











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El cacao (Theobroma cacao L., Magnoliophyta: Malvacea) es un árbol originario de Sudamérica, cultivado en Mesoamérica desde épocas prehispánicas y que actualmente también se cultiva en África Occidental y Asia. Es un cultivo perenne de zonas tropicales de gran importancia para la generación de productos y servicios ambientales (Rice y Greenberg, 2000). Los cacaotales tienen una complejidad estructural elevada, debido a prácticas de cultivo y a la presencia de relictos de vegetación original arbórea y de otros árboles nativos y exóticos usados a manera de sombreado para el cultivo (Salgado-Mora, Ibarra-Núñez y Macías-Sámano, 2007). Existe evidencia de que los cacaotales tienen funciones similares a los bosques húmedos tropicales a los que han sustituido después del cambio de uso de suelo (Rice y Greenberg, 2000). Las características agroecológicas de los cacaotales exhiben un gradiente de intensidad de manejo en el que es posible distinguir cacaotales con manejo tecnificado, orgánico, tradicional, y prácticamente abandonados (Rice y Greenberg, 2000). La complejidad ecológica y de manejo de las plantaciones de cacao han propiciado el interés por conocer la diversidad biológica de la flora y fauna asociada, y los fenómenos ecológicos que regulan la distribución y abundancia de los taxones asociados a estos agroecosistemas (Faria, Barradas-Paciencia, Dixo, Laps y Baumgarten, 2007).
Las arañas (Arachnida, Araneae) son importantes por su acción depredadora y antagonista de insectos fitófagos en los cultivos (Maloney, Drummond y Alford, 2003; Riechert y Bishop, 1990; Sunderland, 1999). A pesar de ser reconocidas como enemigas naturales de las plagas, hay pocos estudios enfocados en conocer la riqueza y/o la ecología de este grupo en los cacaotales de otras regiones del mundo (Korinus, 2007; Stenchly, Clough, Buchori y Tscharntke, 2011). En México, la riqueza y diversidad de arañas de cacaotales ha sido estudiada por Ibarra-Núñez, Moreno-Molina, Ruiz-Colmenares, Trujillo-Olivera y García-Ballinas (2004); Moreno-Mendoza, Ibarra-Núñez, Chamé-Vázquez y Valle-Mora (2012); Moreno-Molina, Ibarra-Núñez y García-Ballinas (2001); Pérez-De la Cruz y De la Cruz-Pérez (2003); Pérez-De la Cruz, Sánchez-Soto, Ortiz-García, Zapata-Mata y De la Cruz-Pérez (2007) y Trujillo-Olivera (2002), quienes se enfocaron en la riqueza de especies de arañas tejedoras y su actividad depredadora sobre poblaciones de insectos fitófagos, y por Ruiz-Colmenares (2004) y Ruiz-Colmenares, Ibarra-Núñez y García-Ballinas (2001) para las arañas del suelo de un cacaotal de Chiapas. Debido al énfasis puesto en conocer a los grupos de arañas tejedoras, solo algunos estudios abarcan parcialmente a los grupos de arañas errantes en cacaotales (Moreno-Molina et al., 2001; Ruiz-Colmenares, 2004; Ruiz-Colmenares et al., 2001).
Los objetivos de este trabajo fueron determinar y analizar las diferencias en composición, riqueza de especies y diversidad, así como su correspondiente variación estacional de los ensambles de arañas arborícolas de 2 cacaotales con diferente tipo de manejo agronómico (tradicional y tecnificado), ubicados en la región del Soconusco, en Chiapas, México.
