











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
México cuenta con un sistema nacional de áreas decretadas y legisladas para la protección de su biodiversidad, llamadas áreas naturales protegidas (ANP), que forman el Sistema Nacional de Áreas Naturales Protegidas (Sinap-Semarnat, 2001). Sin embargo, estas áreas no necesariamente cumplen con las características básicas para mantener poblaciones viables a largo plazo, ya que como sucede con frecuencia, los criterios para su definición y establecimiento han sido conflictivos y en ocasiones oportunistas (Álvarez y Morrone, 2004; Pressey, Humphries, Margules, Vane-Wright y Williams, 1993; Scott et al., 2001). La selección de áreas para la conservación de la biodiversidad se ha realizado a través de una combinación de métodos tanto ecológicos como biogeográficos (Contreras-Medina y Luna, 2007; Urbina-Cardona y Flores-Villela, 2010; Villaseñor, Ibarra y Ocaña, 1998; Villaseñor, Meave, Ortiz e Ibarra-Manríquez, 2003). Los sistemas de áreas protegidas tanto a escala nacional como estatal o local, son los componentes integrales de la conservación de la biodiversidad. Por desgracia, las decisiones sobre la conservación y manejo de la biodiversidad excluyen con frecuencia análisis detallados para la planificación y diseño de redes de áreas que cumplan con este propósito. En la actualidad es motivo de discusión la necesidad de generar medidas alternativas basadas en estudios minuciosos y factibles, que permitan tener mejores estrategias para la conservación de los recursos naturales a largo plazo (Burgman et al., 2001).
La identificación de una red de áreas para la conservación (RAC) de la biodiversidad eficiente ha sido limitada debido, sobre todo, a la confusión que se genera por la importancia relativa que se ha dado a las diferencias entre el endemismo y la riqueza total de especies (Prendergast, Quinn, Lawton, Eversham y Gibbons, 1993). Existe evidencia de que regiones con alta riqueza de especies no necesariamente son las mismas que aquellas con alto endemismo (Kerr, 1997); la controversia sobre la importancia relativa del endemismo y de la riqueza de especies tal vez no sería necesaria si los criterios de conservación fueran más específicos; por ejemplo, los orientados a algunos taxones en particular (Kerr, 1997).
Los métodos empleados en la selección de áreas de conservación a escala nacional o regional pueden contener mucha subjetividad, en especial debido al escaso conocimiento de la riqueza biológica y su distribución geográfica. Por ejemplo, a escala global, México ha destacado tanto por la rica biodiversidad encontrada en sus ecorregiones (Olson y Dinerstein, 2002) como por sus altos niveles de amenaza, debido a las actividades humanas (Myers, Mittermeier, Mittermeier, Da Fonseca y Kent, 2000). Para la identificación de áreas relevantes para la conservación de la biodiversidad, en los últimos lustros se han empleado métodos iterativos que se basan, sobre todo, en los principios de complementariedad e irremplazabilidad. Dichos métodos emplean un algoritmo muy sencillo, con el que se intenta conservar el mayor número de especies en el menor número de sitios. En México, dicho método ha sido utilizado en diversos estudios (Lira, Villaseñor y Ortiz, 2002; Suárez-Mota y Téllez-Valdés, 2014; Urbina-Cardona y Flores-Villela, 2010; Villaseñor et al., 1998, 2003). Entre estos métodos algunos autores dan mayor importancia a las especies endémicas, y consideran que las especies distribuidas más de forma amplia pueden ser protegidas simultáneamente (Lira et al., 2002; Villaseñor et al., 1998, 2003). Sin embargo, se ha encontrado también que las áreas con alta diversidad no siempre son coincidentes con las de endemismo, por lo que es necesario desarrollar diferentes estrategias de conservación de la biodiversidad en áreas con desigual representatividad o falta de correlación entre las variables utilizadas (endemismo vs. riqueza total) (Arita, Figueroa, Frisch, Rodríguez y Santos-Del Prado, 1997; Ceballos, Rodríguez y Medellín, 1998; Gentry, 1992).
Al utilizar el criterio de complementariedad, la selección de áreas prioritarias para la conservación ha demostrado tener mayor eficiencia. Su aplicación ha generado una gran variedad de algoritmos matemáticos, que en esencia son bastante similares y siguen siempre una serie de pasos iterativos. Por ejemplo, para definir áreas de conservación en México se han empleado métodos de complementariedad basados en algoritmos que minimizan o maximizan ciertas condiciones, como la superficie o la cantidad de especies que protegen (Suárez-Mota y Téllez-Valdés, 2014; Torres-Miranda y Luna-Vega, 2007). La selección de tales áreas se determina mediante un algoritmo que las va seleccionando por procesos iterativos hasta lograr la representación de una o más poblaciones de todas las especies, de la manera más económica posible (Ceballos, 1999; Rodrigues, 1999; Rodrigues, Gregory y Gaston, 2000; Vane-Wright, Humphries y Williams, 1991).
Los métodos iterativos se basan en dos principios: la eficiencia y la eficacia. Por medio de la eficiencia se obtiene el número mínimo de sitios o la menor superficie para albergar y proteger la máxima biodiversidad de una determinada región (Rodrigues, 1999; Williams, 2001). Desde el punto de vista de la eficacia, se selecciona también la superficie mínima o el menor número de sitios que contienen a todas las especies, pero a diferencia de la eficiencia, y en adición a esta, se cumple con objetivos o condiciones específicas. Un objetivo común de estos métodos es, por ejemplo, asegurar la persistencia de toda la biodiversidad posible a largo plazo (Rodrigues, 1999; Rodrigues et al., 2000; Sánchez-Cordero, Peterson y Escalante-Pliego, 2001; Williams, 2001). Estos 2 principios priorizan aspectos diferentes; en el caso de la eficiencia es la mínima superficie, mientras que el de la eficacia es la máxima permanencia de las especies en dicha superficie. La calidad de la información y la determinación de las condiciones en el algoritmo pueden producir resultados intermedios entre ambos principios (Rodrigues et al., 2000). Los métodos iterativos de selección de áreas se basan principalmente en el criterio de complementariedad; sin embargo, en la actualidad los algoritmos pueden ser enriquecidos con procedimientos adicionales diseñados para mejorar su eficiencia (Williams, 2001).
