











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los registros de recolecta son una fuente de información valiosa en el estudio de la distribución geográfica de las especies, debido a que permiten relacionar su presencia con atributos ambientales que permiten predecir su aparición (Elith y Leathwick, 2009; Faith et al., 2013; Guisan y Thuillier, 2005; Guisan y Zimmermann, 2000; Hirzel y Lay, 2008). Tales predicciones son un medio para identificar patrones en la riqueza y composición de especies a distintas escalas espaciales, y pueden utilizarse tanto para definir como para priorizar áreas de manejo y conservación de los recursos naturales (Anderson y Martínez-Meyer, 2004; Araujo, 1998; Araujo y New, 2009). Los algoritmos utilizados para predecir la distribución geográfica de las especies parten de los supuestos estadísticos de que los registros fueron obtenidos de forma aleatoria, que las muestras fueron independientes entre sí y que representan las condiciones ambientales en donde se encuentra la especie (Albert et al., 2010; Austin, 2002; Boitani y Fuller, 2000; Hirzel y Lay, 2008). La aleatoriedad se da cuando una muestra se obtiene sin un patrón evidente, y es necesaria para obtener un estimado cercano a la media poblacional (Krebs, 1999; Lájer, 2007). La violación de este supuesto sucede cuando ciertas áreas o condiciones ambientales son mejor representadas que otras (Botts, Erasmus y Alexander, 2011; Parnell et al., 2003; Sokal y Rohlf, 1995). La independencia sucede cuando un evento no influye positiva o negativamente en la elección de una nueva muestra (Albert et al., 2010), y este supuesto es violado cuando el diseño de muestreo se basa en recolectas previas o cuando se incrementa el esfuerzo de recolecta en áreas donde ya se ha registrado la especie (Küper, Sommers, Lovett y Barthlott, 2006; Madin y Lyon, 2005; Romo, García-Barros y Lobo, 2006). Finalmente, la representatividad se obtiene cuando se incluye la mayor variación ambiental de los sitios en que potencialmente la especie puede encontrarse. La representatividad no se cumple cuando se omiten áreas en las que la especie se da en bajas densidades (Nelson, Rerreira, Silva y Kawasaki, 1990; Wolf, Anderegg, Ryan y Christian, 2011). La omisión de los tres supuestos incrementa la posibilidad de sobre o subestimar el parámetro de interés, que a su vez deriva en la generación de conclusiones erróneas (Dennis, Sparks y Hardy, 1999).
En la última década, se ha incrementado el uso de los registros de recolecta para predecir la distribución geográfica de las especies, principalmente por el aumento en la disponibilidad de bases de colecciones biológicas y por el desarrollo de mejores algoritmos de predicción (Droissart, Hardy, Sonké, Dahdouh-Guebas y Stévart, 2012; Faith et al., 2013; Gaiji et al., 2013). Actualmente, existe una gran variedad de bases de datos de acceso libre para grupos taxonómicos específicos (VerNet, MaNIS, OrNis, HerpNet o FishNet2) o de diferentes taxa (GBIF o Conabio). Sin embargo, debe tomarse en cuenta que tales registros fueron obtenidos para resolver distintos objetivos, por lo que reflejan diferentes estrategias de muestreo que, en conjunto, pueden incrementar el riesgo de violar los supuestos de aleatoriedad, independencia y representatividad de los análisis (Feeley y Silman, 2011; Madin y Lyon, 2005). Tales riesgos varían dependiendo del algoritmo de predicción; por ejemplo, cuando existe un sesgo en la elección de los registros (áreas mejor evaluadas que otras), los algoritmos que utilizan solo presencias son más afectados que los modelos de presencia-ausencia (Elith et al., 2011). Aun cuando existen algoritmos que utilizan solo presencias y que parten del supuesto de que existen áreas mejor evaluadas que otras (Elith et al., 2011; Phillips y Dudik, 2009), como el de Máxima Entropía (MaxEnt), se ha observado que excluir parte de las condiciones ambientales en las que habita una especie modifica de forma sustancial la predicción (Lozier, Aniello y Hickerson, 2009). Actualmente, los algoritmos que usan solo presencias son los más utilizados, debido a que los algoritmos que utilizan datos de abundancia resultan muy costosos en términos de la obtención de tal información. Los modelos de presencia-ausencia corren el riesgo de utilizar falsos-positivos, es decir, asumir que la especie está ausente cuando en realidad está presente. Por último, la mayoría de las bases de registros de recolecta solo registran presencias y no ausencias (Albert et al., 2010; Hirzel y Lay, 2008). Así, es necesario evaluar el cumplimiento de la aleatoriedad, independencia y representatividad de los registros de recolecta para obtener conclusiones válidas, robustas y precisas de los patrones de distribución de las especies obtenidas a partir de los algoritmos que utilizan solo presencias.
Los sesgos espaciales pueden ser resultado de mayores esfuerzos de recolecta en áreas cercanas a zonas urbanas (Dennis et al., 1999; Soberón, Llorente y Oñate, 2000), a vías de comunicación (Freitag, Hobson, Biggs y Jaarsveld, 1998; Parnell et al., 2003), al hogar de los colectores (Küper et al., 2006) o a áreas prioritarias para la conservación que comúnmente resultan atractivas para los investigadores (Hortal, García-Pereira y García-Barros, 2004; Reddy y Davalos, 2003; Sánchez-Fernández, Lobo, Abellán, Ribera y Millán, 2008; Sastre y Lobo, 2009). La distancia a áreas urbanas y vías de comunicación son fuentes de variación de particular relevancia, debido a que los recolectores generalmente procuran cubrir una mayor área, a menor costo y en el menor tiempo (Botts et al., 2011; Parnell et al., 2003). El riesgo de utilizar la anterior estrategia es que los registros tienden a estar agregados y solo se capturan las condiciones ambientales asociadas a las áreas urbanas o vías de comunicación, excluyendo las condiciones ambientales de las áreas menos accesibles (Romo et al., 2006; Tobler, Honorio, Janovec y Reynel, 2007).