Materiales y métodos
Área de estudio
Los sitios de estudio se ubican en la región del Soconusco, Chiapas, donde el clima corresponde al tipo Aw(w)ig, cálido húmedo tropical con lluvias en verano (Inegi, 2006), su temperatura media anual es de 26.8 °C, el promedio máximo mensual es de 36.4 °C, el promedio mínimo mensual es de 20 °C y la precipitación media anual es de 2 201 mm (Baumann, 1999). El tipo de suelo es andosol fluvisol y la vegetación original era selva mediana perennifolia, aunque actualmente el uso de la tierra es agrícola de temporal (Inegi, 2006). Los sitios están separados entre sí por 17 km, y difieren en el tipo de manejo agronómico. El Rancho San Antonio (municipio de Huehuetán, 14°59'53.82" N, 92°26'25.23" O, 50 m de altitud), con manejo tecnificado: con canales de riego para reducir el déficit de agua durante los meses secos; podas periódicas (anuales) a los árboles de cacao, para mantener su altura reducida entre 3 y 4 m, con el fin de facilitar la cosecha de frutos; y aplicación de algunos agroquímicos para el control de enfermedades y plagas, además de cal como agente desinfectante al momento de realizar las podas. El segundo sitio forma parte del ejido Hidalgo (municipio de Tapachula, 14°48'55.07" N, 92°21'11.13" O, 30 m de altitud), con manejo tradicional: sin canales de riego, podas irregulares y sin aplicación de agroquímicos, ni de cal (Fig. 1).

Trabajo de campo
Las recolectas se llevaron a cabo en febrero, marzo y abril (época seca) y julio, agosto y septiembre (época de lluvias) de 2008. Entre esos 2 periodos se presenta una fuerte variación de las lluvias, ya que de febrero a abril hay un promedio histórico de precipitación de 126.8 mm (para el conjunto de esos 3 meses), y de julio a septiembre el promedio histórico de precipitación es de 1 102.5 mm (para el conjunto de esos 3 meses), mientras que los promedios históricos de las temperaturas medias mensuales varían solo entre los 26.4 y los 28.6 °C a lo largo del año (Inegi, 2006). En cada una de las localidades se delimitaron 3 parcelas contiguas de 50×50 m. Dentro de las parcelas se consideraron todos los árboles de cacao con un diámetro a la altura del pecho (DAP) mayor a 5 cm, marcándolos con una cinta plástica y asignándoles una clave única con numeración consecutiva para cada sitio, no por parcela. La muestra de estudio original incluyó 48 árboles por sitio (seleccionados al azar mediante el método de "muestreo aleatorio simple" de PopTools 3.0.6; Hood, 2008), pero debido a alteraciones de los árboles por el manejo agronómico, la muestra se redujo a 43 árboles por sitio, 22 en la temporada seca y 21 en lluvias, con diferente número de árboles en cada parcela. Para las arañas se empleó el método de recolecta directa, que consiste en la localización visual de las arañas en el tronco, las ramas, el follaje y la hojarasca acumulada entre las ramas del cacao, a partir de 1 m sobre el nivel del suelo hasta la altura de la copa de cada árbol (con la ayuda de una escalera). Los muestreos fueron realizados en cada ocasión por 2 personas. Como apoyo de la búsqueda visual, se usó un atomizador para asperjar agua sobre las áreas donde las arañas tejedoras ubican normalmente sus redes, con objeto de hacer a estas más visibles y facilitar su localización. Las arañas capturadas se depositaron en frascos con etanol al 75%, con sus respectivos datos de colecta.
Identificación
Para la identificación de las especies se usaron las claves taxonómicas de Ubick, Paquin, Cushing y Roth (2005), así como otras de literatura especializada, y también se compararon con ejemplares de la Colección de Arácnidos de la Unidad Tapachula (ECOTAAR) de El Colegio de la Frontera Sur (ECOSUR). Siempre que fue posible, los juveniles se asignaron a una especie por comparación con ejemplares de la colección de referencia y/o por características morfológicas somáticas en común con los organismos adultos recolectados. Las arañas que no pudieron ser identificadas a nivel de especie, por ser juveniles o por no corresponder con ninguna de las especies descritas en la literatura taxonómica, fueron asignadas a morfoespecies. Los ejemplares recolectados quedaron depositados en la ECOTAAR de ECOSUR. La recolecta de las arañas se realizó al amparo de la autorización de colecta científica (oficio núm. SGPA/DGVS/03022/08) otorgada por la Dirección General de Vida Silvestre; Subsecretaría de Gestión para la Protección Ambiental, Secretaría de Medio Ambiente y Recursos Naturales.
