











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Las actividades humanas han modificado de forma diferencial el ambiente original. En sus inicios, las actividades agrícolas dieron pauta a esos cambios al transformar ambientes naturales en zonas de cultivo. Actualmente las evidencias más conspicuas de estas actividades se detectan en la erosión y la contaminación del suelo y de mantos freáticos debido, en gran parte, al desmonte y a la acumulación de grandes cantidades de desechos, crecimiento urbano irregular en ambientes de riesgo ecológico, entre otros (Cochrane y Barber, 2009; Haberl et al., 2001). A nivel forestal, los cambios en la estructura de la vegetación han favorecido la pérdida de la biota (Santos y Tellería, 2006; Saunders, Hobbs y Margules, 1991). Aunado a esto, el desarrollo urbano tiene efectos irreversibles implicando la alteración y la desaparición del hábitat, cambios geográficos y biológicos en la región, tales como flujo del agua, degradación y contaminación del suelo, además de crecimiento poblacional y actividades industriales (Pisanty, Mazari y Ezcurra, 2009).
En este contexto, los insectos juegan un papel importante para evaluar los efectos que estos cambios generan en la composición y en la estructura de la diversidad local. Samways, McGeoch y New (2010) puntualizan la existencia de estudios que permiten estimar el efecto del cambio climático, la contaminación, el cambio de hábitat, la fragmentación, los efectos causados por las actividades antrópicas, a través del uso de grupos de insectos como indicadores, por ejemplo: Odonata, Hymenoptera y Coleoptera. A partir de un análisis detallado para la elección de grupos indicadores, Favila y Halffter (1997) propusieron a los escarabajos del estiércol de la subfamilia Scarabaeinae (Scarabaeidae), debido principalmente a que es un grupo relativamente bien conocido tanto en su taxonomía como en su historia natural, ya que son de fácil monitoreo, además de ser sensibles al cambio en el uso de suelo o a la fragmentación forestal.
Las larvas y adultos de Coleoptera se pueden encontrar en hábitats diversos; en ellos utilizan una gran variedad de recursos como alimento: fruta, semillas, polen, raíces, hongos, otros insectos, materia orgánica en descomposición tanto de origen animal como vegetal (Lawrence y Britton, 1991; Martínez, Cruz, Montes de Oca y Suárez, 2011; Morón, 2004; Navarrete-Heredia, 2009; Navarrete-Heredia, Delgado y Fierros-López, 2001). De manera particular, aquellos que se encuentran asociados a restos de animales han sido reconocidos como coleópteros necrócolos (Labrador-Chávez, 2005; Naranjo-López y Navarrete-Heredia, 2011). Estos últimos son importantes en los ecosistemas, debido en primera instancia a que remueven los cadáveres del suelo, ayudan a reciclar nutrientes (Quiroz-Rocha, Navarrete-Heredia y Martínez-Rodríguez, 2008), reducen la población de fauna nociva (Bishop, McKenzie, Spohr y Barchia, 2005), tienen relevancia en la medicina forense, junto con el orden Diptera, para estimar el tiempo post mortem (Castillo-Miralbés, 2001; Gennard, 2007), y son importantes como grupos indicadores de biodiversidad (Favila y Halffter, 1997; Yanes-Gómez y Morón, 2010). En México se han registrado varias familias de coleópteros necrócolos, siendo Carabidae, Histeridae, Leiodidae, Silphidae, Staphylinidae, Dermestidae, Scarabaeidae, Trogidae y Cleridae las familias más frecuentes y abundantes, aunque por su biomasa destacan Silphidae y Scarabaeidae (Deloya, Ruiz-Lizárraga y Morón, 1987; Morón, Deloya y Delgado-Castillo, 1988; Naranjo-López y Navarrete-Heredia, 2011, entre otros). Con base en lo anterior, el objetivo del presente estudio fue determinar la diversidad y la estructura del ensamble de coleópteros necrócolos (Scarabaeidae, Trogidae y Silphidae) en un bosque urbano (Bosque Los Colomos) de la ciudad de Guadalajara, Jalisco, México.