En México, la tasa de deforestación y los cambios en el uso del suelo son considerables (Sánchez-Cordero, Cirelli, Murguía y Sarkar, 2005), por lo que la selección de áreas prioritarias de conservación es primordial en la planificación estratégica de la conservación de la biodiversidad. Las decisiones sobre cómo, cuándo y dónde deben sugerirse áreas de conservación se basan generalmente en expectativas para el futuro. La planificación de la conservación biológica puede incorporar una diversidad de información cuantitativa y cualitativa en el proceso de toma de decisiones actuales (Peterson, 2003).
Durante la última década se han implementado los modelos de nicho ecológico o de distribución potencial de las especies utilizando diferentes algoritmos (Araujo, Pearson, Thuiller y Erhard, 2005; Elith, Graham, Anderson y Group, 2006; Graham, Ron, Santos, Schneider y Moritz, 2004; Guisan y Zimmermann, 2000; Soberón y Peterson, 2004; Thuiller, Lafourcade, Engler y Araujo, 2009). Estos métodos se han ubicado entre los nuevos enfoques emergentes relacionados con la ecología, la biogeografía y la biología de la conservación. Los modelos obtenidos se han considerado buenos sustitutos de la biodiversidad, sobre todo cuando se analizan especies cuya distribución geográfica no es bien conocida (Urbina-Cardona y Flores-Villela, 2010). Consideramos que los modelos de nicho ecológico de especies que se distribuyen en la región del Bajío, en el centro de México, aportan elementos importantes para definir una red de áreas de conservación (RAC) prioritarias. La distribución de las especies estimada mediante sus modelos de nicho ecológico puede evaluarse mediante el uso de un algoritmo de complementariedad (ConsNet) que ha resultado ser eficiente y novedoso (Ciarleglio, Sarkar y Barnes, 2008; Suárez-Mota y Téllez-Valdés, 2014; Urbina-Cardona y Flores-Villela, 2010). Dicha combinación podría fortalecer los argumentos para quienes toman decisiones en el decreto de áreas para la conservación biológica.
Un aspecto importante en biología de la conservación es el número de categorías taxonómicas, que pueden servir como indicador indirecto (sustituto o subrogado) de la riqueza de especies. Por ejemplo, Villaseñor, Ibarra-Manríquez, Meave y Ortiz (2005) señalan que familia es un nivel taxonómico adecuado para su empleo como subrogado y puede ser una herramienta valiosa para localizar y diseñar sistemas representativos de áreas adecuadas para conservar la diversidad de plantas vasculares. Esto resulta especialmente importante en países megadiversos, donde los esfuerzos de conservación han sido obstaculizados por la carencia de inventarios completos de biodiversidad (Contreras-Medina y Luna, 2007; Villaseñor et al., 2005).
La familia Asteraceae ocupa un lugar preponderante en la flora de México tanto en lo que respecta a géneros como a especies, y contribuye sustancialmente a la riqueza florística del país con 362 géneros y más de 3,000 especies (Balleza y Villaseñor, 2002; Villaseñor, 1993; Villaseñor, 2004). Su riqueza se distribuye, además, en todos los tipos de vegetación existentes en el país. Aunque su conocimiento es aún deficiente a escala regional o estatal, la familia es una de las mejor conocidas. Por otra parte, estudios recientes señalan que la familia puede ser un buen subrogado de la riqueza florística total (Villaseñor et al., 2005; Villaseñor, Maeda, Rosell y Ortiz, 2007).
La región del Bajío y zonas adyacentes desde hace varios años está siendo explorada para actualizar el inventario de su riqueza florística (Rzedowski y Calderón, 2003), con más de 150 tratamientos de familias o grupos taxonómicos publicados. Recientemente se concluyó un recuento de la diversidad de especies de la familia Asteraceae presentes en su territorio (Villaseñor y Ortiz, 2012). Sin embargo, la región del Bajío no cuenta con un estudio florístico completo que discuta la relevancia de su diversidad florística ni se han reconocido sitios prioritarios para la conservación de su diversidad. Los sitios de conservación se definen utilizando información de los inventarios taxonómicos; sin embargo, para la mayoría de los taxa se dispone de poca información de su distribución y sus colectas son escasas, por lo que consideramos que una definición de áreas de conservación con base en modelos de nicho ecológico permitirá tener una mejor aproximación a los sitios de mayor riqueza y, en consecuencia, una mejor definición de sitios prioritarios para la conservación de la biodiversidad. Diversos tópicos, biológicos, ecológicos, geológicos, morfotectónicos, económicos o sociales, colocan a la región del Bajío como una zona importante de México. Por lo tanto, el presente trabajo se plantea como objetivo definir una RAC biológica, mediante un análisis de complementariedad, utilizando como subrogado de la diversidad total la información sobre la riqueza florística de Asteraceae que contiene esta porción del territorio mexicano.
La definición de sitios prioritarios de conservación en este trabajo se planteó basándose en el análisis de complementariedad, haciendo uso de algoritmos metaheurísticos implementados en el paquete ConsNet (Ciarleglio et al., 2008; Ciarleglio, Barnes y Sarkar, 2009). Primero, se propuso una RAC basándose en datos observados de la distribución geográfica de especies endémicas de la familia Asteraceae, distribuidas en la región del Bajío; así, se definió una RAC con datos reales (observados). Posteriormente, se definió otra RAC mediante modelos de nicho ecológico que se generaron haciendo uso del programa MaxEnt. Los modelos se emplearon como sustitutos de la biodiversidad para definir sitios importantes para su conservación en la región con datos estimados (o esperados). Por último, se definió una RAC en regiones donde todavía existe vegetación natural y se determinaron zonas irreemplazables por ser sitios donde se distribuyen especies restringidas a su territorio.