Análisis de datos
Se emplearon los estimadores de riqueza no paramétricos ACE y Chao1, calculados con el programa EstimatesS, versión 9.1 (Colwell, 2013) con 100 aleatorizaciones para evaluar la integridad de los inventarios (completeness sensu Sorensen, Coddington y Scharff, 2002), es decir, el porcentaje de especies encontradas con relación al número de especies pronosticadas por los estimadores, por sitio, época y en conjunto. Se usó el programa Ecosim 7 (Gotelli y Entsminger, 2001) para construir curvas de rarefacción basadas en ejemplares, por sitio y época, para hacer comparaciones de la riqueza ponderadas por el esfuerzo de muestreo (número de individuos). Las curvas se hicieron con 1,000 aleatorizaciones y la comparación fue con intervalos de confianza a 95%. Se calculó la diversidad (heterogeneidad) de los ensambles mediante el índice de Shannon (H') (Moreno, 2001), y se aplicó la prueba t modificada por Hutcheson para determinar la significancia de las diferencias entre los valores de H' obtenidos (Magurran, 1988). Los valores de H' se analizaron para separar los valores correspondientes a sus 2 componentes: riqueza (ln S) y equitabilidad (ln E, donde E= eH'/S) (Magurran, 2004), con objeto de determinar su variación tanto entre sitios como entre temporadas. Para estimar la diversidad beta, entre transectos y entre sitios, por cada época y para la totalidad de los datos, sin hacer distinción de época, se emplearon 2 coeficientes de similitud: uno basado en incidencia (Chao basado en Sorensen) y el otro basado en abundancia (Bray-Curtis o Sorensen cuantitativo). Ambos coeficientes fueron calculados mediante el programa EstimateS y los valores resultantes se emplearon para determinar las similitudes entre los transectos, por época y sitio, mediante un análisis de conglomerados en el programa PAST, versión 2.17 (Hammer, Harper y Ryan, 2001).
Resultados
Se recolectaron 8 394 arañas, pertenecientes a 28 familias, 66 géneros y 89 morfoespecies, de las cuales 57 fueron identificadas plenamente, más 1 especie que corresponde a un nuevo taxón para la ciencia (Gertschosa sp.). Las familias con mayor riqueza específica para el conjunto de datos fueron Theridiidae (17 especies), Araneidae (11), Salticidae (11), Anyphaenidae (7), Uloboridae (6) y Tetragnathidae (5). En total, se encontraron 79 especies en el sitio tecnificado y 76 en el tradicional. En el sitio tecnificado las familias con mayor riqueza de especies fueron Theridiidae (15 especies), Salticidae (11) Araneidae (10), Anyphaenidae (7) y Uloboridae (6). En el sitio tradicional las familias con mayor riqueza fueron Theridiidae (16 especies), Araneidae y Anyphaenidae (7 especies cada una), Salticidae y Uloboridae (6 especies cada una). En la época de secas se registraron 63 especies en el sitio tecnificado y 65 en el sitio tradicional, mientras que en la época de lluvias, el tecnificado presentó 71 y el tradicional 62 ( Anexo ). Para el conjunto de datos las especies más abundantes fueron, en orden descendente, Uloborus segregatus , Ariston albicans , Uloborus campestratus (Uloboridae), Leucauge venusta (Tetragnathidae), Lyssomanes jemineus (Salticidae), Metagonia asintal (Pholcidae), Philoponella signatella (Uloboridae), Chrysso cambridgei , Tidarren sisyphoides (Theridiidae) y Leucauge argyra (Tetragnathidae). Para el sitio tecnificado las especies más abundantes fueron U. segregatus , A. albicans , L. venusta , U. campestratus , L. jemineus y L. argyra ; mientras que para el sitio tradicional las especies más abundantes fueron U. segregatus , A. albicans , U. campestratus , C. cambridgei y M. asintal .