Materiales y métodos
El Bosque Los Colomos (BLC) es una Área Natural Protegida bajo la categoría de Área Municipal de Protección Hidrológica, cuyo objetivo es proteger los mantos acuíferos, conservar y restaurar los ambientes naturales, así como realizar actividades de recreación (Cordero, 2009). Se ubica en el municipio de Guadalajara, a 1,556 m snm entre las coordenadas 20°42'38" N, 103°24'12" O y 20°42'07" N, 103°23'26" O. Limita al norte con el municipio de Zapopan (fig. 1). Se reconocen asociaciones vegetales de pino (Pinus ayacahuite , P. devoniana , P. douglasiana , P. maximartinezii , P. oocarpa , P. patula , P. tenuifolia ), vegetación secundaria, además de asociaciones con especies introducidas: casuarina (Casuarina cunnighamiana , C. equisetofolia ) y eucalipto (Eucalyptus camaldulensis , E. citriodora , E. globulus , E. robusta ) (Jara-Arce y Orendain-Díaz, 2009). Presenta un clima templado semicálido [(A)C(w1)(w)] con lluvia en verano, con una precipitación anual de 976.5 mm y una temperatura media anual de 19.5 °C (Loza-Ramírez y González-Salazar, 2009). El BLC está dividido en 2 secciones: Colomos-I (CI) y Colomos-II (CII). Colomos-I cuenta con atractivos inmuebles públicos, elementos arquitectónicos utilizados como oficinas, área de asadores, lago artificial, pistas de trote pavimentadas, canchas de baloncesto, quiosco, área de juegos infantiles, entre otros servicios públicos; en esta sección la vegetación está reducida y distribuida en ciertas áreas. Colomos-II presenta vegetación arbustiva y arbórea, con un área recreativa menor y pistas sin pavimentar. Considerando la densidad de servicios públicos (criterio cualitativo), se asumió que CI tiene mayores actividades recreativas y, en consecuencia, una menor densidad de bosque conservado.
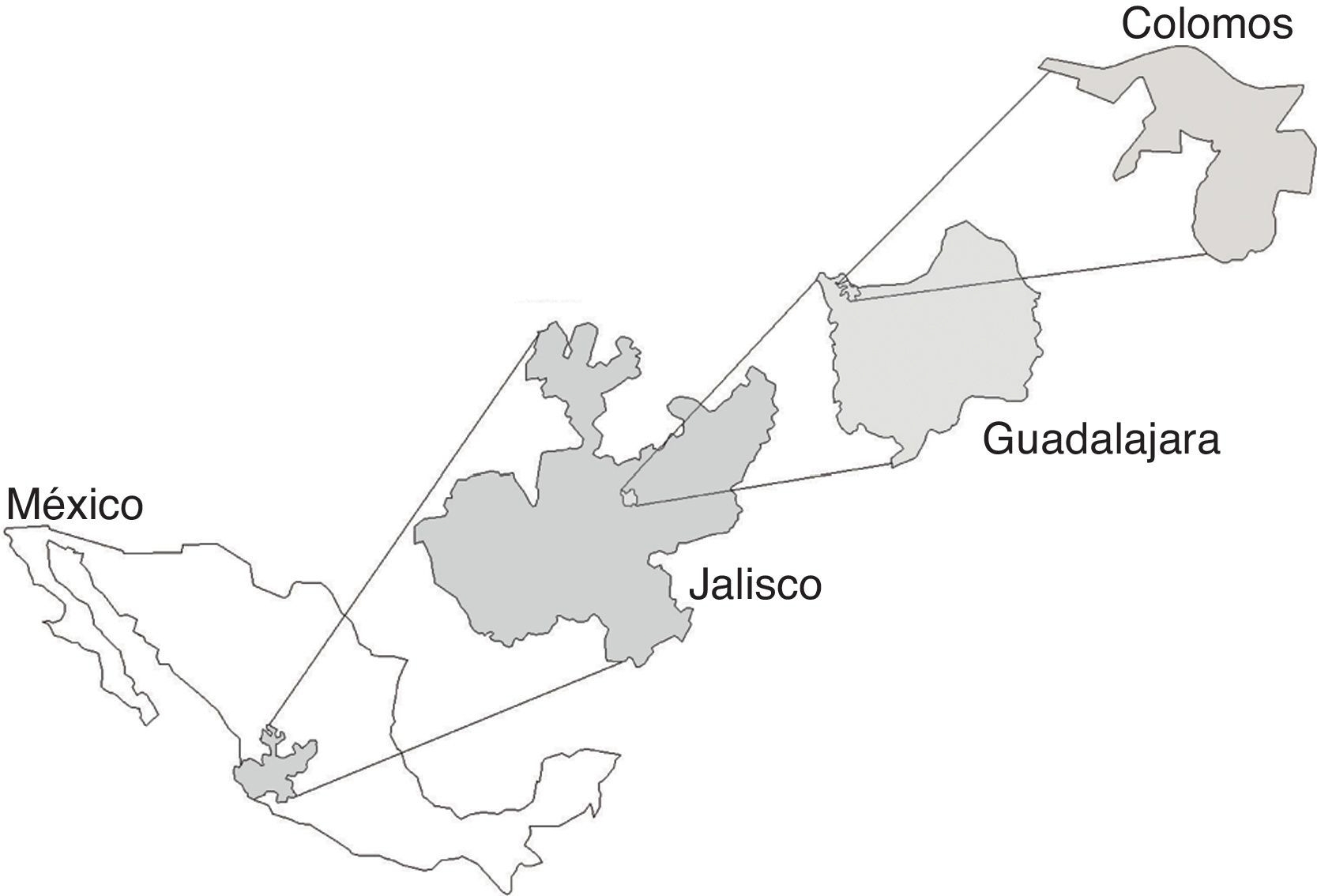
Figura 1 Área de estudio Bosque Los Colomos, Guadalajara, Jalisco, México (adaptada del anexo cartográfico Bosque Los Colomos, 2010: elaborada por L. A. Navarrete-Quiroz).
