











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
La delimitación y la identificación de especies son actividades complementarias y recíprocas que han estimulado la labor de los sistemáticos durante los últimos años. Por una parte, la delimitación permite discernir entre linajes evolutivos distintos, conduciendo eventualmente al descubrimiento de nuevas especies (Wiens, 2007); por otra parte, la identificación permite la asignación de individuos dentro de las categorías definidas previamente (Edwards y Knowles, 2014). Estas actividades son particularmente importantes dentro de la sistemática puesto que permiten descubrir y describir a las especies, como condición primordial para darles nombre y clasificarlas (Morrone, 2013), pero también para formular hipótesis sobre sus relaciones filogenéticas (Wiens, 2007). La sistemática, como parte de la biología comparada, ostenta una notable influencia sobre disciplinas como la biogeografía, la ecología y la biología de la conservación (Reid y Carstens, 2012; Rissler y Apodaca, 2007; Wiens, 2007; Wiens y Servedio, 2000).
Los ejercicios de delimitación y de identificación son complejos en menor o mayor medida, dependiendo del grupo taxonómico, de las fuentes de información empleadas y de los métodos de análisis implementados (Edwards y Knowles, 2014; Rissler y Apodaca, 2007; Sites y Marshall, 2003; Wiens, 2007). La identificación de especies se apoya principalmente en información morfológica y es común el uso de métodos automatizados de análisis (Cope, Corney, Clark, Remagnino y Wilkin, 2012), pero también se admite información genética, ecológica, ambiental o de distribución geográfica y es posible contrastar distintos métodos de análisis (Cope et al., 2012; DeSalle, Egan y Siddall, 2005; Scheinvar, Toledo y Sánchez-Mejorada, 1976).
Neobuxbaumia Backeb. (Cactaceae: Cactoideae: Echinocereeae) es un género endémico de México, que se distribuye desde el centro (Puebla, Hidalgo, Querétaro y San Luis Potosí) hacia las vertientes del golfo de México (Tamaulipas y Veracruz) y del océano Pacífico (Jalisco, Colima, Michoacán, Guerrero y Oaxaca) (Bravo-Hollis, 1978). Las especies se caracterizan por presentar hábito arbóreo, con un porte mayor a 5 m, tallos columnares con ramificación o simples, cristales prismáticos en la hipodermis, de 15 a 40 costillas excepto en N. euphorbioides (8 a 10), areolas relativamente próximas entre sí, con una depresión interareolar, espinas distribuidas regularmente en una serie radial y otra central; flores campanular-infundibuliformes cubiertas por brácteas, generalmente carentes de cerdas y tricomas; frutos verdes a pardos, con restos persistentes del perianto y dehiscencia vertical; semillas de 2-4 mm de largo y pardo-negras (Arias y Terrazas, 2006; Arias, Gama-López, Guzmán-Cruz y Vázquez-Benítez, 2012; Arroyo-Cosultchi, Terrazas, Arias y López-Mata, 2010; Bravo-Hollis, 1978). Los tratamientos taxonómicos más recientes reconocen hasta 9 especies de Neobuxbaumia (Guzmán-Cruz, Arias y Dávila, 2003; Hunt, 2006) (apéndice 1); algunas de ellas pueden identificarse con facilidad, mediante la comparación de atributos vegetativos como el número de costillas, el tamaño y la forma de las areolas. Otras presentan conflictos en su identificación, particularmente en las regiones donde convergen sus áreas de distribución, debido a la similitud de las estructuras vegetativas. Estos conflictos persisten por la escasez y por la duración efímera de las estructuras reproductivas que son indispensables para su identificación, como flores, frutos y semillas, tanto en el campo como en las colecciones botánicas (Arias et al., 2012; Arroyo-Cosultchi et al., 2010; Bravo-Hollis, 1978). Por ejemplo, el estudio comparativo de Neobuxbaumia multiareolata, N. macrocephala, N. polylopha y N. scoparia realizado por Bravo-Hollis, Scheinvar y Sánchez-Mejorada (1970, 1971a, 1971b, 1973) reveló la existencia de combinaciones particulares en caracteres vegetativos y reproductivos que permitieron distinguirlas. Un análisis fenético realizado por Scheinvar et al. (1976), utilizando diferentes coeficientes de similitud y técnicas de agrupamiento permitió identificar 7 especies de Neobuxbaumia, a partir de un conjunto de caracteres morfológicos, distribución geográfica y altitud. Estos últimos autores no consiguieron distinguir a N. squamulosa, debido a la carencia de flores y frutos, identificándola equivocadamente como N. mezcalaensis y dejando para un tiempo posterior su descubrimiento (Scheinvar y Sánchez-Mejorada, 1990).
Un caso particular fue la controversia en la delimitación de N. mezcalaensis y N. multiareolata; mientras Bravo-Hollis et al. (1973) señalan que se trata de 2 especies distintas, para Hunt et al. (2006) se trata de una misma especie. El análisis multivariado realizado por Arroyo-Cosultchi et al. (2010) probó la ausencia de fenotipos intermedios y que estos taxa se agrupan claramente en 2 clases distintas; estos mismos autores encontraron 4 caracteres de la areola del tallo vegetativo (forma y tamaño de areola, tamaño de espina central y patrón de distribución de espinas radiales), más 9 caracteres reproductivos, con los valores de carga más altos. A partir de estos resultados será relevante conocer el valor de los caracteres de la areola del tallo vegetativo para identificar las especies del género en ausencia de estructuras reproductivas.
La areola es una región de crecimiento meristemático, característica de Cactaceae, donde emergen las espinas, además de tricomas, glóquidas, glándulas, flores o ramas (Gibson y Nobel, 1986). Nuestra propuesta parte de una descripción cuantitativa de la areola, como estructura vegetativa perenne que organiza el desarrollo de las espinas, como han sugerido diversos autores (Arroyo-Cosultchi et al., 2010; Baker, 2006; Moore, 1967; Scheinvar et al., 1976) y en el uso de herramientas de estadística multivariada aplicadas con anterioridad (Arroyo-Cosultchi et al., 2010). Estudiamos las especies de Neobuxbaumia y se emplearon las variables de longitud (lineal) y orientación (circular) de las espinas, agrupadas en categorías homólogas, definidas por el orden cronológico ontogenético de aparición (Gibson y Nobel, 1986). La hipótesis de trabajo fue que las espinas y su organización -longitud y orientación en relación con la areola- contienen suficiente información para identificar las 9 especies de Neobuxbaumia reconocidas actualmente. El objetivo de esta investigación fue evaluar la utilidad de la areola y sus espinas para la identificación de las especies de Neobuxbaumia, de manera alternativa ante la carencia de estructuras reproductivas durante la mayor parte del año.