Materiales y métodos
Villaseñor y Ortiz (2012) publicaron un recuento de las especies de Asteraceae dentro de la región que comprende el proyecto Flora del Bajío y de Regiones Adyacentes. Los registros que documentan su presencia, junto con las coordenadas geográficas de los sitios de recolecta, sirvieron para elaborar un mapa que muestra su distribución dentro de la zona de estudio (fig 1). El área que delimita la región del Bajío se ubica en el centro de México e incluye a los estados de Querétaro, Guanajuato y la porción noreste de Michoacán (Rzedowski y Calderón, 2003). Su territorio se dividió en 552 celdas de 0.1° de latitud y longitud, tamaño definido siguiendo las recomendaciones propuestas por la UICN (2001; fig. 1) e implementadas por Suárez-Mota y Villaseñor (2011). Posteriormente, se sobrepuso en un modelo digital de elevación (MDE) con resolución de 1 km2, el cual finalmente se utilizó como base para realizar los subsiguientes análisis, tanto de complementariedad como de los aspectos relacionados con los modelos de nicho ecológico de las especies, que se obtuvieron utilizando los datos documentados por Villaseñor y Ortiz (2012) y el programa MaxEnt 3.3.3k. Los modelos resultantes se editaron con el sistema de información geográfica (SIG) ArcMap 10.0. Tomando en cuenta los argumentos discutidos por autores como Peterson et al. (2011), Soberón y Peterson (2005) o Radosavljevic y Anderson (2013), el área para la selección de los sitios de fondo (background ) para generar los modelos se definió con los límites de la zona del Bajío y regiones adyacentes (fig. 1). Las variables climáticas utilizadas para la generación de los modelos se obtuvieron de la base de datos de WorldClim (Hijmans, Cameron, Parra, Jones y Jarvis, 2005; tabla 1).

Figura 1 Sitios de recolecta de las especies de Asteraceae registradas en la región del Bajío y zonas adyacentes. La región de estudio fue dividida en 552 cuadros de 0.1 grados de latitud y longitud.
Tabla 1 Parámetros bioclimáticos utilizados para generar los modelos de nicho ecológico de las especies de Asteraceae de la región del Bajío. Entre paréntesis, las unidades de medida de las variables.
| 1. Temperatura promedio anual (°C). |
| 2. Oscilación diurna de la temperatura (°C). |
| 3. Isotermalidad (°C). |
| 4. Estacionalidad de la temperatura estacional (%). |
| 5. Temperatura máxima del periodo más cálido (°C). |
| 6. Temperatura mínima del periodo más frío (°C). |
| 7. Oscilación anual de la temperatura (°C). |
| 8. Temperatura promedio del trimestre más húmedo (°C). |
| 9. Temperatura promedio del trimestre más seco (°C). |
| 10. Temperatura promedio del trimestre más cálido (°C). |
| 11. Temperatura promedio del trimestre más frío (°C). |
| 12. Precipitación anual (mm). |
| 13. Precipitación del periodo más húmedo (mm). |
| 14. Precipitación del periodo más seco (mm). |
| 15. Estacionalidad de la precipitación (%). |
| 16. Precipitación del trimestre más húmedo (mm). |
| 17. Precipitación del trimestre más seco (mm). |
| 18. Precipitación del trimestre más cálido (mm). |
| 19. Precipitación del trimestre más frío (mm). |
Tomada de Hijmans et al., 2005.
La información sobre los sitios de recolecta que documentan la distribución de las especies en la zona de estudio, junto con la de las variables climáticas (tabla 1), se incorporaron al programa MaxEnt para generar los modelos de nicho ecológico. Para especies que contaron con 25 registros o más se empleó el 75% de ellos como datos de entrenamiento o calibración, y el 25% restante como datos de prueba o validación. Los modelos de las especies que contaron con menos de 25 registros se obtuvieron empleando todos los datos para entrenamiento. Los resultados del análisis del área bajo la curva (AUC) de la ROC parcial se utilizaron para la evaluación de los modelos. Por último, los archivos tipo ASCII generados por MaxEnt fueron importados al SIG para su edición y generación de los modelos binarios (presencia-ausencia).
Los mapas obtenidos con los modelos de nicho ecológico se superpusieron con el mapa de la región del Bajío dividido en celdas de 1 km2 de resolución espacial, similar a la que se encuentran las capas ambientales utilizadas (tabla 1). La coincidencia de esta superposición permitió elaborar una matriz de datos binarios (presencia-ausencia) para llevar a cabo el análisis de complementariedad mediante el uso de los algoritmos metaheurísticos implementados en el software ConsNet (Ciarleglio et al., 2008, 2009), cuyo uso asegura la representatividad de los objetos de conservación definidos como sustitutos (subrogados) o indicadores de la biodiversidad (especies y/o ecosistemas) en ambientes donde todavía existen hábitats naturales. El programa se basa en criterios de configuración espacial (área, forma y conectividad) como parte del proceso de planeación para definir redes de áreas de conservación (Ciarleglio et al. , 2008).
Para definir las RAC, haciendo uso de ConsNet, el área de estudio se divide en celdas (en este caso 0.1°) que deben contener 2 tipos de datos: en primer lugar, información sobre la riqueza biológica que puede incluir especies amenazadas, endémicas, etc., y en segundo lugar, una relación de datos ambientales como los empleados en los modelos de distribución potencial de las especies analizadas (tabla 1). Con este conjunto de datos el programa diseña una RAC, seleccionando al conjunto de celdas que mejor se ajusten a los escenarios de planeación propuestos (Margules y Sarkar, 2007). Los escenarios se definen con base en 3 algoritmos implementados en ConsNet: 1) selección de celdas con los sustitutos con mayor déficit de representación (MDS, por sus siglas en inglés main deficit sampling ); 2) prioridad a la rareza (RF, por sus siglas en inglés rarity first ), donde las celdas con los subrogados más raros son elegidas de forma prioritaria, y 3) un algoritmo que combina los 2 anteriores, denominado ILV (por sus siglas en inglés interleaves ) (Ciarleglio et al., 2009).