El muestreo se realizó entre julio de 2011 y julio de 2012. En cada una de las 2 secciones (CI y CII) se eligieron 3 sitios, cada uno con diferente tipo de vegetación: pino, casuarina y eucalipto. Se utilizan números romanos para asociar al tipo de vegetación con una sección particular del bosque; por ejemplo, pino-I indica el sitio ubicado en la zona de pino de la sección I del bosque. Se eligió, además, otro sitio en una zona de transición entre las 2 secciones, representado con vegetación secundaria (fig. 2). En cada sitio se colocaron 2 necrotrampas permanentes del tipo NTP-80 (Morón y Terrón, 1984) con alcohol al 70% como líquido fijador y cebadas con calamar fresco. El cebo se reemplazaba cada mes y las muestras obtenidas se conservaron en alcohol al 70%. El material recolectado se lavó, separó, montó y etiquetó. Las determinaciones a nivel específico se basaron en los criterios de Howden y Cartwright (1963) y Morón et al. (1988) para Scarabaeinae, Navarrete-Heredia (2009) para Silphidae, así como Vaurie (1955) y Deloya (2005) para Trogidae. Las determinaciones se corroboraron con ejemplares de la colección del Centro de Estudios en Zoología, Universidad de Guadalajara (CZUG, Zapopan, Jalisco) y de la colección entomológica del Instituto de Ecología (IEXA, Xalapa, Veracruz). Los ejemplares se encuentran depositados en ambas colecciones.
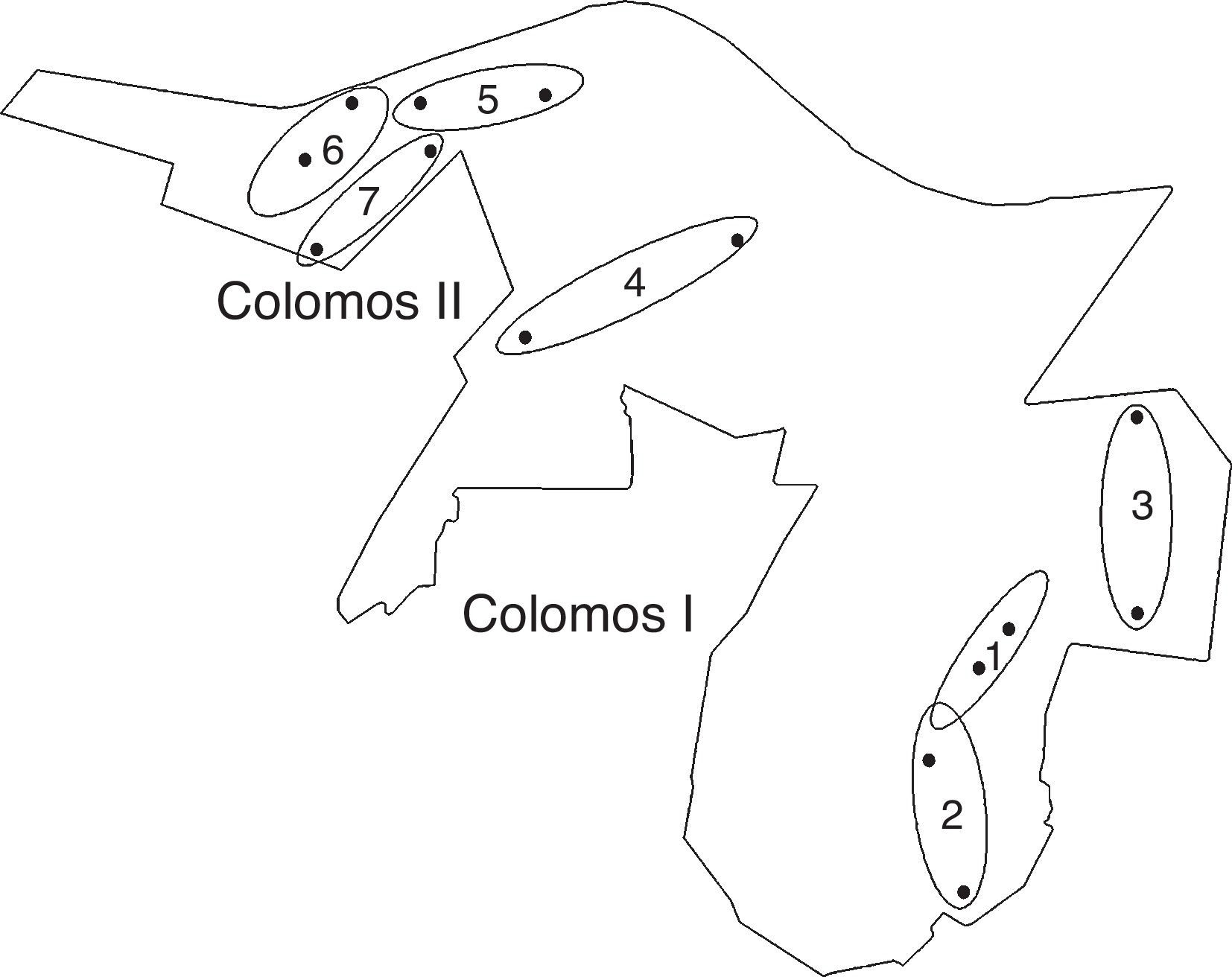
Figura 2 Ubicación de los sitios en el Bosque Los Colomos, Guadalajara, Jalisco, México. Casuarina (1 y 7), pino (2 y 5), eucalipto (3 y 6), vegetación secundaria (4) (adaptada del anexo cartográfico Bosque Los Colomos, 2010: elaborada por L. A. Navarrete-Quiroz).