Materiales y métodos
Se realizó un extenso trabajo de campo durante los años 2012 y 2013 para abarcar la distribución del género. Las localidades de estudio se seleccionaron a partir de ejemplares depositados en el Herbario Nacional de México (MEXU) y mediante la consulta de una base de datos especializada (REMIB). Se muestrearon 367 individuos de 36 localidades, pertenecientes a las 9 especies reconocidas de Neobuxbaumia; incluyendo 2 localidades de la especie relacionada Pseudomitrocereus fulviceps, con la finalidad de contrastar los patrones de espinación. El número de localidades por especie varió desde una hasta 7, para representar la amplitud de la distribución geográfica de cada una (apéndice 1).
Los especímenes recolectados se identificaron siguiendo un método taxonómico tradicional, a partir de claves de identificación y de ejemplares de herbario (MEXU, XAL). En cada localidad se recolectó un ejemplar de herbario y cuando fue posible se preservaron flores y frutos en solución fijadora (FAA; Ruzin, 1999) como material de apoyo para la identificación y como referencia; todos los ejemplares preservados se depositaron en MEXU.
Categorización de las espinas
Se tomó como base un esquema de desarrollo ontogenético de la areola, semejante al descrito por Gibson y Nobel (1986) para 2 especies de Echinocactus Link y Otto. Este esquema se adaptó para categorizar las posiciones en las cuales emergen las espinas en las especies de Neobuxbaumia, en función de un criterio de homología. Primero se distinguen las espinas radiales y centrales, considerando la posición relativa en la cual emergen, su forma, textura y coloración. Se reconocen los lados adaxial y abaxial de la areola, con relación al ápice de la planta. La adaxial corresponde a la región floral, mientras la abaxial corresponde a la zona espinífera (Bravo-Hollis, 1978). También se consideró la simetría de la areola, la cual sigue un patrón de desarrollo bilateral. El orden de aparición de las espinas va de la porción abaxial hacia la adaxial de la areola; inicia con una espina radial y posteriormente se van agregando en pares las siguientes espinas, de manera consecutiva (fig. 1). El número de espinas radiales para las especies incluidas en este estudio varía de 1 a 13, siendo más comunes los individuos con 5 o 7 espinas. En cuanto a las espinas centrales, el número más común es de uno, pero pueden existir hasta 3 o 4. En la categoría de espinas centrales solo se consideró a la primera, por ser la única presente en la mayoría de los casos. Un número reducido de individuos (<10% del total) presentó una serie de espinas de crecimiento secundario, identificables por carecer de un orden y por las diferencias en el color, la forma, el tamaño o la consistencia, respecto a las espinas de crecimiento primario. Estos individuos se omitieron del análisis para evitar que influyeran en los estimadores de tamaño y orientación, principalmente en N. euphorbioides donde fueron evidentes.

Figura 1 1) Escala de medición de la dirección de las espinas radiales. 2) Escala de medición de la inclinación de la primera espina central, también se indica la región del tallo donde se realizaron las mediciones. 3) Categorías arbitrarias usadas para organizar los registros de las espinas radiales en la base de datos; la categoría a (espina radial media) se usó como referencia de medición y para establecer comparaciones; el resto de las espinas radiales (b-f y b’-f’) se consideraron como espinas laterales.
Variables descriptoras y bases de datos
En cada localidad se tomaron fotografías de los tallos en 10 individuos, sin indicios de actividad reproductiva, empleando una escala métrica como referencia. A partir de las imágenes digitales se realizó la medición, usando la herramienta de análisis ImageJ ver. 1.48b (Abràmoff, Magalhães y Ram, 2004), integrada en el programa Bio7 ver. 1.7 (Austenfeld, 2012), para obtener simultáneamente las variables de longitud (cm) y orientación (°) para cada espina. Se usó una escala de 0°-360° para las espinas radiales (fig. 1.1) y de 0°-180° para las espinas centrales (fig. 1.2). La asignación de las categorías (posiciones relativas) se realizó manualmente al integrar las mediciones en una matriz de datos agrupados, designando la primera espina radial (categoría media) con la letra a; de manera ascendente y consecutiva, se designaron las siguientes categorías con las letras b - f, y sus simétricas con las letras b’ - f’ (fig. 1.3). En esta matriz de datos cada fila representa una espina y cada registro incluye las variables categóricas (categoría de espina, número de areola, número de individuo, clave de localidad y clave de especie), así como las 2 variables numéricas (longitud y orientación). Esta base de datos está conformada por 21,115 registros, de los cuales 17,973 corresponden a espinas radiales y 3,142 a espinas centrales, los cuales fueron analizados mediante el programa Oriana 4.0 (Kovach Computing Services, 2014) para obtener los estadísticos circulares básicos, como la media, la mediana, la varianza y la desviación estándar, además de proyectar los gráficos circulares (fig. 2). Estos gráficos representan la orientación de cada una de las espinas radiales, clasificadas hasta en 11 categorías predefinidas, así como la orientación promedio para cada categoría ponderada por su longitud promedio.

Figura 2 Gráficos circulares por especie y agrupaciones por similitud visual. Grupo A: 1) Neobuxbaumia sanchezmejoradae, 2) N. multiareolata, 3) N. sp. 1; grupo B: 4) N. mezcalaensis, 5) N. scoparia, 6) N. squamulosa; grupo C: 7) N. macrocephala, 8) N. tetetzo, 9) P. fulviceps; especies independientes: 10) N. euphorbioides, 11) N. polylopha, 12) N. sp. 2.