Los algoritmos implementados en ConsNet pueden definir RAC con diferentes metas (porcentaje) de conservación, por la presencia obtenida para cada especie, de todas las especies incluidas en el análisis. En este caso, se definieron 2 escenarios: uno considerando el algoritmo MDS con una meta del 10% de la presencia por celda de las especies estimadas con los registros y los modelos de distribución, y otro, considerando el algoritmo RF con una meta del 25% de la presencia de las especies endémicas del Bajío registradas por Villaseñor y Ortiz (2012).
ConsNet es útil cuando se persiguen objetivos encaminados a solucionar problemas espaciales de organización y diseño de áreas de conservación (Ciarleglio et al., 2009). Basándose en la probabilidad de distribución de las especies en cada celda de una retícula determinada, ConsNet toma decisiones de tipo binario, para seleccionar o no una celda sometida a un plan de conservación. Con tales decisiones ordena cada celda jerárquicamente tomando en cuenta su valor de biodiversidad (riqueza de especies). En nuestro análisis definimos como objetivo la reducción del número de celdas que seleccionar, maximizando la contigüidad de la RAC.
El primer escenario de conservación usando el algoritmo MDS, con los datos de los modelos de nicho ecológico en celdas de 1 km2, se realizó con un análisis multicriterio para definir las RAC, tomando en cuenta las coberturas digitales de los tipos de vegetación, las carreteras, municipios y localidades con asentamientos humanos que se encuentran en la región del Bajío (Inegi, 2005). Los tipos de vegetación (Inegi, 2005) se reclasificaron en biomas, siguiendo los criterios propuestos por Villaseñor y Ortiz (2014). Las celdas que, por ejemplo, no contaran con vegetación natural o que incluyeran en mayor medida elementos humanos, como carreteras o asentamientos humanos, fueron excluidas del análisis usando el SIG. Una vez definida la RAC, se obtuvieron los centroides (punto central que cruza las longitudes máxima y mínima) de cada polígono para unirlas mediante una red de tendido mínimo para identificar las mejores rutas de conectividad (mínima distancia) entre los polígonos. El mapa de la RAC obtenido con esta estrategia se sobrepuso con el mapa del Bajío dividido en celdas de 0.1°, para tener un escenario de conservación más homogéneo y con el tamaño de celda y adecuado a este, calculado con los registros de las especies (UICN, 2001).
La RAC obtenida con el algoritmo RF, considerando solamente las celdas (0.1°) donde se han registrado especies endémicas del Bajío, fue jerarquizada utilizando el algoritmo desarrollado por Margules, Nicholls y Pressey (1988) y modificado por Villaseñor et al. (2003). Finalmente, el mapa de las áreas naturales protegidas decretadas en el Bajío se sobrepuso al mapa con la RAC para evaluar correspondencias y diferencias entre los polígonos.
Resultados
La familia Asteraceae registra en la región del Bajío un total de 630 especies (Villaseñor y Ortiz, 2012); de ellas, 330 son endémicas de México y 34 lo son a escala regional; es decir, conocidas solamente de esta región bajo estudio. El Sistema Nacional de Áreas Naturales Protegidas (SINAP, 2014) registra en el Bajío 4 áreas naturales protegidas federales, en cuyo territorio se ha detectado el 61% del total de especies analizadas, el 52% de las especies endémicas de México y el 41% de las especies endémicas del Bajío. De ellas, la Reserva de la Biosfera Sierra Gorda (SINAP, 2014) es la que contiene la mayor diversidad florística dentro de una zona decretada para protección y conservación de la biodiversidad (247 especies de Asteraceae, 12 de las cuales son endémicas de la región del Bajío). Además de esas 4 ANP, en el Bajío se tienen decretadas otras 39 áreas de conservación estatales, en donde se registra el 23.4% de la riqueza total de especies analizadas (tabla 2).
Tabla 2 Riqueza de especies registradas en las ANP decretadas en la región del Bajío. Entre paréntesis el valor porcentual correspondiente.
| Especies | ANP federales (4) | ANP estatales (39) | Todas las ANP (43) |
|---|---|---|---|
| Endémicas de México | 172 (52) | 76 (23) | 205 (62) |
| Endémicas del Bajío | 14 (41) | 00 | 14 (41) |
| Riqueza total | 384 (61) | 145 (23) | 435 (69) |
ANP: áreas naturales protegidas.
Solamente 40.3% de las especies cuentan con 10 o más registros (sitios diferentes de recolecta); otras 266 especies (42.2%) registran 2 a 9 sitios de recolecta y 110 (17.5%) cuentan con un solo registro. Estas 110 especies, aunque no necesariamente se trata de endemismos, representan una rareza en la región del Bajío, pues son especies conocidas hasta la fecha a partir de un solo registro en la zona, si bien se conocen de otras regiones de México. Las celdas donde se distribuyen todas estas especies con un solo registro se seleccionaron como sitios irremplazables para la conservación, pues contienen especies restringidas a ciertas porciones de su territorio. La presencia de las especies ocurre en un gradiente altitudinal de 248 a 3,378 msnm, con el mayor número de sitios de recolecta registrados entre los 2,000 y 2,400 m (fig. 2).

Figura 2 Número de registros de recolecta de Asteraceae a lo largo del gradiente altitudinal (msnm) en la región del Bajío.