Los datos de abundancia/especie se analizaron con base en los criterios propuestos por Colwell (2009) y Moreno, Barragán, Pineda y Pavón (2011). Se registró el número de especies (S ) y la abundancia (n ) para cada una ellas, para cada sitio y mes. Para cada sitio se elaboraron curvas de "rango-abundancia". La diversidad se evaluó con números efectivos (medidas de "diversidad verdadero sensu"; Jost, 2006) siguiendo los criterios de Moreno et al. (2011). Para la diversidad estimada de orden 0 (0D = riqueza de especies) se utilizó el estimador no paramétrico ACE (Chao y Lee, 1992); para la diversidad de orden 1 (1D = exponencial del índice de Shannon) se utilizó el estimador propuesto por Chao y Shen (2003)(Bias-corrected Shannon diversity estimator) . Los cálculos se realizaron en el programa SPADE (Chao y Shen, 2010). Para determinar el grado de similitud entre los sitios se utilizó el coeficiente de Bray-Curtis (Group-Average Clustering) con ayuda del programa Biodiversity Pro (Hammer, Harper y Ryan, 2001). Para evaluar la relación entre la abundancia y los valores de precipitación y temperatura durante los meses de muestreo se realizó un análisis de correlación canónica (ACC). Se utilizó el programa Statistica versión 6.0 (Statsoft, 1995).
Resultados
Se recolectaron 362 ejemplares pertenecientes a las familias Trogidae, Silphidae y Scarabaeidae (Scarabaeinae) que representan a 8 especies: Trox spinulosus dentibius Robinson, 1940, Omorgus rubricans Robinson, 1946, y Omorgus suberosus Fabricius, 1775 (Trogidae); Nicrophorus olidus Matthews, 1888, y Oxelytrum discicolle (Brullé, 1840) (Silphidae) Dichotomius amplicollis (Harold, 1869), Onthophagus lecontei Harold, 1871, y Onthophagus batesi (Howden, 1955) (Scarabaeidae: Scarabaeinae).
Trox spinulosus dentibius (Trogidae) fue la especie con mayor abundancia porcentual relativa (48.3%), seguida por O. discicolle (Silphidae) (28.7%); en cambio, O. batesi obtuvo la menor abundancia (1.3%). Dentro de las secciones del bosque, la distribución y abundancia de las especies fue variable, mostrando diferencias significativas entre CI y CII (χ2 = 58.80, p = 2.6127E-10, gl = 7). La mayor riqueza específica y abundancia se observó en los sitios de CII, donde el bosque de eucalipto-II presentó los valores mayores, con las 8 especies y 97 ejemplares. En el bosque de pino-I se presentó la menor riqueza específica (2 especies) y abundancia (25 ejemplares). Considerando a las 2 especies más abundantes, O. discicolle predominó en CI y T. spinulosus dentibius en CII (tabla 1).
Tabla 1 Macrocoleópteros necrócolos del Bosque Los Colomos, Guadalajara, Jalisco, México (expresado por sitio y número de ejemplares).
| Especies | Colomos I | Colomos II | Vegetación secundaria | Total | ||||
|---|---|---|---|---|---|---|---|---|
| P | C | E | P | C | E | |||
| Trox spinulosus dentibius | 3 | 5 | 5 | 28 | 55 | 36 | 43 | 175 |
| Omorgus rubricans | 0 | 0 | 3 | 10 | 1 | 6 | 0 | 20 |
| O. suberosus | 0 | 0 | 0 | 0 | 0 | 1 | 8 | 9 |
| Nicrophorus olidus | 0 | 5 | 1 | 1 | 0 | 9 | 0 | 16 |
| Oxelytrum discicolle | 22 | 16 | 6 | 7 | 4 | 34 | 15 | 104 |
| Dichotomius amplicollis | 0 | 0 | 0 | 6 | 3 | 6 | 8 | 23 |
| Onthophagus lecontei | 0 | 0 | 6 | 1 | 0 | 3 | 0 | 10 |
| O. batesi | 0 | 0 | 2 | 0 | 0 | 2 | 1 | 5 |
| Total | 25 | 26 | 23 | 53 | 63 | 97 | 75 | 362 |
C: casuarina; E: eucalipto; P: pino.