Análisis multivariados
Los datos agrupados se transformaron para adaptarlos a una matriz rectangular, donde los individuos ocupan las filas (n = 367) y las variables numéricas (longitud, orientación) y categóricas (especie, localidad, individuo) ocupan las columnas. En total se incluyeron 60 variables numéricas divididas en 3 bloques: a) percentil menor (5%), b) mediana y c) percentil mayor (95%). Dichas estimaciones se incluyeron como una medida de la variabilidad de los datos y para acotar el sesgo de los registros extremos. Cada uno de estos 3 bloques se subdivide a su vez en las secciones longitud y orientación, cada una de las cuales está compuesta por 10 variables, 9 corresponden a espinas radiales (categorías e-a-e’; fig. 1.3, tabla 1) y la décima representa la primera espina central. Se aplicó una corrección de los valores en las 3 secciones que contienen variables de orientación, asignando el valor de 360° a las celdas vacías de las categorías b’-f’, en lugar de 0°. La base de datos generada en el paso anterior se visualizó mediante gráficos de variabilidad para identificar las posibles inconsistencias en los datos crudos. Posteriormente, se estandarizaron las variables numéricas de la matriz rectangular. Los estadísticos multivariados básicos se estimaron a partir de la matriz completa para reconocer el agrupamiento de los individuos sin ninguna restricción impuesta. Este análisis también permitió estimar el nivel de correlación entre las variables y la normalidad multivariada del conjunto de datos a partir de la distribución de las distancias de Mahalanobis. El análisis de valores atípicos permitió reconocer los registros con variación extrema; estos registros se descartaron de la matriz en las siguientes pruebas para mejorar el ajuste con la normalidad multivariada y para reducir posibles errores de medición (Dempster, 1971). Los análisis multivariados y las pruebas de significación se obtuvieron mediante XLSTAT® (Addinsoft Inc., 2014) para Microsoft® Excel 2007.
Tabla 1 Caracterización de espinas radiales por especie.
| Especie | # colecta | # esp. | Categorías | CA | GM | [r] |
|---|---|---|---|---|---|---|
| N. euphorbioides | HT001 | 3 | b,a,b’ | 109-246 | 177 | 0.747 |
| HT005 | 3 | b,a,b’ | 135-240 | 184 | 0.778 | |
| HT045 | 3 | b,a,b’ | 113-253 | 178 | 0.700 | |
| Promedio | 3 | b,a,b’ | 130-238 | 181 | 0.751 | |
| N. sanchezmejoradae | HT039 | 5 | c,b,a,b’,c’ | 60-287 | 184 | 0.339 |
| N. macrocephala | HT026 | 11 | f,e,d,c,b,a,b’,c’,d’,e’,f’ | 63-296 | 182 | 0.207 |
| HT028 | 11 | f,e,d,c,b,a,b’,c’,d’,e’,f’ | 57-301 | 188 | 0.160 | |
| Promedio | 11 | f,e,d,c,b,a,b’,c’,d’,e’,f’ | 64-296 | 185 | 0.183 | |
| N. mezcalaensis | HT036 | 7 | d,c,b,a,b’,c’,d’ | 55-309 | 180 | 0.130 |
| HT037 | 7 | d,c,b,a,b’,c’,d’ | 58-308 | 175 | 0.116 | |
| HT042 | 7 | d,c,b,a,b’,c’,d’ | 59-304 | 178 | 0.239 | |
| HT043 | 7 | d,c,b,a,b’,c’,d’ | 43-321 | 176 | 0.147 | |
| HT044 | 7 | d,c,b,a,b’,c’,d’ | 62-300 | 179 | 0.211 | |
| Promedio | 7 | d,c,b,a,b’,c’,d’ | 61-302 | 178 | 0.170 | |
| N. multiareolata | HT034 | 5 | c,b,a,b’,c’ | 28-326 | 210 | 0.011 |
| HT035 | 5 | c,b,a,b’,c’ | 57-305 | 184 | 0.105 | |
| Promedio | 5 | c,b,a,b’,c’ | 50-311 | 182 | 0.087 | |
| N. polylopha | HT007 | 7 | d,c,b,a,b’,c’,d’ | 33-327 | 180 | 0.137 |
| HT008 | 9 | e,d,c,b,a,b’,c’,d’,e’ | 30-334 | 171 | 0.014 | |
| HT009 | 9 | e,d,c,b,a,b’,c’,d’,e’ | 30-329 | 192 | 0.077 | |
| HT025 | 7 | d,c,b,a,b’,c’,d’ | 39-320 | 182 | 0.222 | |
| Promedio | 9 | e,d,c,b,a,b’,c’,d’,e’ | 31-332 | 192 | 0.003 | |
| N. scoparia | HT002 | 5 | c,b,a,b’,c’ | 55-308 | 182 | 0.036 |
| HT003 | 7 | d,c,b,a,b’,c’,d’ | 40-320 | 205 | 0.020 | |
| HT004 | 5 | c,b,a,b’,c’ | 74-286 | 179 | 0.286 | |
| HT010 | 5 | c,b,a,b’,c’ | 56-301 | 177 | 0.143 | |
| Promedio | 5 | c,b,a,b’,c’ | 65-295 | 355 | 0.062 | |
| N. squamulosa | HT006(*) | 5 | c,b,a,b’,c’ | 64-300 | 179 | 0.503 |
| HT011(*) | 3 | b,a,b’ | 112-240 | 177 | 0.685 | |
| HT024(*) | 5 | c,b,a,b’,c’ | 74-280 | 180 | 0.440 | |
| HT030 | 7 | d,c,b,a,b’,c’,d’ | 75-287 | 181 | 0.372 | |
| HT031 | 7 | d,c,b,a,b’,c’,d’ | 68-292 | 182 | 0.294 | |
| HT032 | 7 | d,c,b,a,b’,c’,d’ | 67-300 | 179 | 0.252 | |
| HT033 | 7 | d,c,b,a,b’,c’,d’ | 50-318 | 180 | 0.204 | |
| Promedio | 7 | d,c,b,a,b’,c’,d’ | 71-292 | 179 | 0.277 | |
| N. tetetzo | SNC | 9 | e,d,c,b,a,b’,c’,d’,e’ | 65-300 | 179 | 0.203 |
| SA1413 | 7 | d,c,b,a,b’,c’,d’ | 77-288 | 177 | 0.370 | |
| HT020 | 7 | d,c,b,a,b’,c’,d’ | 75-289 | 180 | 0.375 | |
| HT041 | 9 | e,d,c,b,a,b’,c’,d’,e’ | 59-308 | 178 | 0.210 | |
| Promedio | 9 | e,d,c,b,a,b’,c’,d’,e’ | 65-300 | 178 | 0.209 | |
| N. sp. 1 | HT040 | 5 | c,b,a,b’,c’ | 36-327 | 166 | 0.031 |
| N. sp. 2 | HT038 | 9 | e,d,c,b,a,b’,c’,d’,e’ | 28-335 | 13 | 0.064 |
| P. fulviceps | HT027 | 9 | e,d,c,b,a,b’,c’,d’,e’ | 38-320 | 2 | 0.023 |
| HT029 | 9 | e,d,c,b,a,b’,c’,d’,e’ | 43-320 | 177 | 0.065 | |
| Promedio | 9 | e,d,c,b,a,b’,c’,d’,e’ | 42-318 | 177 | 0.022 |
CA: cobertura angular (o) de espinas radiales, correspondiente al intervalo de confianza de 95%; categorías : categorías correspondientes a las espinas radiales presentes; # colecta : ejemplar de herbario de referencia para cada localidad según Apéndice 1; # esp. : número de espinas radiales; GM : gran media (o); [r] : vector ‘r’; - (*) : especímenes originalmente identificados como N. tetetzo.
