1 | 1 | 2 | 0 | 1 | 39 | |
| RN | 2 | 2 | 0 | 2 | 0 | 45 | 0 | 0 | 51 | ||
| Pteronotus davyi | A | 451 (1) | 143 (5) | 535 | 51 | 85 | 56 | 709 (2) | 178 | 2,208 (8) | |
| RN | 1 | 0 | 0 | 0 | 0 | 9 | 2 | 0 | 12 | ||
| Pteronotus parnellii | A | 16 | 4 | 35 | 7 | 1 | 44 | 1 | 10 | 118 | |
| RN | 11 | 0 | 5 | 2 | 0 | 1 | 0 | 0 | 19 | ||
| Pteronotus personatus | A | 11 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 14 | |
| RN | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | ||
| Familia Vespertilionidae | |||||||||||
| Eptesicus sp. | A | 0 | 11 (11) | 27 | 4 | 0 | 0 | 0 | 0 | 42 (11) | |
| Lasiurus sp. | A | 21 | 31 (11) | 15 | 34 (1) | 15 | 1 | 12 | 5 | 134 (12) | |
| Lasiurusintermedius | RN | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | |
| Myotis sp. | A | 61 | 116 (17) | 72 | 42 | 32 | 20 | 152 | 122 | 617 (17) | |
| Myotiscalifonicus | A | 3 | 0 | 0 | 0 | 60 (1) | 0 | 1 | 0 | 64 (1) | |
| Rhogeessaparvula | RN | 2 | 0 | 0 | 2 | 0 | 1 | 0 | 3 | 8 | |
| Individuos RN | RN | 16 | 3 | 6 | 9 | 0 | 59 | 2 | 5 | 100 | |
| Pases (EC) | A | 784 (4) | 483 (134) | 908 (2) | 260 (2) | 248 (5) | 273 (1) | 1,092 (6) | 470 (0) | 4,518 (154) | |
Ll: temporada lluviosa; S: temporada seca.
Los números entre paréntesis representan los eventos de caza (EC).
El arreglo taxonómico siguió a Ramírez-Pulido, González-Ruiz, Gardner y Arroyo-Cabrales (2014).
Tabla 2 Abundancia relativa (AR, individuos.redes/h) de murciélagos capturados en redes de niebla y el promedio (X̅ ± SD) de 6 parámetros medidos en los llamados de ecolocalización de las especies y géneros identificados a través del detector ultrasónico Anabat SD2 en el trópico mexicano.
| Especie | AR | Dur (ms) | Fmax (kHz) | Fmin (kHz) | Fmean (kHz) | Tk (kHz) | Fk (ms) |
|---|---|---|---|---|---|---|---|
| Familia Emballonuridae | |||||||
| Balantiopteryx io | 0 | 6.42 ± 1.87 | 52.87 ± 1.05 | 45.68 ± 1.26 | 50.68 ± 0.66 | 0.41 ± 0.39 | 52.30 ± 0.69 |
| Balantipteryx plicata | 0 | 7.24 ± 1.52 | 39.64 ± 0.33 | 38.41 ± 0.51 | 39.09 ± 0.34 | 0.24 ± 0.22 | 39.54 ± 0.32 |
| Familia Molossidae | |||||||
| Eumops underwoodi | 0 | 9.72 ± 1.70 | 15.43 ± 1.65 | 11.94 ± 0.50 | 13.25 ± 0.85 | 2.51 ± 1.28 | 13.82 ± 0.82 |
| Molossus sp. | 0 | 15.15 ± 2.31 | 27.34 ± 0.80 | 22.96 ± 1.17 | 25.36 ± 0.89 | 1.03 ± 0.80 | 26.95 ± 0.93 |
| Molossumolossus | 0.000058 | 0 | 0 | 0 | 0 | 0 | 0 |
| Molossusrufus | 0.000008 | 0 | 0 | 0 | 0 | 0 | 0 |
| Promopscentralis | 0 | 64.96 ± 2.24 | 25.95 ± 0.34 | 19.83 ± 0.23 | 22.43 ± 0.09 | 47.59 ± 5.24 | 24.56 ± 0.84 |