Se observaron 2 picos de abundancia: i) en época de lluvias (julio-octubre), donde las especies con mayor abundancia corresponden a Trogidae y Scarabaeinae y ii) en los meses más fríos (noviembre-febrero), siendo dominantes las especies de Silphidae. Durante el periodo de julio-noviembre se presentó la mayor abundancia (85%), correspondiendo a septiembre el valor más elevado (24.0%). Trox spinulosus dentibius presentó su mayor abundancia durante julio y septiembre, mientras que O. discicolle en noviembre (fig. 3). La mayor riqueza específica se observó entre junio-septiembre (S = 8), y la menor riqueza entre enero y abril (S = 2) (fig. 3). Con base en el ACC, el valor global de 0.96 (p < 0.05) indica que las variables ambientales: temperatura y precipitación mensual, se relacionan con las abundancias de las especies de escarabajos, siendo más evidente en las especies más abundantes.
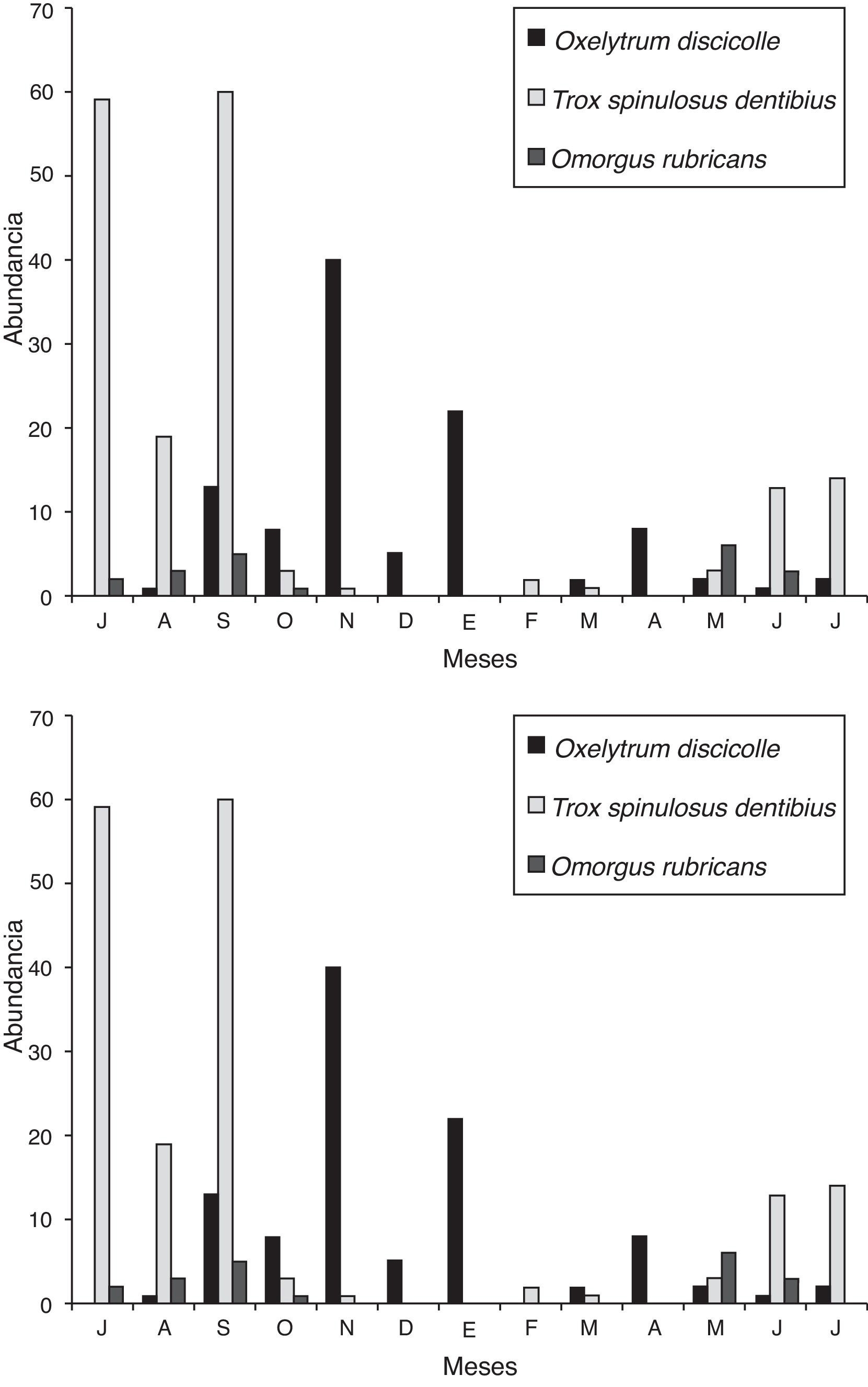
Figura 3 Estacionalidad de coleópteros necrócolos en el Bosque Los Colomos, Guadalajara, Jalisco, México.