La predicción de la distribución geográfica de una especie suele realizarse con el objetivo de predecir la distribución completa, a partir del conocimiento parcial con el que se cuenta (Elith y Leathwick, 2009; Hirzel y Lay, 2008). Tal es el caso de las especies de gastrópodos, pteridofitas y plantas acuáticas en el estado de Tamaulipas, de las cuales existe un notable esfuerzo de recolecta en la parte centro-sur del estado, pero no así en la parte norte (Barrientos-Lozano et al., 2005; Correa-Sandoval, Martínez, Horta y Castro, 2012; Mora-Olivo y Villaseñor, 2007; Mora-Olivo, Villaseñor, Luna-Vega y Morrone, 2008). Por lo anterior, estas especies son candidatas a predecir su distribución geográfica en la región; pero, antes de ello, es necesario analizar la aleatoriedad, independencia y representatividad de los registros de recolecta. Así, este estudio estimó la aleatoriedad, la independencia y la representatividad ambiental de los registros de recolecta de estas especies en el estado de Tamaulipas en función de la distancia a áreas urbanas y vías de comunicación. El estudio parte de la hipótesis de que si estos 3 grupos de especies, que tienen requerimientos de nicho distintos, presentan violaciones similares a estos 3 supuestos estadísticos, entonces el sesgo espacial se debió a un mayor esfuerzo de recolecta en áreas previamente evaluadas, cercanas a vías de comunicación y que solo reflejan las condiciones ambientales de estas áreas.
Materiales y métodos
Los grupos de especies analizados incluyeron las especies de gastrópodos terrestres, pteridofitas y plantas acuáticas del estado de Tamaulipas. Los 3 grupos tienen requerimientos de nicho distintos y una dependencia diferencial al agua, por lo que se encuentran en condiciones ambientales distintas. Los gastrópodos tienen mayor afinidad a ambientes húmedos, así como a la vegetación tropical y templada (Correa-Sandoval, Strenth, Rodríguez-Castro y Horta-Vega, 2009). Las pteridofitas también requieren de ambientes húmedos, pero necesitan de agua en estado líquido durante el desarrollo de la espora a la planta; una vez que se establecen pueden tolerar distintos climas (desiertos, tundras, bosques templados o tropicales). Por último, para poder desarrollarse de forma adecuada, las plantas acuáticas requieren de cuerpos de agua permanentes o, al menos, de suelo saturado (Sculthorpe, 1985).
Los registros de recolecta de los 3 grupos de especies se obtuvieron de 2 bases de datos disponibles en la red (GBIF y Conabio), de tesis y de artículos publicados (Correa-Sandoval y Thompson, 2005; Correa-Sandoval et al., 2012; Mora-Olivo y Villaseñor, 2007; Mora-Olivo et al., 2008). Las localidades utilizadas contaron al menos con su descripción o las coordenadas geográficas; en caso de que solo se registrara el nombre de la localidad, las coordenadas se obtuvieron de cartas topográficas (escala 1:50,000; Inegi) o de Google Earth. En total, se utilizaron 169 localidades únicas para las especies de gastrópodos, 714 localidades únicas de las plantas acuáticas y 183 localidades únicas para las pteridofitas. El 12.2% de las especies de gastrópodos contaron con al menos 7 localidades únicas de recolecta (28 especies), que es el número mínimo de localidades sugeridas para generar los mapas de distribución geográfica (Phillips, Anderson y Schapire, 2006). En el caso de las plantas acuáticas se generaron mapas de distribución potencial en el 42% de las especies (168 especies), y en el 20.2% de las especies de pteridofitas (163 especies).
Aleatoriedad
La disposición espacial de las localidades de recolecta puede ser regular, aleatoria o agregada (Gotelli y Ellison, 2004; Krebs, 1999). La disposición espacial aleatoria, simple o estratificada, se utiliza con frecuencia en el diseño del muestreo de especies, debido a que evita seleccionar áreas en las que previamente se ha registrado y reduce el riesgo de la autocorrelación espacial (Albert et al., 2010). En la disposición agregada, en cambio, los registros están concentrados en una pequeña fracción del área de estudio, y en la disposición regular estos tienen una separación espacial constante (Krebs, 1999). En este estudio se determinó el tipo de disposición espacial al que se ajustaron los registros de recolecta de los 3 taxa y, para ello, se les aplicó la prueba de aleatoriedad de Hopkins (h ) para mapas incompletos (Hopkins, 1954 en Krebs, 1999). La prueba estima el grado de separación que existe entre las localidades observadas con respecto a localidades generadas al azar, bajo una distribución uniforme, en que valores cercanos a 1.0 significan que las localidades tienden a presentar una disposición azarosa; valores menores de 1.0, disposiciones regulares, y valores mayores de 1.0, disposiciones agregadas. La prueba se calculó a partir de la fórmula:
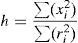
donde: h es la prueba de aleatoriedad de Hopkins; x2i es la distancia cuadrática de cada una de las localidades observadas al vecino aleatorio más cercano, y r2i es la distancia cuadrática de las localidades observadas al vecino observado más cercano.