| Familia Mormoopidae | |||||||
| Mormoopsmegalophylla | 0.000423 | 3.39 ± 1 | 55.86 ± 1.24 | 49.79 ± 1.42 | 53.23 ± 1.8 | 0.22 ± 0.1 | 55.18 ± 0.81 |
| Pteronotusdavyi | 0.000099 | 5.69 ± 0.59 | 72.26 ± 0.49 | 59.58 ± 0.69 | 66.46 ± 0.74 | 2.25 ± 1.71 | 66.69 ± 4.75 |
| Pteronotusparnellii | 0.000157 | 27.47 ± 3.01 | 64.28 ± 0.34 | 59.11 ± 1.70 | 63.80 ± 0.36 | 2.11 ± 0.86 | 64.05 ± 0.30 |
| Pteronotuspersonatus | 0.000008 | 2.01 ± 1.37 | 80.62 ± 3.90 | 72.69 ± 7.61 | 76.14 ± 5.90 | 0.51 ± 0.63 | 76.86 ± 7.04 |
| Familia Vespertilionidae | |||||||
| Eptesicus sp. | 0 | 10.93 ± 0.25 | 41.39 ± 0.81 | 30.46 ± 0.42 | 33.67 ± 0.35 | 4.46 ± 0.62 | 33.86 ± 0.76 |
| Lasiurus sp. | 0 | 10.21 ± 1.33 | 34.47 ± 1.48 | 27.79 ± 0.93 | 29.69 ± 1.05 | 3.79 ± 1.04 | 30.04 ± 0.99 |
| Lasiurusintermedius | 0.000008 | 0 | 0 | 0 | 0 | 0 | 0 |
| Myotis sp. | 0 | 3.58 ± 0.81 | 70.94 ± 9.22 | 41.65 ± 2.17 | 50.02 ± 3.05 | 2.25 ± 0.57 | 46.05 ± 2.87 |
| Myotiscalifonicus | 0 | 5.24 ± 0.83 | 79.51 ± 3.42 | 52.80 ± 0.79 | 59.85 ± 1.56 | 2.83 ± 0.57 | 56.59 ± 1.32 |
| Rhogeessa parvula | 0.000066 | 0 | 0 | 0 | 0 | 0 | 0 |
Dur: duración; Fk: frecuencia de la inclinación o rodilla; Fmax: frecuencia máxima; Fmean: frecuencia media; Fmin: frecuencia mínima; Tk: tiempo de inicio del llamado a la inflexión o rodilla.
El arreglo taxonómico siguió a Ramírez-Pulido et al. (2014).
Para las redes de niebla, se obtuvo un esfuerzo de muestreo total de 127,872 m red/h; para 2013 el esfuerzo fue de 55,296 m red/h, y 72,576 m red/h para 2014. Por condición, en el área DPE el esfuerzo de muestreo fue de 85,240 m red/h, mientras que para el área FPE fue de 42,624 m red/h. En el caso de las temporadas, se registraron 63,936 m red/h para cada una.
Las curvas de acumulación de especies no alcanzaron la asíntota para ambas áreas. En el área DPE la riqueza estimada fue de 6.92 especies con Chao 2 y de 6.78 especies con Bootstrap (Sobs = 6), por lo que se registraron el 86.71 y el 88.50% de la riqueza de especies estimada, respectivamente. Para el área FPE, con los estimadores Chao 2 (Sobs = 7 y Sest = 8.38) y Bootstrap (Sobs = 7 y Sest = 8.74) se registraron el 83.53 y el 80.09% de la riqueza de especies esperada (Fig. 3b).

a) Muestreo a través de detección ultrasónica.
b) Muestreo a través de redes de niebla.
Singletons: especie registradas en una ocasión; Doubletons: especie registradas en 2 ocasiones.
Figura 3 Curva de acumulación de especies por condición (DPE y FPE) en el istmo de Tehuantepec, Oaxaca, México.