La superficie ocupada por la región del Bajío se estima en 60,171.42 km2. De dicha superficie, la ocupada por los biomas (Villaseñor y Ortiz, 2014) que se encuentran en la zona del Bajío (tabla 3) en conjunto cubre 30,205 km2. Dicha cifra indica que 49.8% del territorio del Bajío se ha transformado y/o fragmentado por diversas actividades humanas. El bosque templado, el matorral xerófilo y el bosque tropical estacionalmente seco son los biomas con mayor cobertura y los que también albergan el mayor número de especies analizadas (44.9, 35.9 y 17.2%, respectivamente; tabla 3). Un total de 386 especies (61.3% del total) contaron con el número de registros considerado suficiente (5 registros) para generar modelos de nicho ecológico (MNE). La extensión geográfica registrada por los MNE, más los sitios de recolecta de las otras especies para las que no se generaron modelos, sirvieron para estimar la riqueza de especies en cada una de las celdas en que se dividió la zona de estudio. La división del Bajío en una retícula generó 552 celdas de 0.1° de longitud y latitud (fig. 1). Cada celda a esta escala contiene una superficie aproximada de 100 km2. Se identificaron sobre dicha retícula los sitios de recolecta de las especies, la extensión cubierta por los modelos de nicho ecológico y sus valores de riqueza. De esta manera, la riqueza de especies por celdas varió entre 1 y 144 especies.
Tabla 3 Biomas registrados en la región del Bajío, superficie que abarcan y superficie considerada en las RAC definidas en este estudio.
| Bioma | Superficie (km2) | Especies (N = 630) | ||
|---|---|---|---|---|
| Región del Bajío | RAC(MDS) | RAC(RF) | ||
| Bosque templado | 21,148 | 2,713 | 1,745 | 486 |
| Bosque tropical estacionalmente seco | 8,121 | 1,584 | 255 | 236 |
| Bosque tropical húmedo | 806 | 3 | 25 | 10 |
| Matorral xerófilo | 16,910 | 2,097 | 649 | 232 |
| Superficie con vegetación natural | 46,985 | 6,397 | 2,674 | --- |
MDS: red definida con datos de los modelos de nicho ecológico; RAC: redes de áreas de conservación; RF: red definida con el endemismo encontrado en el Bajío. Adaptado de Inegi, 2005
La estrategia que usó el algoritmo MDS, que tiene como meta de conservación el 10% de las poblaciones de cada especie (suponiendo que cada celda contiene al menos una población de las especies allí registradas o estimadas), seleccionó una red de áreas que cubre una superficie de 8,700 km2 (fig. 3). La RAC generada con este algoritmo consta de 698 polígonos, que varían desde 1 km2 el más pequeño hasta 596 km2 el más amplio. La RAC obtenida incluye 21 de las 110 celdas consideradas como irremplazables (fig. 3). Esta RAC, que incluye la información de los MNE, seleccionó 301 celdas de 0.1° como áreas prioritarias de conservación (fig. 3). En ellas se detectan 6,454 km2 (74.2%) con cobertura vegetal y 2,246 km2 (25.8%) sin vegetación aparente. La red de tendido mínimo mostrada en la figura 4 señala las posibles rutas de conectividad entre los polígonos definidos por la RAC. Los resultados son alentadores para la conservación, pues en dichas áreas seleccionadas se incluyen 590 especies (94.6%) de la riqueza total, 304 (92%) de las especies endémicas de México y 25 (73.5%) de las endémicas del Bajío.

Figura 3 Red de áreas de conservación (RAC) definida con los modelos de nicho ecológico (tono verde) empleando el algoritmo MDS. Los cuadros con tono gris obscuro señalan las celdas de 0.1 grados de latitud y longitud seleccionadas por la RAC. Los polígonos en negro son las áreas naturales protegidas existentes en la región y los puntos negros, los sitios definidos como irremplazables por contener especies exclusivas.

Figura 4 Rutas de conectividad (mínima distancia) entre las áreas prioritarias de conservación en la región del Bajío definidas con la riqueza total y potencial.
El algoritmo RF, que toma en consideración de manera prioritaria la rareza, se utilizó para definir una RAC tomando en cuenta las 34 especies endémicas del Bajío (fig. 5). Dicho algoritmo se utilizó proponiendo como meta la conservación del 25% de la presencia de cada especie. El resultado fue la generación de una RAC formada por 17 polígonos, con los que se podría conservar el 100% de las especies endémicas del Bajío, además de 201 (61%) de las especies endémicas de México y 392 (61%) de la riqueza total de las especies de Asteraceae distribuidas en esta región (tabla 4). La tabla 4 presenta la jerarquización de cada polígono de esta RAC en función de sus valores de riqueza. A cada polígono se le aplicó el nombre del municipio más cercano a su centroide (tabla 5). En el polígono Pinal de Amoles se registraron los valores más altos de endemismo y riqueza florística, por lo que se consideró como el de máxima prioridad; en cambio, el polígono Colón 1 fue el de menor prioridad (fig. 5; tabla 5), pues es la celda que registra el menor número de especies. Las celdas donde se distribuyen especies endémicas de México que no formaron parte de la RAC se seleccionaron como áreas complementarias para, de esta manera, registrar el mayor número de especies endémicas que conservar. Con este complemento sería posible conservar 522 (83%) especies de la riqueza total (630 especies) y 280 (84%) de las 330 especies endémicas de México distribuidas en la región del Bajío (fig. 5).