Las localidades aleatorias se generaron en el programa ArcGis 10.1 (ESRI, 2002), a una distancia mínima de separación de 10 m, debido a que fue la distancia mínima observada entre las 2 localidades más cercanas. El número de localidades generadas fue igual al número de localidades observadas, las cuales se distribuyeron solo en el estado de Tamaulipas (extensión de 7,806.000 ha) y se ajustaron a una distribución uniforme. El análisis se realizó para cada taxón, y para cada uno de ellos se utilizaron diferentes localidades generadas al azar. El número de localidades generadas al azar fue igual al de localidades observadas para cada grupo (169 únicas para las especies de gastrópodos, 714 para las plantas acuáticas y 183 para pteridofitas). Para cada una de las localidades observadas se calculó la distancia euclidiana entre la localidad vecina observada más cercana (x i) y la localidad más cercana generada al azar (r i). La prueba de hipótesis se realizó contrastando el valor de h , calculado con respecto al valor teórico obtenido de la distribución de F de Fisher, con 2n (u ) y 2n (v ) grados de libertad, debido a que los valores de la prueba se ajustan a la distribución de F (Krebs, 1999). Los valores de F se obtuvieron de la distribución de F documentada por Sokal y Rohlf (1995). La regla de decisión para esta prueba fue: a) si el valor de h calculado fue menor que el valor teórico de F a una probabilidad de 0.975, entonces se sugiere una disposición regular; b) cuando el valor de h fue mayor que el valor de F a una probabilidad de 0.025, entonces se presentó una disposición agregada, y por ultimo c) si el valor de h se encontró entre el valor de F a una probabilidad de 0.975 y 0.025, entonces se presentó una disposición aleatoria. Además, se obtuvo el índice de disposición (I h) que toma valores de 0.0 a 1.0:

El índice se aproxima a 1.0 cuando la agregación se incrementa y 0.0 cuando la uniformidad es máxima; se consideró como aleatorio cuando el valor osciló alrededor del valor de 0.5.
Independencia a vías de comunicación o áreas urbanas
De forma ideal los registros de recolecta tendrían que ser independientes de atributos espaciales, que incrementan o reducen la probabilidad de ser registrados (Albert et al., 2010). En el presente estudio solo se analizó la asociación de las localidades de recolecta con áreas urbanas y vías de comunicación con el sesgo espacial, pero debe considerarse que existen otros factores que incrementan tal sesgo, como la distancia a áreas de conservación, al hogar de los colectores o a la disponibilidad de agua. Comúnmente existe un mayor esfuerzo de recolecta en áreas de fácil acceso, como son los caminos, carreteras o áreas urbanas, debido a que los costos para la obtención de estos registros se reducen (Botts et al., 2011). El resultado es que se tiende a estimar parámetros sesgados e imprecisos de la respuesta de las especies a la variación ambiental (Albert et al., 2010) y esto, por lo tanto, afecta la predicción de la distribución geográfica de la especie. La prueba de independencia a áreas de fácil acceso se realizó con el análisis de correspondencia, la cual es una técnica descriptiva/exploratoria multivariada modificada de la prueba de la X2, que está diseñada para analizar tablas de contingencia de dos o más vías (Gotelli y Ellison, 2004).
En este estudio se estimó el grado de asociación entre el intervalo de distancia a zonas urbanas-vías de comunicación y el tipo de localidad, ya sea observada o generada al azar. La variable categórica "intervalo de distancia a las zonas urbanas-vías de comunicación" incluyó 6 categorías, 3 representaron la cercanía a las vías de comunicación (a 1, 3 y 7 km) y 3, la cercanía a zonas urbanas (0, 1 y 3 km); mientras que la variable "tipo de localidad" consideró 2 categorías: localidades observadas y localidades generadas al azar. Las localidades generadas al azar fueron las mismas que las utilizadas en la prueba de aleatoriedad. A cada localidad se le asignó una categoría de intervalo de distancia a zonas urbanas-vías de comunicación, dependiendo de la distancia que presentó con respecto a las vías de comunicación-áreas urbanas y al tipo de localidad, ya sea observada o generada al azar. Esta clasificación se utilizó para generar una matriz de contingencia entre la frecuencia de localidades por intervalo de vía de comunicación-zona urbana y el tipo de localidad. A la matriz se le aplicó el análisis de correspondencia para crear un menor número de dimensiones que representan la asociación entre las categorías que componen a las 2 variables categóricas (Gotelli y Ellison, 2004). Las categorías que se encontraron cercanas en la gráfica de dispersión presentaron una mayor asociación (frecuencia) que aquellos pares de categorías que se encontraron distantes. La hipótesis nula a contrastar fue que las frecuencias observadas por intervalo de distancia a zonas urbanas-vías de comunicación fueron similares con respecto a registros obtenidos al azar, con un nivel de significancia de 0.05.
Representatividad ambiental
La representatividad ambiental de las localidades de recolecta se estimó comparando las condiciones ambientales asociadas a las localidades observadas con las condiciones ambientales de las localidades generadas al azar. Las localidades se agruparon en los 6 intervalos de distancia a las vías de comunicación y zonas urbanas, tal como se hizo en la prueba de independencia. Las variables ambientales utilizadas para comparar los grupos de localidades fueron climáticas (WorldClim), el porcentaje de cobertura vegetal (Global Land Cover Facility) y la elevación (WorldClim; tabla 1), las cuales presentaron una resolución espacial de 30 arco segundos (≈ 1 km). Las comparaciones se realizaron con el análisis de funciones discriminantes lineales, un análisis multivariado que tiene como objetivo estimar diferencias significativas entre grupos e identificar las variables asociadas a tales diferencias (Legendre y Legendre, 1998). Este análisis es una extensión de la prueba de ANOVA, que maximiza las diferencias entre grupos y minimiza las variaciones dentro de ellos, pero incluye más de una variable independiente (Gotelli y Ellison, 2004). El análisis creó combinaciones lineales de las variables originales, que extrajeron la mayor variación entre los grupos (raíces). El número de raíces generadas fue igual al número de grupos comparados. La interpretación de los resultados se llevó a cabo con las raíces que aportaron variación significativa (prueba sucesiva de la X2) o cuyo valor acumulado aportara, al menos el 75%, de la variación total. La posición promedio de los grupos (centroides) fue graficada en las raíces significativas. Las variables utilizadas para interpretar las diferencias entre grupos fueron las que presentaron el valor de la estructura de factores mayor de 0.6 o menor de -0.6. El valor de la estructura de factores refleja la correlación existente entre el valor de las variables originales con respecto a los nuevas variables generadas (raíces). Por último, se determinó entre qué pares de grupos de localidades se presentaron diferencias significativas con un nivel de significancia de 0.05.