Se capturaron 100 individuos de 8 especies agrupados en 3 familias; la más capturada fue Mormoops megalophylla (n = 51). Molossus rufus, P. personatus y Lasiurus intermedius se capturaron solo en una ocasión (Tabla 2). En el área DPE se registraron 34 individuos, y Pteronotus parnellii y M. megalophylla tuvieron el mayor número de registros (70.6%). En el área FPE se capturaron 66 individuos, M. megalophylla y Pteronotus davyi fueron las especies más capturadas (84.8%). Al analizar las capturas entre las 2 temporadas, la mayor se observó durante la seca (n = 76) respecto a las lluvias (n = 24). En el análisis entre años, en el 2013 se registró el mayor número de ejemplares (n = 78) a diferencia de 2014 (n = 22, Tabla 1). Sin embargo, no se encontraron diferencias significativas por condición (DPE, FPE) (H = 0.09, g.l. = 1, p = 0.75), ni por temporadas (H = 3.89, g.l. = 2, p = 0.23).
Para la detección ultrasónica, las curvas de acumulación de especies para ambas áreas alcanzaron la asíntota en el doceavo mes de recolecta. La riqueza estimada en el área DPE fue de 13 especies con Chao 2 y de 13.54 con Bootstrap (Sobs = 13), por lo que se registró el 100% y 96.01% respectivamente de la riqueza de especies estimada. En el área FPE la riqueza estimada fue de 9 con Chao 2 y 9.42 con Bootstrap (Sobs = 9) con registró de 100% y 95.54% respectivamente (Fig. 3a).
Debido a que la prueba de Kruskall-Wallis no mostró diferencias significativas en el número de pases entre los sitios con aerogeneradores (H = 4.37, g.l. = 3, p = 0.22) ni la prueba de Friedman entre los sitios sin ellos (X2 = 1.60, g.l. = 1, p = 0.38), se trataron como 2 áreas independientes: dentro de parques eólicos (DPE) y fuera de parques eólicos (FPE).
Se obtuvieron 345 horas de grabación, 221 para el área DPE y 124 para el área FPE, en ambas se registraron 4,518 pases, que sirvieron para identificar 9 especies y 4 géneros pertenecientes a 4 familias (Tabla 1). Para la familia Emballonuridae se identificaron 2 especies (Balantiopteryx io y Balantipteryx plicata). De la familia Molossidae se obtuvieron registros de 2 especies y 1 género (Eumops underwoodi, Promops centralis y Molossus sp.). Para la familia Mormoopidae se registraron 4 especies (Mormoops megalophylla, P. davyi, P. parnellii y P. personatus). En la familia Vespertilionidae se obtuvo una especie y 3 géneros (Myotis californicus, Eptesicus sp., Lasiurus sp. y Myotis sp.) (Fig. 2a y b, Tabla 2).
Al analizar los datos del número de pases, utilizando los factores DPE y FPE/temporada seca y temporada lluviosa, se generó solamente un modelo, por lo que no se reportan datos de Akaike (AIC) y devianza.
En el área DPE se identificaron 2,435 pases (54%), que corresponden a 9 especies y 4 géneros. En el área FPE se obtuvieron 2,083 pases (46%) de 6 especies y 3 géneros. No se observaron diferencias significativas de la actividad con el número de pases entre condiciones a través del MLG (χ2 de Wald = 2.52, p = 0.11, Tabla 3). Seis especies y 3 géneros son compartidos en ambas áreas, mientras que 3 especies y un género son exclusivos de los sitios DPE; el género que estuvo presente en ambas áreas a lo largo de todos los meses fue Molossus sp. (Tabla 1).