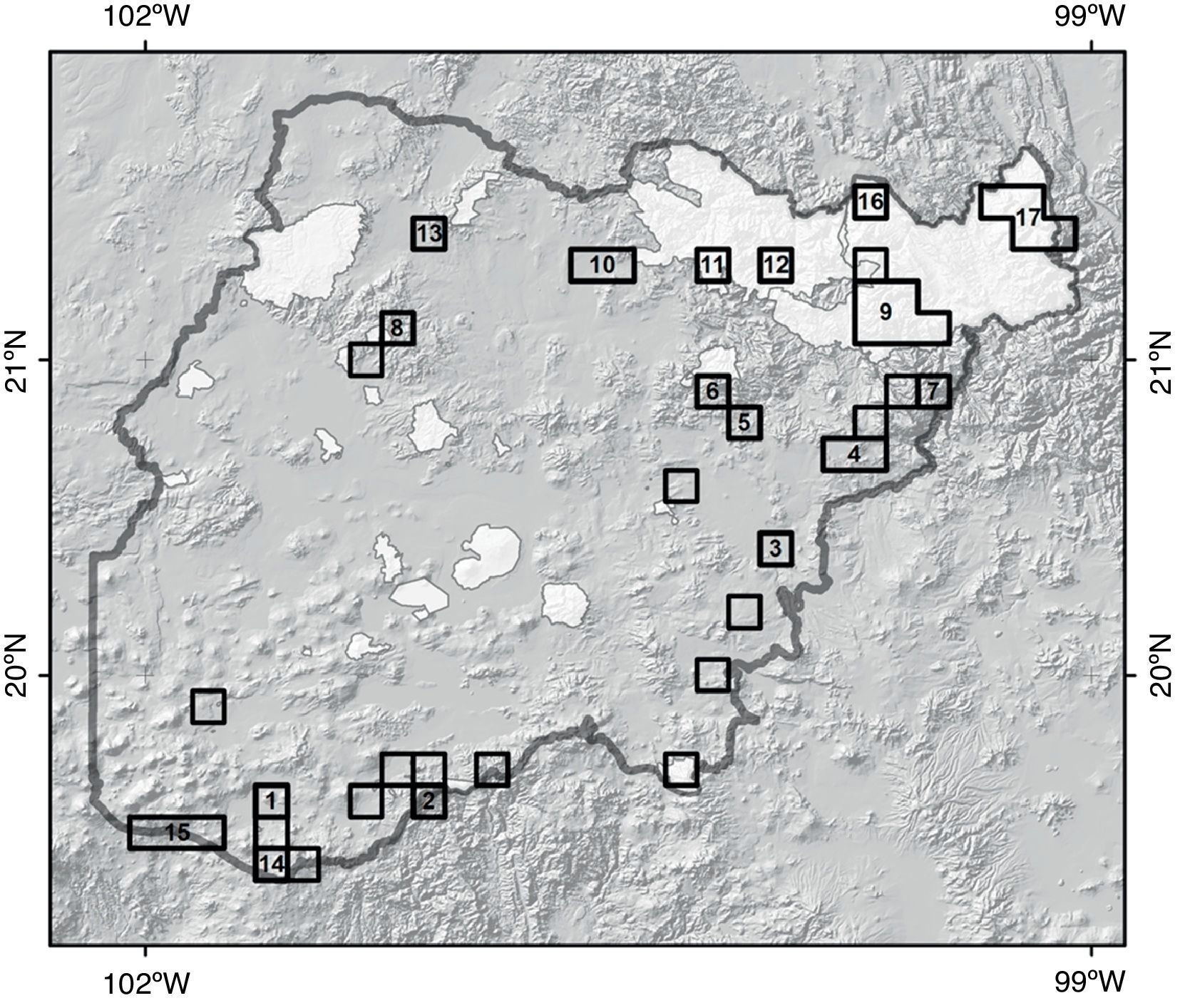
Figura 5 Red de áreas prioritarias de conservación en el Bajío (polígonos numerados), definida con base en las especies endémicas. Los polígonos sin número son áreas complementarias y las zonas claras corresponden a las áreas naturales protegidas.
Tabla 4 Red de áreas de conservación obtenida evaluando tanto las especies de Asteraceae endémicas de México como las endémicas de la región del Bajío.
| Polígono | Nombre del polígono | Riqueza observada (N = 630) | Riqueza estimada (N = 630) | Endémicas de México (N = 330) | Endémicas del Bajío (N = 34) |
|---|---|---|---|---|---|
| 1 | Tzintzuntzan | 45 | 524 | 18 | 1 |
| 2 | Morelia | 72 | 511 | 32 | 3 |
| 3 | San Juan del Río | 20 | 542 | 8 | 1 |
| 4 | Cadereyta de Montes 1 | 60 | 451 | 33 | 2 |
| 5 | Colón 1 | 11 | 449 | 6 | 1 |
| 6 | Colón 2 | 44 | 452 | 24 | 3 |
| 7 | Cadereyta de Montes 2 | 21 | 452 | 11 | 3 |
| 8 | Guanajuato | 78 | 500 | 30 | 1 |
| 9 | Pinal de Amoles | 137 | 459 | 62 | 8 |
| 10 | San Luis de la Paz | 41 | 447 | 18 | 2 |
| 11 | Victoria | 30 | 454 | 14 | 1 |
| 12 | Xichú | 17 | 449 | 10 | 1 |
| 13 | San Felipe | 8 | 414 | 2 | 1 |
| 14 | Salvador Escalante | 33 | 497 | 14 | 1 |
| 15 | Tingambato | 35 | 505 | 15 | 2 |
| 16 | Arroyo Seco | 10 | 441 | 4 | 1 |
| 17 | Landa de Matamoros | 73 | 382 | 27 | 5 |
El número de polígono corresponde a los mostrados en la figura 5; la riqueza observada es el número total conocido de especies, la estimada se obtuvo sumando los modelos de nicho ecológico.