Tabla 1 Variables utilizadas para comparar las condiciones ambientales entre los intervalos de distancia a zonas urbanas-vías de comunicación de los registros observados de las especies de gastrópodos y los registros generados al azar.
| Variable | Abreviación | Unidad | Fuente |
|---|---|---|---|
| Temperatura mínima registrada en el año | Tmín | °C | http://www.worldclim.org/ |
| Precipitación media mensual | PP prom | mm | http://www.worldclim.org/ |
| Precipitación mínima en el trimestre más lluvioso | PP PMICLL | mm | http://www.worldclim.org/ |
| Temperatura promedio mensual | Tprom | °C | http://www.worldclim.org/ |
| Precipitación máxima en el trimestre más seco | PMACS | mm | http://www.worldclim.org/ |
| Porcentaje de herbáceas en 1 km | % herbáceas | porcentaje | http://glcfapp.glcf.umd.edu/ |
| Porcentaje de árboles en 1 km | % árboles | porcentaje | http://glcfapp.glcf.umd.edui/ |
| Porcentaje de árboles de hoja ancha en 1 km | % árboles hoja ancha | porcentaje | http://glcfapp.glcf.umd.edui/ |
| Altitud | altitud | m snm | http://www.worldclim.org/ |
| Porcentaje de suelo desnudo en 1 km | % suelo desnudo | porcentaje | http://glcfapp.glcf.umd.edu/ |
Resultados
Los análisis de disposición espacial sugieren que los registros de recolecta de las especies de gastrópodos (h = 46.6 > Fg.l.169,169 = 1.23, I h = 0.92), plantas acuáticas (h = 12.2 > Fg.l.183,183 = 1.23, I h = 0.9) y pteridofitas (h = 3.6 > Fg.l.714,714 = 1.1, I h = 0.98) presentaron una disposición espacial agregada con respecto a lo esperado al azar. Asimismo, los registros de recolecta de las especies de pteridofitas (eigenvalor = 0.18, Xgl. = l5 = 71.2, p < 0.0001, figura 1a), gastrópodos (eigenvalor = 0.04, Xgl = l5 = 15.1, p = 0.01; figura 1b) y plantas acuáticas (eigenvalor = 0.18, Xgl = l5 = 250.8, p < 0.0001; figura 1c) tendieron a concentrarse con mayor frecuencia cerca de zonas urbanas (1 y 3 km) y con menor frecuencia a una distancia mayor de 1 km de las vías de comunicación que lo esperado al azar.

Figura 1 Posición canónica de los registros observados y generados al azar de las especies de gastrópodos (a), pteridofitas (b) y plantas acuáticas (c) en los intervalos de distancia a zonas urbanas y vías de comunicación.
Las condiciones ambientales de los registros obtenidos de forma aleatoria, agrupados en los 6 intervalos de distancia a áreas urbanas y vías de comunicación, fueron diferentes, excepto entre los intervalos a 0 y 1 km de las zonas urbanas (tabla 2). Por lo anterior, cada categoría de distancia a vías de comunicación o zonas urbanas refleja una variación ambiental distinta. Las localidades de recolecta de las especies de gastrópodos que se encontraron a una distancia de 0, 1 y 3 km de las zonas urbanas presentaron características ambientales similares, lo cual también ocurrió entre las localidades que se encontraron a 1, 3 y 7 km de las vías de comunicación (lambda de Wilks: 0.6, Fg.l. = 110,6613 = 4.2, p < 0.0000). Los registros cercanos a zonas urbanas se caracterizaron por asociarse a áreas con menos del 15.7% de árboles, de los cuales no más del 20.1% correspondieron a árboles de hoja ancha; con más del 73.7% de herbáceas, bajos porcentajes de suelo desnudo (menores del 9.7%) y una precipitación mayor de los 81.5 mm en el cuatrimestre más seco del año (figura 2a). En contraste, los registros que se encontraron cercanos a las vías de comunicación se caracterizaron por asociarse a más del 15.7% de árboles, de los cuales más del 20.1% correspondieron a árboles de hoja ancha; con menos del 73.7% de herbáceas, altos porcentajes de suelo desnudo (más del 9.7%) y una precipitación menor de los 81.5 mm en el cuatrimestre más seco del año. Las condiciones ambientales menos exploradas para las especies de gastrópodos fueron las que se encontraron en áreas con menos del 15.7% de árboles, de los cuales no más del 20.1% correspondieron a árboles de hoja ancha; con más del 73.7% de herbáceas, porcentajes de suelo desnudo mayores al 9.7% y con una precipitación por debajo de los 81.5 mm en el cuatrimestre más seco (figs. 2a1 y a2).
Tabla 2 Valores de p (diagonal superior) y F (diagonal inferior, g. l.= 10; 882) derivados de la comparación de las condiciones ambientales entre los intervalos de distancia a zonas urbanas-vías de comunicación de los registros observados de las especies de gastrópodos y los registros generados al azar el estado de Tamaulipas, México.