Tabla 3 Estimación de parámetros con un modelo lineal generalizado de distribución Poisson.
| Parámetro | B | Error estándar | 95% de intervalo de confianza de Wald | Contraste de hipótesis | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Inferior | Superior | χ2 de Wald | g.l. | Sig. | ||||||
| (Interceptación) | 4.045 | 0.3017 | 3.453 | 4.636 | 179.771 | 1 | 0.000 | |||
| [Condición A] | −0.537 | 0.3378 | −1.199 | 0.125 | 2.527 | 1 | 0.112 | |||
| [Condición B] | 0a | |||||||||
| [Temporada 1] | 0.713 | 0.3361 | 0.054 | 1.372 | 4.503 | 1 | 0.034 | |||
| [Temporada 2] | 0a | |||||||||
Condición A: dentro de parques eólicos; Condición B: fuera de parques eólicos; Temporada 1: temporada de lluvias; Temporada 2: temporada de secas.
a Definido en cero porque este parámetro es redundante.
Durante la temporada de lluvias 2013 se registró el 22.84% de pases y para secas, el 16.73%. Para lluvias del 2014, el 44.27% de pases, y para secas, el 16.16%. Balantiopteryx plicata, Molossus sp., M. megalophylla, P. davyi, P. parnellii, P. personatus, Myotis sp. y Lasiurus sp. estuvieron presentes en ambas temporadas de 2013 y 2014 (Tabla 1). Las diferencias en los porcentajes de pases registrados entre temporadas, tuvieron diferencias significativas de acuerdo al MLG (χ2 de Wald = 4.50, p = 0.03; Tabla 3).
En cuanto a los eventos de caza (EC), se registraron 154 pases para 3 especies y 4 géneros, los cuales estuvieron presentes en el 3.4% de archivos del total. El género y la especie con más EC fue Molossus sp. (n = 103, 66.9%) y P. davyi (n = 8, 5.19%), respectivamente, en contraposición M. californicus y Eptesicus sp. con 1 y 11 (0.65% y 7.14%) tuvieron el menor número de EC; mientras que 6 especies, no registraron EC (Tabla 1).
Se registró el mayor porcentaje de EC en el área DPE (92.2%), en la temporada seca (88.96%) y en el año 2013 (93.50%). A partir del análisis con un MLG, se observaron diferencias significativas para condición (χ2 de Wald = 28.49, g.l. = 1, p = 0.00) y temporadas (χ2 de Wald = 56.77, g.l. = 1, p = 0.00; Tabla 4).
Tabla 4 Estimación de parámetros con un modelo lineal generalizado de distribución Poisson para eventos de caza (EC).
| Parámetro | B | Error estándar | 95% de intervalo de confianza de Wald | Contraste de hipótesis | |||||
|---|---|---|---|---|---|---|---|---|---|
| Inferior | Superior | χ2 de Wald | g.l. | Sig. | |||||
| (Interceptación) | 0.831 | 0.1707 | 0.497 | 1.166 | 23.725 | 1 | 0.000 | ||
| [Condición A] | 0.970 | 0.1818 | 0.614 | 1.327 | 28.498 | 1 | 0.000 | ||
| [Condición B] | 0a | ||||||||
| [Temporada 1] | −1.183 | 0.1570 | −1.491 | −0.875 | 56.778 | 1 | 0.000 | ||
| [Temporada 2] | 0a | ||||||||
Condición A: dentro de parques eólicos; Condición B: fuera de parques eólicos; Temporada 1: temporada de lluvias; Temporada 2: temporada de secas.
a Definido en cero porque este parámetro es redundante.