Tabla 5 Prioridad de selección de los polígonos que forman la red de áreas prioritarias de conservación definida con base en el endemismo de la región del Bajío.
| Prioridad | Polígono | Riqueza | Endémicas de México | Endémicas del Bajío | |||
|---|---|---|---|---|---|---|---|
| N = 392 | Complemento | N = 201 | Complemento | N = 34 | Complemento | ||
| 1 | 9 | 137 (34.9) | 255 | 62 (30.5) | 141 | 8 (23.5) | 26 |
| 2 | 2 | 58 (49.7) | 197 | 30 (45.3) | 111 | 3 (32.4) | 23 |
| 3 | 8 | 41 (60.2) | 156 | 21 (55.7) | 90 | 1 (35.3) | 22 |
| 4 | 17 | 33 (68.6) | 123 | 11 (61.1) | 79 | 4 (47.1) | 18 |
| 5 | 4 | 28 (75.8) | 95 | 24 (72.9) | 55 | 2 (52.9) | 16 |
| 6 | 10 | 21 (81.1) | 74 | 9 (77.3) | 46 | 2 (58.8) | 14 |
| 7 | 6 | 20 (86.2) | 54 | 13 (83.7) | 33 | 3 (67.6) | 11 |
| 8 | 1 | 12 (89.3) | 42 | 7 (87.2) | 26 | 1 (70.6) | 10 |
| 9 | 14 | 12 (92.3) | 30 | 7 (90.6) | 19 | 1 (73.5) | 9 |
| 10 | 15 | 8 (94.4) | 22 | 5 (93.1) | 14 | 1 (76.5) | 8 |
| 11 | 7 | 5 (95.67) | 17 | 2 (94.1) | 12 | 2 (82.4) | 6 |
| 12 | 3 | 4 (96.7) | 13 | 3 (95.6) | 9 | 1 (85.3) | 5 |
| 13 | 16 | 4 (97.7) | 9 | 2 (96.6) | 7 | 1 (88.2) | 4 |
| 14 | 11 | 3 (98.5) | 6 | 3 (98.0) | 4 | 1 (91.2) | 3 |
| 15 | 13 | 3 (99.2) | 3 | 2 (99.0) | 2 | 1 (94.1) | 2 |
| 16 | 12 | 2 (99.7) | 2 | 1 (99.5) | 1 | 1 (97.1) | 1 |
| 17 | 5 | 1 (100) | 0 | 1 (100) | 0 | 1 (100) | 0 |
Entre paréntesis se indica el porcentaje de frecuencia acumulada. Los números de los polígonos se explican en la tabla 4.
Discusión
Los resultados permiten identificar áreas prioritarias para la conservación en el Bajío y las zonas adyacentes, usando como subrogado de la biodiversidad a la familia Asteraceae. Dicha familia ha demostrado una correlación significativa entre sus miembros con las demás especies de la flora de México (Villaseñor et al., 2007). Dicha correlación permite conjeturar que la RAC obtenida confirmará su eficiencia en la conservación de otros miembros de la riqueza florística de la región, cuando se tengan inventarios de tales sitios. Por esto se propone como prioridad llevar a cabo los censos florísticos de varios de esos polígonos, con el propósito de confirmar la veracidad de estas conjeturas. Esta RAC permitirá también en un futuro identificar y seleccionar, con bases conceptuales más sólidas, áreas que, al integrarse al SINAP, podrán coadyuvar a mitigar los efectos de la degradación de la cobertura vegetal tan pronunciada en la región bajo estudio.
Los ejercicios de identificación de redes de áreas de conservación utilizando el algoritmo MDS de ConsNet derivaron en la selección de 301 celdas (de un total de 552) como prioritarias para la conservación, una extensión territorial (8,700 km2) no viable para propósitos de conservación. Sin embargo, se identificaron algunas estrategias, como por ejemplo las derivadas del algoritmo RF, que considera áreas más adecuadas para ser consideradas para propósitos de conservación de la riqueza florística de la región bajo estudio (tablas 4 y 5). En conjunto, se identifican como relevantes para la conservación 17 polígonos, que incluyen en total 28 celdas, lo que supone una superficie de aproximadamente 2,800 km2. La superficie que abarca y la riqueza de especies que incluye son más adecuadas para considerarse en futuras estrategias de conservación (fig. 5).
La implementación de una RAC tomando en cuenta los sitios irremplazables, aquellos que registran 110 especies conocidas solamente en esa porción de territorio, obtenida con el algoritmo MDS es una opción importante que debe considerarse. Además de seleccionar sitios para la conservación del endemismo estricto y exclusivo de la región del Bajío, permitiría coadyuvar a la protección de poco más de la mitad de las especies nativas y/o endémicas de México registradas en la zona de estudio. Estos sitios, además, contienen importantes zonas con vegetación natural y todavía con menor impacto antropogénico (tabla 5). Por otra parte, su exclusión dentro del SINAP provocará sin lugar a dudas la extinción del endemismo local y de poblaciones regionales de especies endémicas de México, así como la disminución del acervo genético de otras especies conviviendo con este endemismo en la región. Indudablemente los 2 componentes endémicos, nacional y local, requieren de estrategias de conservación más específicas (Margules, Pressey y Williams, 2002; Margules y Sarkar, 2007), y una primera aproximación es la identificación de áreas donde pueden ubicarse y monitorearse sus poblaciones, como las especificadas en este trabajo, además de aquellas otras que han sido reconocidas en trabajos previos usando otros grupos florísticos (Hernández y Bárcenas, 1996; Hernández-Magaña, Hernández-Oria y Chávez, 2012; Rzedowski, Calderón de Rzedowski y Zamudio, 2012; Sánchez-Martínez, Chávez-Martínez, Hernández-Oria y Hernández-Martínez, 2006).
Los resultados obtenidos tomaron en consideración, como factor importante, los sitios con vegetación natural remanente en la región del Bajío (Inegi, 2005). Por desgracia, la cartografía utilizada puede ser algo obsoleta en la actualidad y la región puede haber sufrido importantes modificaciones debido a la acelerada pérdida de vegetación natural ocasionada por las actividades humanas. Será importante la prospección in situ de las zonas seleccionadas para verificar de forma fehaciente su relevancia actual como sitios importantes para la conservación. Las áreas de conservación definidas en nuestro análisis muestran coincidencias con las propuestas para especies de otros grupos, como los señalados por Hernández y Bárcenas (1996), Hernández-Magaña et al. (2012) o Rzedowski et al. (2012), lo que sugiere que la familia Asteraceae es un buen subrogado de la biodiversidad, con valor predictivo para otros componentes de la flora como se señaló previamente (Villaseñor et al., 2005; 2007).