| Registros observados | Registros azarosos | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Urbana | Vías | Urbana | Vías | |||||||||
| 0 km | 1 km | 3 km | 1 km | 3 km | 7 km | 0 km | 1 km | 3 km | 1 km | 3 km | 7 km | |
| Obs 0 km urbana | 0.44 | 0.35 | 0.02 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 1 km urbana | 1.00 | 0.37 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 3 km urbana | 1.11 | 1.09 | 0.01 | 0.00 | 0.00 | 0.11 | 0.10 | 0.03 | 0.00 | 0.00 | 0.00 | |
| Obs 1 km vías | 2.18 | 2.85 | 2.27 | 0.42 | 0.15 | 0.06 | 0.01 | 0.01 | 0.06 | 0.22 | 0.02 | |
| Obs 3 km vías | 4.08 | 6.09 | 3.77 | 1.02 | 0.25 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | |
| Obs 7 km vías | 3.29 | 4.84 | 2.66 | 1.47 | 1.26 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | |
| Azar 0 km urbana | 3.05 | 3.71 | 1.57 | 1.80 | 6.21 | 4.84 | 0.06 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Azar 1 km urbana | 3.65 | 4.54 | 1.60 | 2.39 | 6.71 | 5.47 | 1.78 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Azar 3 km urbana | 6.74 | 7.19 | 2.00 | 2.46 | 8.38 | 8.31 | 4.69 | 2.55 | 0.00 | 0.00 | 0.00 | |
| Azar 1 km vías | 5.87 | 8.23 | 3.94 | 1.79 | 2.46 | 2.23 | 8.54 | 6.46 | 10.36 | 0.00 | 0.00 | |
| Azar 3 km vías | 5.00 | 6.92 | 3.25 | 1.31 | 3.29 | 2.57 | 3.31 | 4.43 | 7.25 | 3.43 | 0.00 | |
| Azar 7 km vías | 5.41 | 6.27 | 2.80 | 2.12 | 6.29 | 5.07 | 4.29 | 2.70 | 3.88 | 5.45 | 3.10 | |
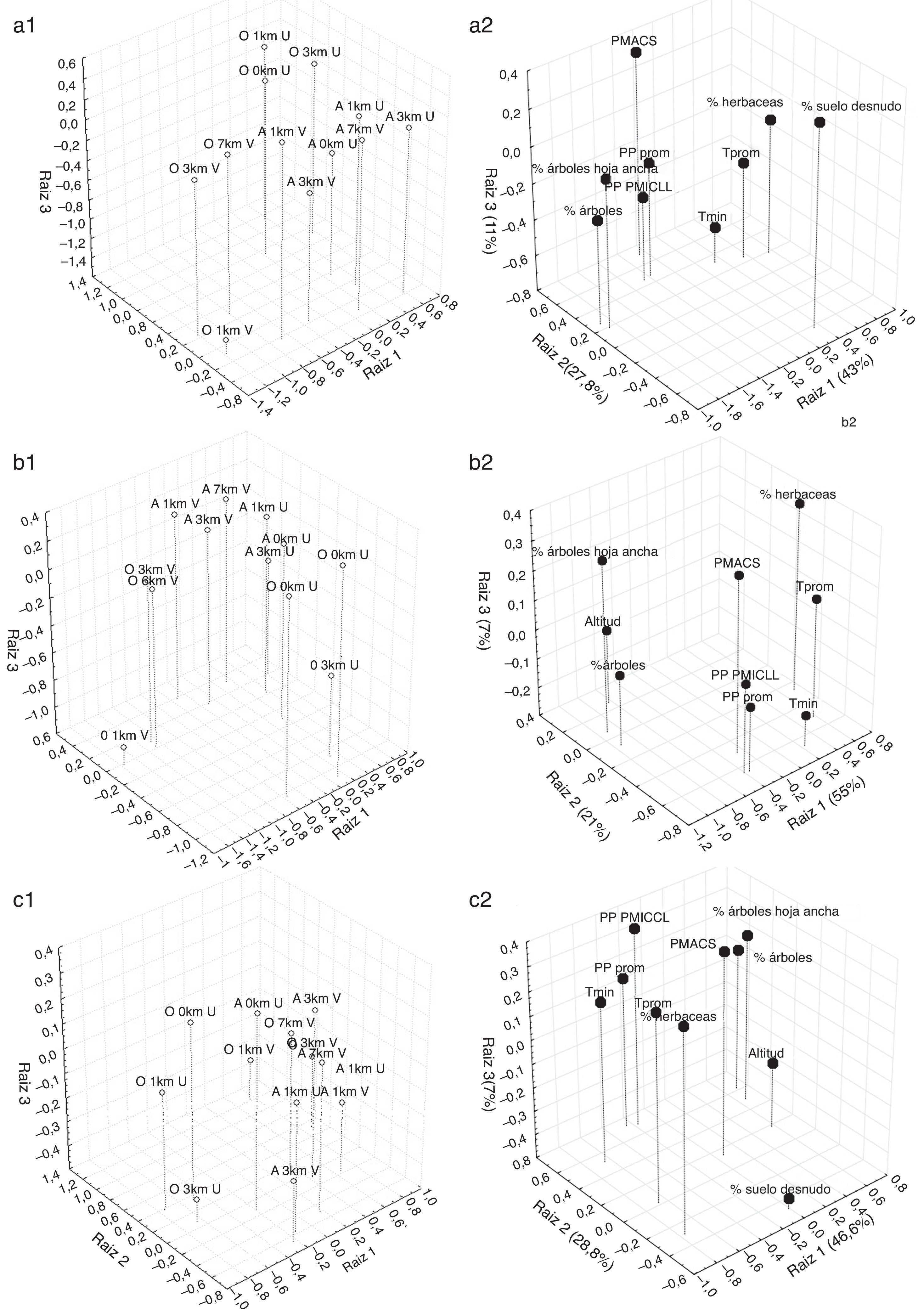
Figura 2 Centroides de los registros observados (O) y generados al azar (A) de las especies de gastrópodos (a1), pteridofitas (b1) y plantas acuáticas (c1) en los intervalos de distancia a zonas urbanas (U) y vías de comunicación (V), así como la estructura de las primeras3 raíces para los3 grupos de especies (a2, b2, y c2, respectivamente).