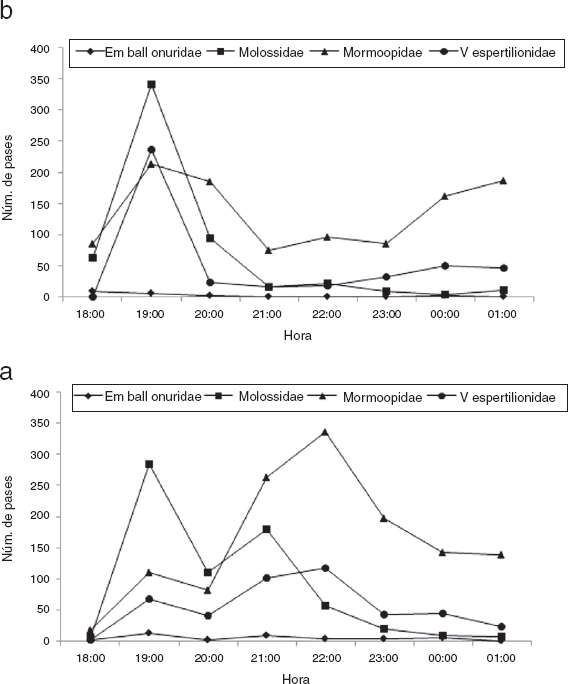
La representación de la actividad de los murciélagos por horas se agrupó por familia y área, dado que mostraba patrones similares por especies y por sitios (Fig. 4). Para el área DPE se registró actividad a las 19:00 h y entre las 22:00 y las 23:00 h. En el área FPE se observaron 2 picos de actividad, a las 19:00 y entre a las 24:00 y la 01:00 h. La familia Emballonuridae tuvo escasa representación, por lo que no se observó un patrón definido. La Molossidae tuvo mayor actividad entre las 19:00 y las 20:00 h en ambas áreas. La Mormoopidae mostró mayor actividad en el área DPE de las 19:00 a las 20:00 h y de las 22:00 a las 23:00 h (Fig. 4a). De manera similar, en el área FPE se registraron 2 picos de actividad (Fig. 4b). La Vespertilionidae mostró actividad en el área DPE a lo largo de la noche, sin picos bien definidos; en contraste, en los sitios FPE se observó un pico a las 19:00 h. En el análisis de los horarios de actividad se observó que el área DPE, las 19:00 y las 20:00 h acumularon el 18.88 y el 11.04% (n = 853, 499) de pases, respectivamente, mientras que en el área FPE las 19:00 h acumuló el 17.66% de pases (n = 798). A través del análisis de los intervalos de actividad se encontró que hay diferencias significativas entre los pases registrados por hora (H = 48.78, g.l. = 7, p = 0.0001).
Discusión
La fauna de murciélagos en esta región está constituida por al menos 70 especies (Briones-Salas, Cortés-Marcial y Lavariega, 2015), de las cuales aproximadamente el 51% pertenecen al gremio artropodívoro. Las especies y géneros registrados de este gremio en el estudio (n = 17) representan el 36% para la región. Bolívar-Cimé et al. (2016) registraron 20 especies de murciélagos artropodívoros en una zona con presencia de parques eólicos colindante a la zona de estudio; de estas, 10 coinciden con este estudio, 3 de nuestros registros no aparecen en sus listas y las especies restantes no se registraron. Esto posiblemente se deba a que el sitio de estudio de los autores antes mencionados se encuentra con una mayor cobertura vegetal, relativamente más cerca de un bosque tropical caducifolio en buen estado de conservación (Pérez-García, Meave y Salas, 2010).
En sitios con parques eólicos para Estados Unidos y Europa, el número de especies registradas mediante detección ultrasónica es menor que las reportadas aquí (Amorim, Rebelo y Rodrigues, 2012, 3 especies; Baerwald y Barclay, 2009, 5 especies; Fiedler, 2002, 6 especies; Minderman, Pendlebury, Pearce-Higgins y Park, 2012, 4 especies). Lo anterior es de esperarse dada la mayor riqueza de especies de la región neotropical, donde se ubica la zona con parques eólicos estudiada (Jenkins et al., 2013). No obstante, coincide con el número de registros obtenidos en los Chimalapas, Oaxaca, a 35 km al norte del área de estudio (9 especies y 4 géneros; Kraker-Castañeda, Santos-Moreno y García-García, 2013).