La distribución de las especies analizadas entre los biomas identificados en la región del Bajío es heterogénea (tabla 3). Dominan por su riqueza de especies los bosques templados, seguramente porque ocupan una mayor extensión en la región. Los bosques tropicales estacionalmente secos y los matorrales xerófilos, aunque difieren notablemente en extensión (tabla 3), registran valores de riqueza más o menos equivalentes en la región del Bajío. Las coberturas de los biomas en las RAC definidas comprenden áreas contrastantes en sus dimensiones. Por ejemplo, la cobertura vegetal en la RAC definida con el endemismo (algoritmo RF) es menor, por ser la estrategia que seleccionó el menor número de celdas. El bosque templado predomina en todos los escenarios, lo que sugiere que la mayoría de las especies tienen una mayor afinidad por los ambientes templados.
La combinación del conocimiento derivado de los modelos de nicho ecológico o distribución potencial y los algoritmos para la selección de áreas prioritarias para la conservación permiten identificar zonas para orientar o establecer mejores estrategias de conservación. En este trabajo, el escenario de conservación mediante dichos modelos permitió localizar zonas del Bajío adicionales a los sitios conocidos con uno o pocos registros. Dicha estrategia permite llenar los vacíos que el esfuerzo de recolecta no ha podido cubrir, aunque siempre será necesaria su validación con trabajo de campo adicional, para verificar fehacientemente la presencia de dichas especies en las zonas propuestas donde se estima su presencia. Lo importante es que ejercicios como este permiten ubicar con mayor precisión los lugares con mayor relevancia para llevar a cabo dicho trabajo de campo.
Se han llevado a cabo estudios basados en el principio de complementariedad en el Eje Volcánico Transversal, proponiendo inclusive corredores biológicos (Sánchez-Cordero et al. , 2005; Suárez-Mota y Téllez-Valdés, 2014). La mayor parte de la región del Bajío se ubica en la porción septentrional de dicha cadena montañosa, por lo que sería importante evaluar la coincidencia geográfica entre dichos trabajos. Tal coincidencia permitiría reforzar la selección de sitios prioritarios para la conservación de la biodiversidad de la región. Con ello, se podría complementar el SINAP con zonas protegidas seleccionadas con criterios más rigurosos que los criterios ad hoc utilizados en la actualidad en muchas de las áreas de protección decretadas (Peterson, Egbert, Sánchez-Cordero y Price, 2000). A la luz de los resultados aquí presentados, es indudable que en la región del Bajío se debe incrementar la superficie de las áreas naturales protegidas ya existentes. Las celdas contiguas a dichas áreas protegidas, identificadas en este ejercicio (figs. 3, 5), podrían ser un recurso que coadyuve con dichas estrategias de conservación de la riqueza florística. Por ejemplo, la Reserva de la Biosfera Sierra Gorda, que abarca la sierra Gorda de Guanajuato y Querétaro, se ha definido en este estudio como un "corredor" que une sitios con alta diversidad biológica en el Bajío (fig. 4). En México, muchas de las áreas naturales protegidas con decreto carecen de información acerca de la composición de sus atributos, tanto biológicos como ambientales. Análisis como el aquí presentado destacan la poca coincidencia entre las áreas identificadas como importantes para la conservación (RAC) con las áreas naturales protegidas consideradas por el SINAP. Los resultados igualmente sugieren que el uso de subrogados, ya sean grupos taxonómicos o características ambientales, permiten identificar sitios adicionales que contribuirían a una redefinición de estas áreas, lo cual redundaría en mejores y más eficientes planes de conservación de la biodiversidad no solo en el Bajío, sino también en todo el territorio nacional. Los escenarios que se obtuvieron en este trabajo son ejemplos de que empleando métodos metaheurísticos se pueden definir RAC más eficientes, pues consideran posibles estrategias de conservación en superficies más pequeñas que las consideradas actualmente por el SINAP, incluyendo, además, una mayor riqueza florística y porcentajes más importantes de elementos raros, como son sus especies endémicas locales (tabla 5). Por ejemplo, la Reserva de la Biosfera Sierra Gorda es la zona protegida en la región del Bajío que registra el mayor número de especies endémicas locales (12). Las RAC obtenidas en este trabajo pueden ofrecer alternativas para la selección de sitios adicionales de conservación de la biodiversidad con menor costo, comparadas con las estrategias de conservación convencionales actualmente establecidas.
En la actualidad, las bases de datos disponibles y los sistemas de información geográfica facilitan la recopilación y la cartografía de mucha información biológica (Margules y Austin, 1994). Esta información, analizada con algoritmos como los implementados en ConsNet, permite seleccionar de manera jerárquica áreas de conservación biológica, como la que aquí se presenta, ligadas a posibles rutas de conectividad entre ellas (fig. 4). De esta manera, los tomadores de decisiones, encargados de seleccionar sitios para la conservación, tienen argumentos adicionales con mejores fundamentos ecológicos y biogeográficos. Por último, los resultados de este trabajo pueden complementar las estrategias para la conservación de la vegetación de México planteada por la Conabio (2012).
Agradecimientos
El primer autor agradece al Conacyt la beca postdoctoral (163037) asociada al Posgrado en Botánica del Colegio de Postgraduados campus Montecillos. La base de datos de Asteraceae analizada en este estudio fue generada en parte con apoyo económico de la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad y del Instituto de Biología de la UNAM (Programa SIBA). Se agradece la asistencia técnica proporcionada por Enrique Ortiz, así como a los 2 revisores anónimos por sus comentarios y sugerencias.