Integridad de los inventarios. Los porcentajes de integridad de los 2 estimadores empleados indican que los inventarios fueron bastante completos, con más de 90% en todos los casos, excepto para el sitio tecnificado en la época de lluvias. Las curvas de acumulación de especies mostraron una eficiencia de recolecta similar entre los 2 sitios durante la época de secas (Fig. 2A), con 92.2% (ACE) y 96.4% (Chao 1) en el sitio tecnificado, así como de 91.1% (ACE) y 93.5% (Chao 1) en el sitio tradicional. Durante la época de lluvias (Fig. 2B) la eficiencia fue de 89.7% (ACE) y 89.9% (Chao1) en el sitio tecnificado y de 95.2% (ACE) y 92.2% (Chao1) en el sitio tradicional. De manera global ( Fig.3 ), la integridad del inventario del sitio tecnificado fue de 90.98% (ACE) y 95.47% (Chao1), mientras que la del sitio tradicional fue de 95.12% (ACE) y 97.03% (Chao1).
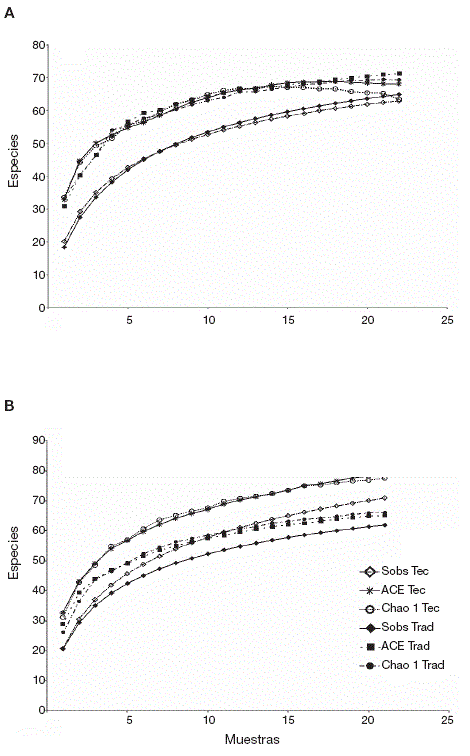
Figura 2 Curvas de acumulación de especies, basadas en la muestras de cada sitio y época. Las curvas corresponden a los valores de las especies observadas (Sobs) y a los de los estimadores Chao1 y ACE. Tec: tecnificado; Trad: tradicional. A: secas; B: lluvias.
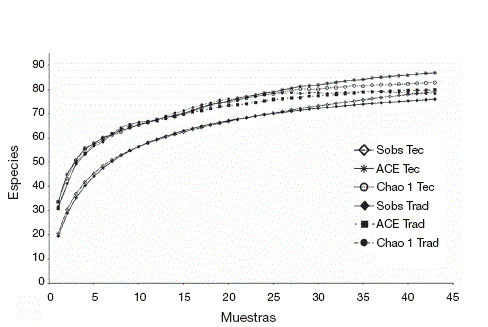
Figura 3 Curvas de acumulación de especies, basadas en el conjunto de muestras de los 2 periodos de estudio de cada sitio. Se incluyen los valores de las especies observadas (Sobs) y los de los estimadores Chao1 y ACE. Tec: tecnificado; Trad: tradicional.