Se realizó un análisis canónico discriminante para identificar el subconjunto de caracteres que permite discriminar las 12 clases incluidas en este estudio. Cada especie estuvo representada por un número variable de individuos y localidades, por lo que se ajustaron las probabilidades de presencia de cada una de manera proporcional. Para los fines del presente estudio, se buscó maximizar el poder discriminante reteniendo todas las variables en el análisis, descartando la reducción de la dimensionalidad y la selección de variables. En este caso, se analizó la contribución aditiva del conjunto de variables y de la matriz de correlaciones. Después se aplicó un análisis discriminante clasificatorio (ADC) y una prueba de validación para corroborar la correcta asignación de los individuos dentro de la especie identificada por el análisis canónico discriminante, así como la probabilidad de cada individuo de pertenecer a una especie distinta. Para la prueba de validación, los individuos se distribuyeron aleatoriamente en los grupos de estimación y de validación. El primero se conformó por 287 individuos, los cuales representan el 80% del total de la base de datos, mientras el segundo grupo incluyó 70, que representan el 20% restante. El grupo de validación contó con representantes de cada localidad y de cada especie.
Resultados
Identificación de especies de Neobuxbaumia
ómico empleado para la identificación de individuos a priori permitió corroborar las especies en 31 de las 36 localidades estudiadas. Los individuos de 3 localidades de Oaxaca (HT 006, 011, 024) no correspondieron con la especie identificada previamente (N. tetetzo) y fueron determinados como N. squamulosa a partir de nuestro trabajo (tabla 1; apéndice 1); mientras El método taxonque los individuos pertenecientes a las 2 localidades restantes (HT 038, 040) no se pudieron asignar a alguna de las especies conocidas (tabla 1; apéndice 1). Esta identificación de especies se conservó para los análisis subsecuentes mostrando un alto nivel de congruencia con los patrones de espinación observados, así como con los análisis estadísticos aplicados.
Patrones de espinación
La proyección de los gráficos circulares contribuyó a reconocer 3 conjuntos de especies, considerando el número y la longitud promedio de las espinas radiales, la relación entre la longitud de la espina radial media (a) y las espinas radiales laterales (b-f, b’-f’; tabla 2), la cobertura angular de las espinas radiales (tabla 1), así como la longitud de la primera espina central (tabla 2). El primer conjunto está conformado por N. sanchezmejoradae, N. multiareolata y N. sp. 1, las cuales presentan 5 espinas radiales, con una longitud promedio de 0.7 cm, la espina a puede medir más del doble que las espinas laterales, la cobertura angular abarca de 60 a 300° y la espina central es pequeña o está ausente (grupo A, fig. 2; tabla 1). El siguiente conjunto incluye a N. mezcalaensis, N. squamulosa y N. scoparia, con 5 o 7 espinas radiales, de 0.8 a 1.1 cm de longitud, la espina a es ligeramente más grande que las espinas radiales laterales, la cobertura angular abarca de 50 a 310° (grupo B, fig. 2; tabla 1). El tercer conjunto lo conforman N. tetetzo, N. macrocephala y P. fulviceps, con 9 a 11 espinas radiales, llegando a presentarse ocasionalmente hasta 13 espinas en N. macrocephala, de 1 a 1.2 cm de longitud, la espina a es hasta 1.5 veces mayor que las espinas radiales laterales, la cobertura angular abarca de 40 a 320°; la espina central puede ser de gran tamaño (> 2 cm) (grupo C, fig. 2; tabla 1). Las 3 especies restantes presentan un patrón específico y reconocible con facilidad. N. polylopha mostró la mayor homogeneidad entre poblaciones, presenta de 7 a 9 espinas radiales, las cuales son de mayor longitud que en el resto de las especies (>1.2 cm), de tamaño decreciente a partir de la espina a, siendo considerablemente menores las categorías de la porción apical de la areola (categorías e y e’; tabla 2), la cobertura angular abarca de 31 a 332°; la espina central es relativamente pequeña en comparación con la longitud de las espinas radiales y surge casi perpendicular al eje del tallo (fig. 2.11; tabla 1). N. euphorbioides y N. sp. 2 presentan la menor longitud y homogeneidad en espinas radiales (en promedio 0.3 cm), resultando casi imperceptibles en N. sp. 2 (tabla 2). El patrón particular de N. euphorbioides consiste en presentar un menor número de espinas radiales (de 1 a 3), cortas (∼3 mm), la cobertura angular abarca entre 90 y 270° y la espina central es mayor a las espinas radiales (fig. 2.10; tabla 1). En el caso de N. sp. 2 presenta 9 espinas radiales, cortas (∼3 mm), la cobertura angular abarca de 60 a 300° y la espina central es inconspicua o totalmente ausente (fig. 2.12; tablas 1, 2).