Los registros de recolecta de las pteridofitas en los 6 intervalos de distancia a zonas urbanas y vías de comunicación resultaron estadísticamente diferentes entre sí (lambda de Wilks: 0.51, Fg.l. = 110,7444 = 6.2, p < 0.0000), excepto entre las áreas a 3 y 7 km a las vías de comunicación (tabla 3). Las localidades de recolecta observadas se encontraron en condiciones ambientales distintas a las localidades obtenidas de forma azarosa. Los registros cercanos a zonas urbanas se asociaron a áreas con más del 19.4% de árboles, de los cuales más del 24.2% correspondieron a árboles de hoja ancha (figs. 2b1 y b2). En contraste, los registros que se encontraron cercanos a las vías de comunicación se caracterizaron por asociarse a una precipitación promedio mensual menor de 63.8 mm y una temperatura mínima anual por debajo de los 10.6 °C. Las condiciones ambientales menos exploradas para las especies de pteridofitas fueron aquellas que se encontraron en áreas con una precipitación promedio mensual mayor de 63.8 mm y una temperatura mínima anual por encima de los 10.6 °C (figs. 2b1 y b2).
Tabla 3 Valores de p (diagonal superior) y F (diagonal inferior, g. l.= 10; 993) derivados de la comparación de las condiciones ambientales entre los intervalos de distancia a zonas urbanas-vías de comunicación de los registros observados de las especies de pteridofitas y los registros generados al azar en el estado de Tamaulipas, México.
| 0 | Registros observados | Registros azarosos | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | Urbana | Vías | Urbana | Vías | ||||||||
| 0 | 0 km | 1 km | 3 km | 1 km | 3 km | 7 km | 0 km | 1 km | 3 km | 1 km | 3 km | 7 km |
| Obs 0 km urbana | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 1 km urbana | 2.28 | 0.03 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 3 km urbana | 3.11 | 2.03 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 1 km vías | 3.88 | 6.90 | 5.29 | 0.01 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 3 km vías | 3.42 | 7.42 | 6.19 | 2.41 | 0.94 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 7 km vías | 4.26 | 10.35 | 8.16 | 2.26 | 0.42 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Azar 0 km urbana | 3.86 | 3.97 | 3.40 | 6.22 | 8.57 | 14.50 | 0.05 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Azar 1 km urbana | 5.72 | 5.88 | 4.15 | 7.49 | 9.79 | 17.33 | 1.86 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Azar 3 km urbana | 7.46 | 9.54 | 4.20 | 8.17 | 12.85 | 23.50 | 5.06 | 2.66 | 0.00 | 0.00 | 0.00 | |
| Azar 1 km vías | 4.70 | 8.86 | 6.32 | 4.81 | 3.21 | 6.27 | 8.01 | 5.95 | 9.70 | 0.00 | 0.00 | |
| Azar 3 km vías | 3.90 | 7.56 | 5.71 | 4.76 | 4.07 | 7.58 | 3.13 | 4.30 | 7.10 | 3.35 | 0.00 | |
| Azar 7 km vías | 6.31 | 7.89 | 5.85 | 7.00 | 8.03 | 14.81 | 4.36 | 2.70 | 3.83 | 5.14 | 3.09 | |
Por último, los registros de recolecta de las plantas acuáticas en los 6 intervalos de distancia a zonas urbanas y vías de comunicación resultaron estadísticamente diferentes entre sí (lambda de Wilks: 0.6, Fg.l. = 110,11507 = 7.2, p < 0.0000; tabla 4). Los registros de recolecta observados sucedieron en condiciones ambientales distintas a los registros obtenidos de forma azarosa (tabla 4). Los registros cercanos a zonas urbanas se encontraron en áreas con porcentajes de árboles menores del 16.3%, de los cuales menos del 20.3% correspondieron a árboles de hoja ancha; con más del 73% de cobertura de herbáceas y con una temperatura media mensual mayor de los 22.8 °C. En contraste, los registros que se encontraron cercanos a vías de comunicación se asociaron a porcentajes de árboles mayores del 16.3%, de los cuales más del 20.3% correspondieron a árboles de hoja ancha; con un menos del 73% de cobertura de herbáceas y con una temperatura media mensual por debajo de los 22.8 °C (figs. 2c1 y c2). Las condiciones ambientales menos exploradas fueron aquellas que presentaron una precipitación mínima menor a los 101 mm durante el cuatrimestre más lluvioso del año y con una precipitación promedio mensual menor a los 65.2 mm.
Tabla 4 Valores de p (diagonal superior) y F (diagonal inferior, g. l.= 10; 1,536) derivados de la comparación de las condiciones ambientales entre los intervalos de distancia a zonas urbanas-vías de comunicación de los registros observados de las especies de plantas acuáticas y los registros generados al azar en el estado de Tamaulipas, México.