La diversidad estimada de orden 0 (0D ), muestra diferencias importantes para cada uno de los sitios dentro del bosque. Eucalipto-II presentó la mayor diversidad en comparación con pino-I y casuarina-I que presentaron los valores más bajos. Al considerar la medida de diversidad de Orden 1 (1D ), el valor de máxima diversidad se presentó en eucalipto-I, seguido de eucalipto-II y pino-II (tabla 2).
Tabla 2 Valores de diversidad, diversidad de orden 0 (0 D) y diversidad de orden 1 (1 D).
| Diversidad | Sitios | ||||||
|---|---|---|---|---|---|---|---|
| Pino I | Eucalipto I | Casuarina I | Vegetación secundaria | Casuarina II | Eucalipto II | Pino II | |
| 0D | 2 | 6,4 | 3 | 5,5 | 4,5 | 8,5 | 7,5 |
| 1D | 0,38 | 1,75 | 0,93 | 1,21 | 0,56 | 1,57 | 1,39 |
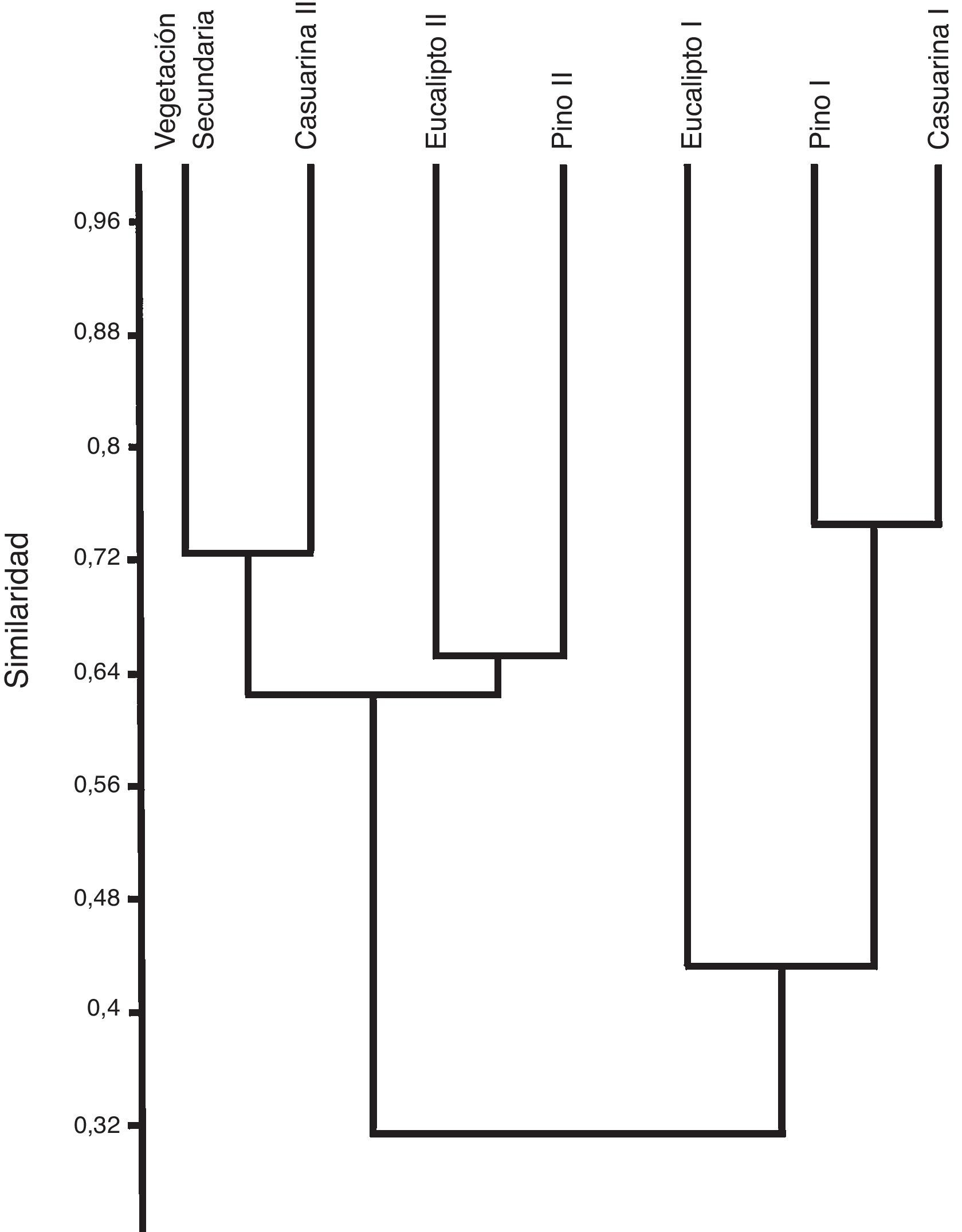
El ensamble de coleópteros necrócolos muestra la formación de 2 grupos bien diferenciados, estrechamente relacionados con las secciones del bosque en función de su grado de perturbación que corresponden a C-I y a C-II. Aquellos sitios dentro del bosque con vegetación similar se integraron en diferentes grupos. La vegetación secundaria (zona de transición) quedó incluida dentro del agrupamiento de C-II, sección que está caracterizada por la dominancia de T. spinulosus dentibius , y de 2 especies exclusivas (D. amplicollis y O. suberosus ). Los sitios de eucalipto-II y pino-II presentaron la mayor riqueza específica y una similitud del 65%, mientras que la vegetación secundaria y casuarina-II tuvieron una similitud del 72.5%. En CI, el valor más alto de similitud se presentó entre los sitios de pino y casuarina (74.5%), donde O. discicolle es la especie dominante. En este mismo grupo de CI, la similitud entre los sitios anteriores respecto de eucalipto-I fue del 43%, siendo este último el mejor representado de la sección CI (fig. 4).