Tabla 2 Longitud promedio (cm) de espina central y espinas radiales. El bloque de espinas radiales está dividido hasta en 11 categorías, de acuerdo con la figura 1.3. Las categorías se agrupan en espinas laterales derechas (b-f), espina media (a), espinas laterales izquierdas (b’-f’) y media aritmética (μ).
| Especie | Espina central | Espinas radiales | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Laterales derechas | Media | laterales izquierdas | μ | ||||||||||
| f’ | e | d | c | b | a | b’ | c’ | d’ | e’ | f’ | |||
| N. euphorbioides | 1.675 | 0.326 | 0.364 | 0.333 | 0.350 | ||||||||
| N. sanchezmejoradae | 0.534 | 0.450 | 0.484 | 1.356 | 0.567 | 0.569 | 0.754 | ||||||
| N. macrocephala | 3.687 | 0.806 | 0.917 | 1.030 | 1.158 | 1.247 | 1.592 | 1.265 | 1.250 | 1.066 | 0.834 | 0.832 | 1.155 |
| N. mezcalaensis | 1.316 | 0.566 | 0.655 | 0.901 | 1.121 | 0.893 | 0.651 | 0.559 | 0.799 | ||||
| N. multiareolata | 0.792 | 0.475 | 0.557 | 1.370 | 0.578 | 0.500 | 0.777 | ||||||
| N. polylopha | 1.408 | 0.519 | 0.451 | 0.863 | 1.729 | 2.436 | 1.805 | 0.861 | 0.457 | 0.538 | 1.200 | ||
| N. scoparia | 1.342 | 0.434 | 0.679 | 0.967 | 0.969 | 0.972 | 0.692 | 0.417 | 0.862 | ||||
| N. squamulosa | 1.363 | 0.732 | 0.842 | 1.168 | 1.594 | 1.110 | 0.790 | 0.700 | 1.142 | ||||
| N. tetetzo | 2.701 | 0.570 | 0.629 | 0.831 | 1.202 | 1.441 | 1.150 | 0.832 | 0.639 | 0.524 | 0.971 | ||
| N. sp. 1 | 0.357 | 0.542 | 1.257 | 0.505 | 0.369 | 0.667 | |||||||
| N. sp. 2 | 0.198 | 0.374 | 0.374 | 0.300 | 0.327 | 0.395 | 0.322 | 0.235 | 0.358 | 0.353 | 0.335 | ||
| P. fulviceps | 4.017 | 0.844 | 0.996 | 1.204 | 1.529 | 1.832 | 1.484 | 1.138 | 0.954 | 0.775 | 1.218 | ||
Análisis canónico discriminante
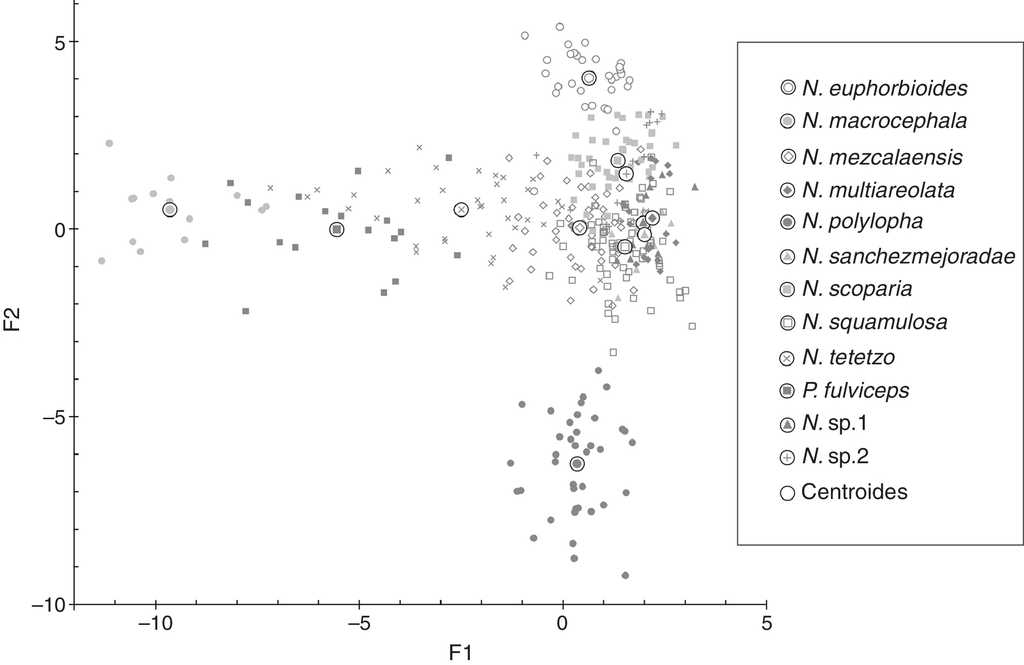
Las primeras 2 funciones canónicas explican 59.2% de la variabilidad total; la función 1 explica 31.6% (valor propio = 7.4024), la función 2 el 27.6% (valor propio = 6.4562), y se requiere un total de 11 funciones para explicar el 100% de la variación. La prueba de lambda de Wilks, que evalúa la existencia de diferencias entre los vectores medios de las clases, indicó una diferencia significativa (p < 0.0001, α = 0.05). Este resultado soporta la existencia de las clases establecidas a priori. La prueba de Fisher (LDA) corroboró que la distancia entre los centroides de las 12 clases fue significativamente diferente, mientras que la prueba unidimensional de igualdad de las medias, indicó la existencia de distintas medias para las clases en la mayoría de las variables (p < 0.0001, α = 0.05), con excepción de una variable que presentó solo una media para todas las clases.
Las variables con mayor contribución en la primera función canónica fueron orientación y longitud de las espinas radiales e y e’, así como la longitud de la espina central (tabla 3). De manera particular, las variables orientación y longitud de las espinas radiales contribuyeron significativamente a la separación de los centroides en la primera función, presentando una gradación de menor a mayor valor desde la espina a. La primera función ubicó a las especies del grupo C con los valores más bajos, separadas de las especies de los grupos B y A, cuyos valores fueron más altos. Por su parte, las variables con mayor contribución en la segunda función canónica fueron la longitud de las espinas radiales en las categorías inferiores (a, b y b’). En este caso las variables también presentaron una gradación, de mayor a menor valor a partir de la espina a. Esta función separó las especies que no pertenecen a ningún grupo; N. polylopha presentó los valores más elevados, mientras N. euphorbioides tuvo los valores más reducidos (tabla 3, fig. 3).