| Registros observados | Registros azarosos | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Urbana | Vías | Urbana | Vías | |||||||||
| 0 km | 1 km | 3 km | 1 km | 3 km | 7 km | 0 km | 1 km | 3 km | 1 km | 3 km | 7 km | |
| Obs 0 km urbana | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 1 km urbana | 3.79 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 3 km urbana | 4.37 | 2.65 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 1 km vías | 6.52 | 7.84 | 7.05 | 0.03 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 3 km vías | 11.96 | 15.75 | 12.93 | 2.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Obs 7 km vías | 11.11 | 15.17 | 10.94 | 4.07 | 3.61 | 0.00 | 0.00 | 0.00 | 0.00 | 0.14 | 0.00 | |
| Azar 0 km urbana | 3.93 | 5.98 | 4.74 | 6.76 | 9.31 | 4.83 | 0.03 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Azar 1 km urbana | 8.11 | 9.99 | 5.62 | 9.12 | 11.58 | 7.88 | 2.00 | 0.05 | 0.00 | 0.00 | 0.01 | |
| Azar 3 km urbana | 9.94 | 11.74 | 5.11 | 9.80 | 12.68 | 10.95 | 4.27 | 1.87 | 0.00 | 0.00 | 0.00 | |
| Azar 1 km vías | 12.35 | 16.82 | 11.46 | 5.60 | 4.67 | 4.98 | 7.96 | 6.02 | 9.26 | 0.00 | 0.00 | |
| Azar 3 km vías | 8.52 | 12.74 | 9.27 | 4.87 | 4.18 | 1.49 | 3.05 | 4.54 | 6.70 | 3.39 | 0.00 | |
| Azar 7 km vías | 10.30 | 13.56 | 8.68 | 8.03 | 9.53 | 5.68 | 3.97 | 2.50 | 3.73 | 5.25 | 3.11 | |
Discusión
La disponibilidad de información de colecciones nacionales e internacionales ha permitido analizar interrogantes apremiantes sobre la pérdida de especies a escala local, regional o mundial (Gaiji et al., 2013). La mayoría de los registros de recolecta se han obtenido con objetivos muy específicos, como realizar inventarios taxonómicos, estudios poblacionales o comunitarios locales, y ha sido hasta recientemente que se ha promovido la obtención de datos para múltiples propósitos (Faith et al., 2013). Los registros de recolecta presentan sesgos espaciales, debido a que solo representan una fracción del ambiente en donde puede estar presente una especie (Gaiji et al., 2013). La accesibilidad y el atractivo científico de una área geográfica son 2 de los principales sesgos espaciales que se han registrado en las localidades de recolecta (Dennis et al., 1999; Freitag et al., 1998; Küper et al., 2006; Tobler et al., 2007). Ambas fuentes de sesgo reflejan una estrategia de muestreo distinto: por un lado, la accesibilidad se asocia a un mayor muestreo en áreas cercanas a zonas urbanas o caminos (Parnell et al., 2003), y por otro, el atractivo científico se asocia a un mayor esfuerzo de recolecta en áreas que poseen atributos ecológicos-evolutivos relevantes (Sastre y Lobo, 2009).
En el presente estudio se evidenció que las zonas más accesibles fueron las que concentraron la mayor cantidad de localidades, y que estas no reflejan la variabilidad ambiental a la que puede estar sujeta una especie. La ventaja logística de realizar recolectas en zonas cercanas a centros urbanos o vías de comunicación es que los costos de operación se reducen y se cubre una mayor área. El sesgo asociado a las áreas urbanas o vías de comunicación resultó independiente del taxón analizado, lo cual también se ha registrado en otras regiones; es decir, es un sesgo recurrente en las recolectas históricas (Sastre y Lobo, 2009; Soares-Filho et al., 2004; Soberón et al., 2000).
Las áreas que presentan un atractivo científico suelen ser mejor evaluadas que las menos atractivas (Reddy y Davalos, 2003). Por ejemplo, estudios en las áreas protegidas para la conservación o centros de alta riqueza de especies tienden a recibir un mayor financiamiento para la realización de estudios que las áreas menos atractivas (Sánchez-Fernández et al., 2008). Asimismo, en las áreas con un mayor financiamiento tiende a crearse una infraestructura que facilita la realización de inventarios (Küper et al., 2006). En el caso del estado de Tamaulipas, existen 2 zonas de alta relevancia ecológica: la Reserva de la Biósfera El Cielo y la "Laguna Madre", en las cuales se ha presentado una alta recolecta. La mayoría de las recolectas de las plantas acuáticas se enfocaron al complejo de lagunas situadas en el sureste del estado de Tamaulipas, específicamente en la zona conurbada de Tampico-Madero-Altamira (Mora-Olivo y Villaseñor, 2007), debido que es considerada como una de las regiones del país con mayor riqueza de cuerpos de agua (Rzedowski, 1985).
Una forma de reducir el sesgo en las recolectas es promover el diseño de un muestreo estratificado azaroso, que permita incrementar el esfuerzo de recolecta en las condiciones ambientales pobremente evaluadas (Ahrends et al., 2011; Dennis et al., 1999; Nelson et al., 1990). A escala regional, el muestreo aleatorio estratificado es considerado como una de las mejores estrategias para obtener estimados precisos y exactos de las condiciones ambientales asociadas a la presencia de las especies (Albert et al., 2010). Es conocido que la mayoría de los estudios por medio de los cuales se obtienen registros de recolecta tienen como objetivo analizar preguntas a escala local (Mora-Olivo y Villaseñor, 2007; Mora-Olivo et al., 2008), en que las variables que influyen en la presencia-abundancia de las especies son diferentes a las variables que responden a escalas regionales o globales (Guisan y Zimmermann, 2000; Hirzel y Lay, 2008). El caso de Tamaulipas no es ajeno a este patrón; por ejemplo, el muestreo de gastrópodos en el sur de Tamaulipas siguió un diseño aleatorio por tipo de vegetación (Correa-Sandoval et al., 2012), cumpliendo el supuesto de aleatoriedad a escala local. Sin embargo, cuando estos mismos datos se utilizan para realizar estudios a escala regional, como es la generación de modelos de distribución geográfica, presentan una alta agregación espacial. La información generada para analizar interrogantes a una escala espacial no necesariamente puede utilizarse para realizar estudios a otra escala espacial, a menos que se realice un pretratamiento de estos datos (Anderson y González, Jr., 2011).
Los 3 grupos (gastrópodos, pteridofitas y plantas acuáticas) presentaron una alta agregación espacial, aun cuando son especies que se asocian a condiciones ambientales distintas, resultado que puede deberse a que se siguió una estrategia de recolecta similar. Con frecuencia se tiende a evaluar los sitios donde ya se tienen registros de la especie, como es el caso de las localidades tipo, mientras que otras áreas no son visitadas, por lo que quedan fuera del muestreo otras condiciones ambientales en las que vive la especie. Para evitar este sesgo, es necesario reorientar la estrategia de recolecta y darle mayor peso a las áreas con menor cantidad de registros (Parnell et al., 2003).