Discusión
La diversidad de coleópteros necrócolos en el BLC es baja tanto en riqueza (S = 8) como en abundancia (N = 362), comparado con estudios similares realizados en otras localidades de Jalisco (Morón et al., 1988; Naranjo-López y Navarrete-Heredia, 2011; Rivera-Cervantes y García-Real, 1998; Quiroz-Rocha et al., 2008), donde la riqueza específica (S = 6-23) y la abundancia (N = 941-6,280) registrada fue mayor, inclusive con un esfuerzo de muestreo menor. En Jalisco, la mayor diversidad de coleópteros necrócolos se ha registrado en sitios con bosque de pino y pino-encino (Quiroz-Rocha et al., 2008; Rivera-Cervantes y García-Real, 1998), incluyendo bosque mesófilo (Quiroz-Rocha et al., 2008) y bosque tropical caducifolio (Naranjo-López y Navarrete-Heredia, 2011). En el BLC destaca la abundancia de T. spinulosus dentibius y la baja representatividad de especies de los Scarabaeinae. Las especies de Silphidae son referidas con frecuencia en diversos estudios sobre la entomofauna necrócola y se han registrado en bosque tropical y mesófilo (Navarrete-Heredia y Fierros-López, 2000; Rivera-Cervantes y García-Real, 1998). A O. discicolle y N. olidus se les ha encontrado cohabitando en zonas templadas, tropicales y subtropicales (Navarrete-Heredia, 2009), como sucedió en el BLC. Sin embargo, la explotación diferencial del recurso, la estacionalidad desfasada y la dominancia diferencial han sido interpretadas como cualidades que favorecen su coexistencia (Navarrete-Heredia y Fierros-López, 2000), situación que ha sido corroborada en estudios específicos. Por ejemplo, en un bosque mesófilo de montaña del centro de Veracruz Arellano, Favila y Huerta (2005) encontraron que ambas especies tienen su actividad temporal desfasada, situación que sugiere que la competencia por cadáveres es muy alta en estas especies y la separación temporal en sus ciclos de actividad sea un mecanismo que reduce la competencia entre ambas especies.
Los coleópteros necrócolos se encuentran activos todo el año, presentando la mayor riqueza durante los meses de junio y agosto (S = 7) y septiembre (S = 6) y la menor en febrero (S = 2) y marzo (S = 3), patrón similar observado en Malinalco, Estado de México (> junio y septiembre; < enero y febrero) (Trevilla-Rebollar, Deloya y Padilla-Ramírez, 2010) y Gómez Farías, Jalisco (> junio y octubre; < marzo y abril) (Naranjo-López y Navarrete-Heredia, 2011). La abundancia presentó 2 picos, uno en los meses con mayor precipitación, similar a lo observado en otros estudios similares (Deloya, Parra-Tabla y Delfín-González, 2007; Morón y Terrón, 1984), y otro en los meses más fríos. Tanto en el BLC como en Gómez Farías, las especies con mayor abundancia correspondieron a Silphidae.
De las 8 especies registradas en este trabajo, la mayoría se distribuye en los sitios del bosque. Seis especies cohabitan en las 2 secciones del bosque y O. suberosus y D. amplicollis son exclusivas de Colomos-II, por lo cual podrían considerarse en la categoría de raras a nivel local. Sin embargo, D. amplicollis ha sido registrada en la categoría de especie común, al igual que N. olidus en bosque de Pinus (Rivera-Cervantes y García-Real, 1998), mientras que D. amplicollis y O. suberosus se han observado en bosque tropical caducifolio y subcaducifolio (Morón et al., 1988); N. olidus , O. discicolle y D. amplicollis en bosque mesófilo y bosque de Pinus (Quiroz-Rocha et al., 2008). Dichotomius amplicollis solo representa el 6.3% de los ejemplares en BLC. Es importante mencionar que los sitios con eucalipto registraron la mayor riqueza y abundancia.