Tabla 3 Coeficientes estandarizados de las funciones discriminantes canónicas. Los caracteres con valores más altos de contribución para cada función se resaltan en negrita.
| Bloque | Variable | F1 | F2 |
|---|---|---|---|
| Orientación | EC | 0.563 | -0.090 |
| ER-e | 0.815 | -0.016 | |
| ER-d | 0.704 | -0.247 | |
| ER-c | 0.535 | -0.396 | |
| ER-b | 0.274 | -0.510 | |
| ER-a | 0.106 | -0.092 | |
| ER-b’ | -0.267 | 0.517 | |
| ER-c’ | -0.525 | 0.394 | |
| ER-d’ | -0.706 | 0.257 | |
| ER-e’ | -0.818 | -0.019 | |
| Longitud | EC | 0.838 | 0.120 |
| ER-e | 0.841 | -0.037 | |
| ER-d | 0.768 | -0.238 | |
| ER-c | 0.591 | -0.321 | |
| ER-b | 0.394 | -0.660 | |
| ER-a | 0.299 | -0.763 | |
| ER-b’ | 0.378 | -0.672 | |
| ER-c’ | 0.598 | -0.333 | |
| ER-d’ | 0.784 | -0.248 | |
| ER-e’ | 0.824 | -0.013 |
EC : espina central; ER : espina radial- categoría (en correspondencia con las claves de la figura 1.3).
Análisis discriminante clasificatorio y prueba de validación
Neobuxbaumia tetetzo y N. sp. 2 se agruparon correctamente en un 86% de los casos, mientras que N. sanchezmejoradae, N. mezcalaensis y N. squamulosa tuvieron el porcentaje de agrupación correcta más bajo (80 a 85%) y se pueden confundir con 2 o 3 clases distintas (tabla 4). En cuanto a la prueba de validación, el grupo de observaciones empleado para estimar la regla discriminante (80% del total) mostró como promedio 92% de individuos correctamente agrupados y presentó una variación entre las clases de 77.8 a 100%. Por su parte, el grupo de validación (20% del total) mostró un desempeño menor, con 78.6% de individuos correctamente asignados en promedio y una variación de 0 a 100% (tabla 5).
Tabla 4 Número de individuos clasificados correctamente por el análisis discriminante clasificatorio para las especies de Neobuxbaumia y Pseudomitrocereus. En negrita se resalta el número de individuos correctamente asignados a su especie.
|
N. euphor- bioides |
N sanchez- mejoradae |
N. macro- cephala |
N. mezca- laensis |
N. multia- reolata |
N. polylopha |
N. scoparia |
N. sp. 1 | N. sp. 2 |
N. squa- mulosa |
N. tetetzo | P. fulviceps | Total | % correcto | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N. euphorbioides | 30 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 33 | 90.91% |
| N. sanchezmejoradae | 0 | 8 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 10 | 80.00% |
| N. macrocephala | 0 | 0 | 14 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 14 | 100.00% |
| N. mezcalaensis | 0 | 0 | 0 | 42 | 0 | 0 | 4 | 0 | 0 | 3 | 1 | 0 | 50 | 84.00% |
| N. multiareolata | 0 | 1 | 0 | 0 | 18 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 20 | 90.00% |
| N. polylopha | 0 | 0 | 0 | 0 | 0 | 38 | 0 | 0 | 0 | 0 | 0 | 0 | 38 | 100.00% |
| N. scoparia | 0 | 0 | 0 | 2 | 0 | 0 | 42 | 0 | 0 | 1 | 0 | 0 | 45 | 93.33% |
| N. sp. 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 12 | 91.67% |
| N. sp. 2 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 13 | 0 | 0 | 0 | 15 | 86.67% |
| N. squamulosa | 2 | 0 | 0 | 5 | 3 | 0 | 0 | 0 | 0 | 56 | 0 | 0 | 66 | 84.85% |
| N. tetetzo | 0 | 0 | 1 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 31 | 0 | 36 | 86.11% |
| P. fulviceps | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 18 | 94.44% |
| Total | 357 | 89.64% |
Tabla 5 Conteo clasificatorio para la regla discriminante (A) y para la prueba de validación (B).
| (A) |
N. euphorbioides |
N. sanchezme- joradae |
N. macrocephala |
N. mezcalaensis |
N. multiareolata |
N. polylopha |
N. scoparia |
N. sp. 1 | N. sp. 2 |
N. squa- mulosa |
N. tetetzo | P. fulviceps | Total | % correcto |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N. euphorbioides | 24 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 26 | 92.31 |
| N. sanchezmejoradae | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 100.00 |
| N. macrocephala | 0 | 0 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 100.00 |
| N. mezcalaensis | 0 | 0 | 0 | 36 | 0 | 0 | 1 | 0 | 0 | 2 | 1 | 0 | 40 | 90.00 |
| N. multiareolata | 0 | 0 | 0 | 0 | 16 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 17 | 94.12 |
| N. polylopha | 0 | 0 | 0 | 0 | 0 | 29 | 0 | 0 | 0 | 0 | 0 | 0 | 29 | 100.00 |
| N. scoparia | 0 | 0 | 0 | 1 | 0 | 0 | 35 | 0 | 0 | 1 | 0 | 0 | 37 | 94.59 |
| N. sp. 1 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 9 | 77.78 |
| N. sp. 2 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 13 | 84.62 |
| N. squamulosa | 1 | 0 | 0 | 2 | 3 | 0 | 0 | 0 | 0 | 49 | 0 | 0 | 55 | 89.09 |
| N. tetetzo | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 27 | 0 | 30 | 90.00 |
| P. fulviceps | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 12 | 13 | 92.31 |
| Total | 287 | 91.99 |
| (B) |
N. euphorbioides |
N. sanchezme- joradae |
N. macrocephala |
N. mezcalaensis |
N. multiareolata |
N. polylopha |
N. scoparia |
N. sp. 1 | N. sp. 2 |
N. squa- mulosa |
N. tetetzo | P. fulviceps | Total | % correcto |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N. euphorbioides | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 100.00 |
| N. sanchezmejoradae | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 2 | 0.00 |
| N. macrocephala | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 4 | 50.00 |
| N. mezcalaensis | 0 | 0 | 0 | 9 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 10 | 90.00 |
| N. multiareolata | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 3 | 66.67 |
| N. polylopha | 0 | 0 | 0 | 0 | 0 | 9 | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 100.00 |
| N. scoparia | 0 | 0 | 0 | 1 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 8 | 87.50 |
| N. sp. 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 3 | 100.00 |
| N. sp. 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 100.00 |
| N. squamulosa | 1 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 11 | 63.64 |
| N. tetetzo | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 1 | 6 | 66.67 |
| P. fulviceps | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 3 | 5 | 60.00 |
| Total | 70 | 78.57 |
Finalmente, en la prueba de validación cruzada se obtuvo un 74.6% de individuos correctamente clasificados y una variación de 46.1 a 89.7% entre clases. En esta prueba, las especies con mayor número de individuos asignados en otras clases (∼50%) fueron N. mezcalaensis, N. multiareolata, N. squamulosa y N. tetetzo debido a la semejanza que guardan con otras especies respecto a su espinación. Particularmente, N. squamulosa y N. mezcalaensis presentan individuos asignados en 3 clases distintas, además de la propia, indicando que estas especies son las más variables del género. Otro conjunto conformado por N. euphorbioides, N. sanchezmejoradae, N. multiareolata, N. scoparia y N. tetetzo presentan individuos asignados en 2 clases distintas; mientras N. sp. 1, N. sp. 2 y P. fulviceps presentan individuos que se asignan en una sola clase distinta (datos no mostrados).