En estudios de la región neotropical, mediante el método de redes en Brasil, se identificaron 10 especies, 7 artropodívoras (Barros et al., 2015), mientras que en Puerto Rico se registraron 11, 5 artropodívoras (Rodríguez-Durán y Feliciano-Robles, 2015); en ambos estudios el número de especies identificadas es menor a las registradas en este trabajo. Lo anterior se atribuye al sesgo que tienen las redes en la captura de especies; se ha documentado que el uso de detectores de ultrasonido incrementa el registro de especies un 30 a 40% (MacSwiney et al., 2008, Pech-Canché et al., 2010). Esto resulta importante si se considera que la mayoría de las especies en las que se han registrado altas tasas de mortalidad por choque con las aspas de los aerogeneradores pertenecen a especies artropodívoras (Bolívar-Cimé et al., 2016, Kunz et al., 2007b). De la misma manera, se han realizado otros trabajos con mamíferos en la región del istmo de Tehuantepec, como el de Cervantes y Yépez (1995) (26 especies de murciélagos, 8 artropodívoras) y el de López, Lorenzo, Barragán y Bolaños (2009) (28 especies, 9 artropodívoras), que reportan más especies que las de este trabajo, debido a que no solo consideran registros resultado de trabajo de campo sino también registros de colecciones.
Destaca la presencia de P. davyi y el género Molossus como las especies más registradas en este trabajo. P. davyi se ha relacionado a vegetación de bosque (Jung y Kalko, 2011); sin embargo, el área de estudio posee escasa vegetación, y la presencia de esta especie puede atribuirse a la cercanía (10 km) con un área comunal protegida (Ojo de agua Cerro del Tolistoque), con remanentes de bosque tropical caducifolio en buen estado de conservación y que tiene una cueva que funciona como refugio de mormópidos y embalonúridos (Briones-Salas, Peralta-Pérez y García-Luis, 2013). Para el caso del género Molossus, esto coincide con lo encontrado por otros autores (Jung y Kalko, 2010, Jung y Kalko, 2011, Kraker-Castañeda et al., 2013, Bolívar-Cimé et al., 2016), quienes encontraron que en sitios urbanos, pastizales y con parques eólicos en Gamboa (Panamá), Santa María Chimalapas y Juchitán (México) la actividad de molósidos es alta. Lo anterior se debe a que algunas especies de este género poseen cierta plasticidad adaptativa a condiciones de perturbación (Jung y Kalko, 2011). Además, es posible, que la presencia de parques eólicos permita que se mantengan en la zona, debido a que los miembros de esta familia habitualmente forrajean en áreas abiertas, necesarias para el óptimo funcionamiento de los aerogeneradores (Schnitzler y Kalko, 2001).
La mayor cantidad de registros de especies de las familias Mormoopidae y Molossidae en los sitios con parques eólicos coincide con lo reportado por otros autores en Brasil y Puerto Rico (Barros et al., 2015, Rodríguez-Durán y Feliciano-Robles, 2015) debido a la afinidad neotropical de las especies de esas familias; por otro lado, se contrapone con lo encontrado en parques eólicos de la región neártica, donde Vespertilionidae tiene mayor presencia (Amorim et al., 2012, Arnett et al., 2008).
La actividad de los murciélagos medida a través de pases no mostró diferencias significativas entre áreas, lo que se contrapone con lo esperado al inicio del trabajo. En otros estudios tampoco se han observado diferencias entre sitios con ciertos niveles de perturbación, como es el caso de lo observado (Kraker-Castañeda et al., 2013) en sitios con selva y pastizales y lo encontrado en áreas con fragmentos de bosque, bosque continuo y huertos con asentamientos humanos, en Veracruz, México (Estrada, Jiménez, Rivera y Fuentes, 2004).
El no encontrar diferencias significativas en la actividad entre condiciones podría responder a que ambas zonas de estudio (DPE y FPE) se ubican en una matriz de agricultura y ganadería con procesos significativos de fragmentación y pérdida en la cobertura vegetal. Estas condiciones se han mantenido desde hace más de 30 años, por lo que actualmente la vegetación nativa representa menos del 50% en la zona (Muñoz et al., 2016, Velázquez et al., 2003). Es posible que estas perturbaciones afectaran de manera importante la composición original de especies y solo sobrevivan aquellas que se adaptaron a estos cambios. Debido a lo anterior, la presencia de parques eólicos podría no tener grandes repercusiones en la actividad de los murciélagos, a diferencia de los cambios en la vegetación (e.g., Jung y Kalko, 2010, Jung y Kalko, 2011). Adicionalmente, los resultados del análisis de actividad entre condiciones sugieren que el paisaje es usado de manera similar para el movimiento y la alimentación por las poblaciones de artropodívoros. Sin embargo, no se descarta que exista mortandad debido a choques con turbinas, y aunque en este estudio no se realizó búsqueda de cadáveres de murciélagos, en un parque eólico cercano al área de este estudio se registró una tasa de mortandad de 2.18-20.20 individuos por turbina y año (Bolívar-Cimé et al., 2016).