Trox spinulosus dentibius y O. discicolle son las especies dominantes en el BLC, aunque el predominio en cada sección fue diferente: O. discicolle en CI y T. spinulosus dentibius en CII. Estudios con especies indicadoras de escarabaeinos indican un efecto sobre su abundancia o preferencia por determinadas condiciones de cobertura vegetal (bosque, borde, potreros), causado por la acción antrópica, la fragmentación y el cambio de uso de suelo en selvas tropicales (Favila, 2004; Reyes-Novelo, Delfín-González y Morón, 2007). En el BLC 3 especies se localizaron en pocos sitios: D. amplicollis es exclusiva de CII, O. batesi de sitios de eucalipto y vegetación secundaria, y O. lecontei de los sitios de eucalipto y pino-II. Por sus escasas abundancias no tenemos la certeza de su preferencia hacia estos sitios o que sus bajas abundancias sean producto de las condiciones predominantes en el bosque que afecten su presencia en él.
Halffter y Arellano (2002) observaron que la abundancia de los escarabaeinos se ve favorecida por la cobertura vegetal, más que por la disponibilidad de alimento. En este estudio CII, por su grado de conservación y restauración presenta mayor densidad de vegetación, lo que podría explicar la mayor abundancia de los escarabaeinos en esta sección. Rivera-Cervantes y García-Real (1998) observaron que la abundancia disminuye en bosque de Pinus quemado con relación al bosque sin perturbación (utilizando los mismos índices de diversidad 1D ), aunque la diversidad observada fue alta en ambos bosques (Pinus conservado 1D = 4.334; bosque de Pinus quemado 1D = 3.760). Al interior del BLC, donde hay más actividad antrópica (pino-I), se obtuvo menor riqueza y abundancia en relación con pino-II y, por lo tanto, una similitud baja (36%). Asimismo, puede evidenciarse la diferencia entre sitios con el mismo tipo de vegetación (ubicada en las diferentes secciones), por lo cual se podría considerar que las especies estarían asociadas a los sitios con mejor calidad, de aquellos alterados, debido probablemente a que en estos últimos la infraestructura para actividades de recreación humana y las actividades de mantenimiento del bosque, como remoción de cadáveres y excretas de caballo, poda de maleza, aplicación de fungicidas e insecticidas, generan condiciones desfavorables para este grupo de insectos.
Los valores de diversidad verdadera evidencian una diversidad variable dentro de los sitios del bosque. Destaca el valor de orden 1 (1D ) para eucalipto-II (1D = 5.729), que fue similar al reconocido para el bosque mesófilo en Mascota, Jalisco (1D = 5.723), a pesar de las diferencias en el número de especies y abundancia (S = 18, N = 3,001) (Quiroz-Rocha et al., 2008). Un valor similar también fue obtenido en un estudio realizado en localidades con bosque de pino (D1 = 5.042, S = 15, n = 506) en Gómez Farías (Naranjo-López y Navarrete-Heredia, 2011). Por el contrario, los valores fueron bajos en localidades con bosque tropical caducifolio (1D = 3.987, S = 28, n = 2,027). A diferencia de lo observado en otros estudios, en los cuales los Scarabaeidae (Scarabaeinae), Silphidae y Staphylinidae son las familias mejor representadas (Deloya, 1996; Deloya et al., 1987; Morón y Terrón, 1984; Naranjo-López y Navarrete-Heredia, 2011; Sánchez-Ramos, Lobo, Lara-Villalón y Reyes-Castillo, 1993), en el BLC predominó Staphylinidae y Trogidae, situación especial para esta última familia, ya que con frecuencia se ha registrado una baja abundancia en otros estudios (Deloya, 2003; Deloya et al., 1987; Díaz, Galante y Favila, 2010; Morón et al., 1988; Trevilla-Rebollar et al., 2010). Como parte del paisaje urbano de la zona metropolitana de Guadalajara, el BLC, en cuanto a la composición y abundancia de coleópteros Silphidae, Scarabaeinae y Trogidae, muestra una composición y valores de diversidad de especies baja en comparación con otras zonas en el estado de Jalisco. ¿Qué factores son los causantes de tal situación? Aunque se pueden sugerir algunos de ellos, como la presencia de áreas amplias de vegetación exótica, el impacto por las actividades humanas o el uso de químicos para el control de plagas, es necesaria la realización de estudios específicos para evaluar algunas de estas variables y tener respuestas satisfactorias.
Agradecimientos
Queremos agradecer especialmente a la Dra. Gabriela Castaño-Meneses por su apoyo en la realización del análisis de covarianza canónica. Este trabajo es producto de la investigación de tesis de maestría del primer autor en el programa BIMARENA del CUCBA, Universidad de Guadalajara. Al Dr. Mario E. Favila por sus comentarios y sugerencias para mejorar el documento.