Discusión
Este estudio se centró en las variables de orientación, longitud y número de espinas, así como en la proporción entre las espinas radiales y centrales, para un conjunto amplio de localidades que representan a todas las especies conocidas de Neobuxbaumia y a Pseudomitrocereus fulviceps. Los datos se organizaron para establecer comparaciones únicamente entre espinas homólogas (categorías de espinas; tablas 1, 2). El nivel de correlación entre las variables de orientación fue alto en el conjunto de datos, particularmente entre las categorías opuestas (e.g.: b-b’; fig. 1.3), debido a la simetría bilateral que presenta la areola. Los resultados revelan que la organización de las espinas en la areola es de utilidad en la identificación de las 9 especies de Neobuxbaumia. Incluso, con base en estos análisis proponemos que los individuos encontrados en un par de localidades podrían representar especies aún no descritas.
De acuerdo con nuestros resultados, N. polylopha se identifica por presentar las espinas radiales más largas en el género para las categorías a (> 2.4 cm), b y b’ (> 1.7 cm) (fig. 2.11; tabla 2), y las espinas radiales presentan la cobertura angular más amplia (tabla 1). Estos 2 caracteres, en conjunto con el número de costillas en el tallo (33 ± 0.74) y el tamaño de las areolas (0.26 × 0.19 cm) registrado por Arroyo-Cosultchi et al. (2010) permiten identificarla con certeza. N. macrocephala presenta cierta similitud con N. polylopha, pero sus individuos se distinguen por presentar un mayor número de espinas radiales (11) y la espina central de mayor longitud (3.6 cm; tabla 2). Estos 2 caracteres, en combinación con la forma de la areola (obovada triangular) y el número de costillas (17-26) registrados por Arias et al. (2012) y Bravo-Hollis (1978) son consistentes para distinguir a esta especie.
Neobuxbaumia scoparia y N. euphorbioides también resultaron identificables con atributos de las espinas. La primera se caracteriza por presentar 5 espinas radiales, raramente 7, con una longitud homogénea de 0.8 cm en promedio. Este patrón de espinación se comparte con N. mezcalaensis y N. squamulosa, mostrando una gran similitud en el número y longitud promedio de las espinas radiales y centrales (figs. 2, 3; tabla 2). No obstante, al combinar los 3 atributos de las espinas radiales que encontramos en N. scoparia con otros caracteres vegetativos como la ramificación profusa, el diámetro de sus ramas (8-10 cm), la separación entre areolas (1.2-2 cm) y la presencia de una depresión interareolar marcada (Bravo-Hollis, 1978), se puede diferenciar claramente a esta especie. Por su parte, N. euphorbioides presenta el menor número (3) y la menor longitud (0.3 cm) de sus espinas radiales (tablas 1, 2). El ADC revela la presencia de un porcentaje bajo de individuos asignados a otras especies (tabla 4), los cuales representan el extremo de la variación morfológica de la especie. El patrón de espinación aquí reconocido para N. euphorbioides no se comparte con las otras especies analizadas. Además de los 2 caracteres que permiten discriminar a esta especie (número y tamaño menores de las espinas radiales), existen otros caracteres vegetativos que permiten identificarla de manera fiable, como el crecimiento monopódico del tallo, el número reducido de costillas (6-8), la altura de las costillas (3 cm) y la distancia entre areolas (ca. 1 cm) (Bravo-Hollis, 1978).
Neobuxbaumia sanchezmejoradae se conoce de una sola localidad, donde presenta una baja densidad poblacional, por lo que el número de individuos analizado es menor en relación con otras especies. Además, es el taxón más recientemente descrito del género y no ha sido integrado en ningún estudio comparativo. La mayor similitud en los atributos de la areola fue con N. multiareolata (figs. 2.1, 2.2, 3) por compartir el número de espinas radiales, la proporción de la espina media a que es 2 o más veces más grande que el resto de las espinas radiales y el promedio de longitud de las espinas radiales es de 0.7 cm (tabla 2). Los caracteres que permiten identificar a N. sanchezmejoradae incluyen la cobertura angular intermedia y la proximidad de las espinas b-c y b’-c’ (tabla 1; fig. 1). Otras características morfológicas distintivas de N. sanchezmejoradae son los tallos con ramificación y el número de costillas igual o mayor a 25. Por su parte, N. multiareolata se identifica por una cobertura angular amplia y por una mayor separación entre las espinas b-c y b’-c’ (tabla 1; fig. 2.2). Otras características del tallo que permiten diferenciar a N. multiareolata de la especie anterior, son el número de costillas (≤ 20) y los tallos monopódicos (Arroyo-Cosultchi et al., 2010; Bravo-Hollis, 1978). Esta especie fue segregada de N. mezcalaensis por Bravo-Hollis et al. (1973) y más recientemente Arroyo-Cosultchi et al. (2010) reconocieron que no existen fenotipos intermedios y que los individuos se clasifican en sus respectivas especies. Nuestros resultados apoyan la separación de estos taxa y se registra por primera vez la semejanza de N. multiareolata con N. sanchezmejoradae, considerando únicamente atributos vegetativos.