En el caso de las temporadas, la mayor actividad se presentó durante la temporada de lluvias, y es posible que esto se deba a que durante esta existe una mayor cantidad de alimento. Lo que se contrapone con lo encontrado en otros estudios (López-González et al., 2016, MacSwiney et al., 2009), que registraron una mayor actividad durante la temporada seca pero en sitios con presencia de cuerpos de agua, que puede atribuirse a la necesidad de hidratación de los murciélagos y a la disponibilidad de alimento.
Por otro lado, se obtuvo un alto porcentaje de EC (n = 154), comparado con lo obtenido en granjas del Reino Unido (1,747 pases, 47 EC; Wickramasinghe, Harris, Jones y Vaughan, 2003) y en la isla de Gotland en el mar Báltico (6 pases, 0 EC; Rydell y Wickman, 2015). En contraposición, se obtuvo un bajo porcentaje de EC, si se compara con lo obtenido en sitios con cenotes y sin ellos en la región neotropical (2,111 pases, 665 EC en total; MacSwiney et al., 2009). A pesar de que el área DPE registró casi la totalidad de EC y que la diferencia entre condiciones es significativa, el bajo número de estos en relación con otras regiones, sugiere que la zona es usada en mayor medida como un área de paso de las especies hacia otros sitios con mayores recursos, como canales y/o ríos. Esto porque los sitios de grabación no estuvieron cercanos o en cuerpos de agua, condición que ha sido reportada de importancia para los murciélagos, por la presencia de insectos de algunos de los cuales se alimentan (Fiedler, 2002).
Los horarios de actividad de los murciélagos en zonas con parques eólicos no han sido medidos anteriormente en el neotrópico. Los horarios de la familia Vespertilionidae y Molossidae estuvieron acotados a las primeras horas de la noche, lo que coincide con lo reportado por Agosta, Morton, Marsh y Kuhn (2005) en refugios en Maryland y Pennsylvania en EUA. En cuanto a la familia Mormoopidae, se observaron 2 picos de actividad, entre las 22:00 y las 23:00 h y entre las 12:00 y la 01:00 h. Es probable que un análisis detallado de la composición de la dieta de estas familias pudiera mostrar alguna relación con este comportamiento. Las diferencias de actividad entre horas coinciden con lo que se ha encontrado para la actividad de algunos insectos de los cuales se alimentan los murciélagos de estas familias (Rydell et al., 1996). Por tanto, al considerar que existe mortalidad DPE de acuerdo a lo encontrado en la región (Bolívar-Cimé et al., 2016) y que la actividad de murciélagos en el área de estudio se concentra en las primeras horas del atardecer y la noche (de 19:00 a 20:00 h y de 22:00 a 23:00 h), se sugiere la programación de altos al funcionamiento de los aerogeneradores en estos horarios, lo que podría beneficiar a las poblaciones de murciélagos estudiadas.
Finalmente, se considera que otros factores pueden influir sobre la actividad y los EC, por mencionar algunos: variables ambientales (viento, humedad y temperatura; Erickson y West, 2002, Lewis, 1993) y configuración del paisaje (presencia de caminos, cuerpos de agua, fragmentos de bosque, cultivos [Ávila-Flores y Fenton, 2005, Burles et al., 2009]). Ello muestra futuras líneas de investigación en la zona, para descartar los posibles efectos de los aerogeneradores sobre las especies de murciélagos artropodívoros.