Las especies, aun cuando pertenecen al mismo género, tienden a responder de forma diferencial a las mismas presiones ambientales (Hirzel y Lay, 2008). Los resultados obtenidos son consistentes con lo que se conoce de estos grupos; por ejemplo, las pteridofitas prefieren zonas frescas con coberturas vegetales dominadas por árboles de hojas anchas y con menor presencia de herbáceas. Esto se entiende, en buena medida, tomando en cuenta que los helechos y plantas afines prosperan mejor en condiciones sombrías y más húmedas (Mickel y Smith, 2004). Por el contrario, las plantas acuáticas tuvieron recolectas más frecuentes en zonas más cálidas, abiertas con mayor proporción de herbáceas y menos árboles de hojas anchas. Esto también refleja el carácter heliófilo de una gran mayoría de las hidrófitas, que ocupan grandes extensiones en aguas abiertas de lagunas y otros cuerpos de agua (Sculthorpe, 1985).
Los resultados pueden ser afectados por la distancia mínima entre los puntos generados al azar que fue de 10 m, ya que los puntos no fueron generados totalmente de forma aleatoria. Sin embargo, debido a que la extensión en la que se realizó el estudio fue de aproximadamente 7,806.000 ha, y tomando en cuenta que el área de un radio de 10 m es de 0.031 ha, consideramos que es una distancia que no alteraría la tendencia espacial que se encontró. Posiblemente se tendrían puntos a una distancia menor de 10 m, pero no sería la constante, ya que dada la distancia al N (600 km) y al E (260 km) sería altamente improbable que se generasen muchos puntos al azar con valores similares. Por otro lado, debido a que la mayoría de los registros de recolecta de las especies analizadas se obtuvieron antes del uso generalizado del GPS, el error asociado a la posición geográfica real de recolecta con respecto a lo registrado en las bases de registros puede ser mayor de 10 m. Tal error se debe a que solo se presentan descripciones textuales aproximadas del sitio de recolecta, por ejemplo, "a 4.5 km de NE de Ciudad Victoria" o "Desviación a Estación Velasco, carretera Ébano-Tamuín".
Analizar el grado de independencia, aleatoriedad y representatividad de los registros de recolecta para llevar a cabo estudios a escala regional, como en la generación de los modelos de distribución geográfica potencial, tiene gran relevancia en el uso, manejo y conservación de los recursos naturales (Albert et al., 2010). La tendencia actual es utilizar estos registros para generar modelos de distribución geográfica potencial que, al combinarlos, permitan llenar los huecos de conocimiento de la diversidad y priorizar los esfuerzos de conservación (Faith et al., 2013). La agregación de los registros de recolecta refleja un sobremuestreo en las áreas donde la especie es más abundante o donde es más fácil su captura (Küper et al., 2006; Madin y Lyon, 2005; Romo et al., 2006). El riesgo asociado a este tipo de muestreo es que se omiten las áreas menos accesibles o en las que la especie es poco frecuente. Las especies se observan con más frecuencia en un intervalo de condiciones ambientales, y por encima y por debajo de esas condiciones la frecuencia de observación disminuye (Hirzel y Lay, 2008). Los modelos de distribución de especies tratan de identificar estas condiciones para realizar la predicción de la distribución de las especies (Guisan y Zimmermann, 2000). Sin embargo, cuando los registros son agregados esta tendencia puede enmascararse, ya que puntos cercanos tienden a presentar valores similares (autocorrelación positiva). El resultado es que la predicción será sesgada a las características de la mayoría de los registros, que no necesariamente reflejarán las condiciones más óptimas para la especie (Dennis et al., 1999).
Entre las aplicaciones de los modelos de distribución de especies destacan el reconocimiento de las áreas prioritarias para la conservación de las especies que se encuentran fuera de las áreas naturales protegidas (Conabio-Conanp-TNC-Pronatura-FCF-UANL, 2007), la predicción de la riqueza de especies debido a escenarios de cambio climático (Diniz-Filho et al., 2009; Peterson et al., 2002), la delimitación de corredores biológicos (Munguía, Peterson y Sánchez-Cordero, 2008) o la predicción de la invasión de especies exóticas (Kearney et al., 2008). Esto significa que si la tendencia agregada de los registros, resultado de este estudio, se repite en otros taxa , debemos tener precaución en el uso de los modelos de distribución obtenidos con registros sesgados espacialmente (Botts et al., 2011). También se han detectado sesgos espaciales de los registros de recolecta en especies de plantas del Amazonas y los Andes (Feeley y Silman, 2011), especies de ranas en Sudáfrica (Botts et al., 2011) o de libélulas en España (Romo et al., 2006).
En la literatura existe una amplia variedad de métodos para reducir la colinealidad, redundancia de información y autocorrelación espacial (Feeley y Silman, 2011). Sin embargo, se presta poca atención a analizar la aleatoriedad de la posición de los registros y el sesgo al que se incurre al utilizar registros obtenidos en sitios cercanos a áreas urbanas o vías de comunicación. A pesar de los sesgos en los insumos básicos para la generación de modelos de distribución geográfica, estos continúan siendo una buena aproximación para definir estrategias y políticas públicas que promuevan la conservación, uso y manejo de los recursos naturales (Zimmermann, Edwards, Graham, Pearman y Svenning, 2010). Esto significa que es prioritario poner atención en la no aleatoriedad, no independencia y no representatividad de los registros de recolecta que se utilicen para fines de modelación. Por último, los resultados presentados sugieren la necesidad de modificar las estrategias de muestreo con el objetivo de reducir tales sesgos espaciales para tener conclusiones y propuestas más contundentes y robustas.
Agradecimientos
El primer autor agradece el apoyo financiero otorgado por el Programa de Mejoramiento del Profesorado (Promep) a la incorporación de nuevos profesores de tiempo completo en el Instituto Tecnológico de Ciudad Victoria (ITCVIC-PTC-003). Asimismo, los autores agradecen a 2 revisores anónimos por los atinados comentarios y sugerencias que mejoraron el presente documento.