Neobuxbaumia tetetzo comparte el patrón de espinación con N. macrocephala y P. fulviceps (fig. 2; tabla 1). El ADC indicó que el 13.9% de sus individuos se clasificó incorrectamente en N. macrocephala y N. mezcalaensis debido a que comparte la longitud de la espina central con la primera y el número y cobertura angular con la segunda (tablas 1, 2, 5), pero no se confunde con P. fulviceps en ningún caso. N. tetetzo tiene una distribución amplia en el sur de México, entre los estados de Puebla y Oaxaca, con una marcada variación en el número (7-9) y longitud (0.5-1.4 cm) de sus espinas radiales. Sin embargo, se puede identificar a partir de la longitud promedio de la primera espina central (2.7 cm), además de otros caracteres señalados por Bravo-Hollis (1978) y Arias et al. (2012) como la presencia de podarios, depresión interareolar, número de costillas (13-17), distancia interareolar (7-35 mm), ramificación escasa y tallos de hasta 60 cm de diámetro.
N. squamulosa comparte el patrón de espinación con N. mezcalaensis y N. scoparia (fig. 2), mientras que el ADC revela que el 15.2% de sus individuos se asigna en N. multiareolata, N. mezcalaensis y N. euphorbioides. Los caracteres que permiten identificar a N. squamulosa son el número (3-7) y la longitud promedio de las espinas radiales (1.1 cm), con una cobertura angular intermedia (71° -292°). La amplia distribución geográfica de esta especie, en comparación con otras especies del mismo género (Jalisco a Oaxaca), puede relacionarse con la variabilidad morfológica registrada, la cual podría analizarse en un estudio particular. El atributo con una variación más evidente fue el número de espinas radiales: de 3 a 5 en las localidades meridionales (Oaxaca) y 7 en aquellas más septentrionales (costas de Jalisco, Colima, Michoacán y Guerrero) (tablas 1 y 2). Sin embargo, otros caracteres vegetativos fueron consistentes con datos documentados previamente (Scheinvar y Sánchez-Mejorada, 1990), como el número de costillas (18), el diámetro de las ramas (12-17.5 cm), una ramificación escasa y areolas de 5 × 4 mm con forma obovada. Los individuos de N. squamulosa del istmo de Tehuantepec y Valles Centrales, Oaxaca, fueron asignados previamente como N. tetetzo var. nuda (Bravo-Hollis, 1978) con base en algunos atributos vegetativos -altura de plantas, reducción de espinas-, pero no en el patrón de espinación o en las estructuras reproductivas. Los resultados indican que el patrón de espinación fue consistente entre las localidades de Oaxaca (H. Tapia 006, 011, 024) y las localidades de otros estados (tabla 1; fig. 2.6); del mismo modo, los análisis multivariados indican que nunca se confundieron los individuos de estas localidades con N. tetetzo (fig. 3; tabla 4). N. mezcalaensis presentó 16% de individuos clasificados en N. squamulosa, N. scoparia y N. tetetzo, pero nunca con N. multiareolata, como ya lo habían señalado Arroyo-Cosultchi et al. (2010). En particular, N. mezcalaensis se distingue de N. scoparia por presentar 7 espinas radiales y se distingue de N. squamulosa por una longitud promedio menor de sus espinas radiales. Otros caracteres vegetativos que permiten identificar a N. mezcalaensis son el tallo monopódico, el diámetro del tallo hasta 40 cm, el número de costillas (13 a 25), el margen angulado de sus costillas y la distancia interareolar (18 a 26 mm) (Bravo-Hollis, 1978; Arias et al., 2012).
Neobuxbaumia sp. 1 y N. sp. 2 incluyen individuos de 2 localidades que no pudieron ser asignadas a ninguna de las especies reconocidas. Neobuxbaumia sp. 1 comparte el patrón de espinación con N. sanchezmejoradae y N. multiareolata (fig. 2), cuyo conjunto conforma un conglomerado muy estrecho en el gráfico de las funciones canónicas (fig. 3), siendo su principal diferencia la espina central ausente. Por su parte, N. sp. 2 presenta un patrón de espinación único, considerando el número (9) y la longitud promedio de sus espinas radiales (< 0.3 cm), así como la reducción de la espina central. Otros atributos vegetativos no evaluados cuantitativamente, pero únicos para esta localidad, son la ramificación acrótona, la forma y la profundidad de las costillas, así como la forma y tamaño de la areola. Con base en la información para identificar especies contenida en las espinas y su organización (longitud y orientación), consideramos la hipótesis de que los individuos encontrados en ambas localidades representan especies aún no descritas. Por su parte, Pseudomitrocereus fulviceps comparte el patrón de espinación con N. macrocephala y N. tetetzo. El tamaño de las espinas radiales de P. fulviceps fue ligeramente mayor que en las otras 2 especies (1.2 cm) y la primera espina central tuvo el promedio de longitud más grande de todas las especies analizadas (4 cm). Otros caracteres vegetativos que complementan la identificación de esta especie, incluyen la consistencia flexible de la espina central, el color de las espinas (moreno a amarillento), la forma (circular a obovada) y el tamaño de la areola (1 cm), el color del tallo (verde limón), la ramificación profusa, el número de costillas (11 a 14) y el diámetro del tronco (1 m) (Arias et al., 2012; Bravo-Hollis, 1978).
Los datos de espinación y el uso de análisis multivariados, incluidos los conteos clasificatorios, corroboraron que las especies asignadas a priori son unidades congruentes en todos los casos y que la variación morfológica es mayor entre individuos de diferentes especies que entre individuos de una misma especie procedentes de distintas localidades. Se consideró de alto valor predictivo el empleo de los atributos de las espinas del tallo para una correcta determinación de las especies de Neobuxbaumia y permitieron construir una clave de identificación (apéndice 2). Otros estudios morfométricos apoyan en algún grado esta propuesta (Arroyo-Cosultchi et al., 2010; Baker, 2006; Moore, 1967; Sánchez, Arias y Terrazas, 2013; Scheinvar et al., 1976). Los grupos de especies identificadas a partir de la similitud de sus patrones de espinación (fig. 2) representan hipótesis de homología que podrán someterse a prueba en un eventual análisis filogenético del género.
Se propone la aplicación de estos métodos de análisis en otros géneros de Cactaceae, considerando la dificultad para encontrar estructuras reproductivas durante la mayor parte del año debido a la corta duración de la floración (Valiente-Banuet, Rojas-Martínez, Arizmendi y Dávila, 1997); en muchos casos no es posible observar estructuras reproductivas durante largos periodos, pero sí se cuenta con estructuras persistentes como las areolas y sus espinas. Es evidente que otros caracteres morfológicos, anatómicos y genéticos, podrían sumarse a las variables propuestas en el presente estudio.