Riqueza de especies
En las curvas de rarefacción para cada temporada (Figs. 4A y 4B ) al igual que para el conjunto de los datos (Fig. 5), los intervalos de confianza no se intersectaron en el punto correspondiente al sitio con menor abundancia (línea vertical), evidenciando diferencias en la riqueza, pero en sentido opuesto según la época, con una notable mayor riqueza para el sitio tecnificado en la época de lluvias y para el conjunto de datos, y una ligera mayor riqueza para el sitio tradicional solamente en la época de secas.
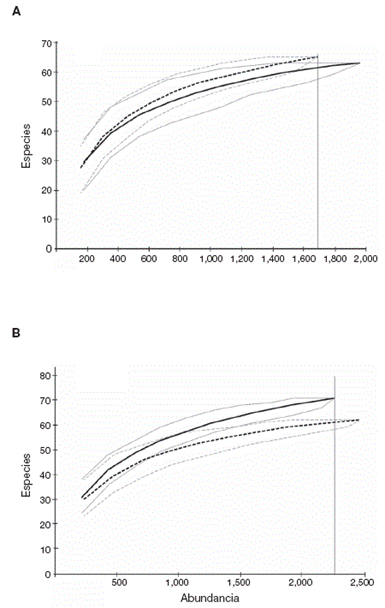
Figura 4 Curvas de rarefacción del número de individuos de cada sitio y época, con sus correspondientes intervalos de confianza de 95%. Líneas continuas: tecnificado; líneas discontinuas: tradicional. A: secas; B: lluvias.

Heterogeneidad
En todos los casos los valores del índice de Shannon fueron mayores para el sitio tecnificado y los resultados de la comparación entre sitios mediante la prueba t de Hutcheson (Tabla 1) indican que el sitio tecnificado fue significativamente más heterogéneo que el tradicional. Al disgregar los 2 componentes de la heterogeneidad (Tabla 1), se observa que en la época de secas el componente de riqueza (ln S) fue mayor para el sitio tradicional que para el tecnificado, en cambio el componente de equitabilidad (E y ln E) fue más alto en el sitio tecnificado. En la época de lluvias tanto el componente de riqueza como el de equitabilidad fueron más altos para el sitio tecnificado, y la misma relación se observa para el conjunto de los muestreos (Tabla 1).
Tabla 1 Valores del índice de Shannon (H') y su desagregación en riqueza (ln S) y equitabilidad (ln E) por sitio y época, con los valores de las comparaciones entre sitios mediante la prueba t de Hutcheson (gl: grados de libertad; * p < 0.001).
| Sitio / Época | Secas | Lluvias | Total |
|---|---|---|---|
| Tecnificado H' | 3.07 | 3.00 | 3.14 |
| Tecnificado ln S | 4.143 | 4.263 | 4.369 |
| Tecnificado E | 0.343 | 0.284 | 0.291 |
| Tecnificado ln E | -1.070 | -1.258 | -1.234 |
| Tradicional H' | 2.89 | 2.85 | 2.95 |
| Tradicional ln S | 4.174 | 4.127 | 4.331 |
| Tradicional E | 0.277 | 0.279 | 0.252 |
| Tradicional ln E | -1.284 | -1.275 | -1.378 |
| t de Hutcheson | 4.034 (gl 3 311)* | 3.850 (gl 4 726)* | 6.171 (gl 8 289)* |
Diversidad beta
Todoslos valores de los índices de similitud de Chao-Sorensen entre parcelas de ambos sitios y épocas fueron superiores al 0.90 (Tabla 2A), mientras que de los valores de los índices de similitud de Bray-Curtis el más alto fue 0.773 y la mayoría fue menor a 0.65. Los análisis de conglomerado de los valores del índice de similitud de Chao-Sorensen por parcela y época (Tabla 2A; Figs. 6CS y 6CL ) muestran una tendencia de separación entre las parcelas de cada sitio para cada época, aunque con la inclusión de una parcela del sitio tradicional, diferente en cada época, con las parcelas del sitio tecnificado. Al agrupar los datos de las 2 épocas para cada parcela, los 2 sitios se separaron claramente (Tabla 3; Fig. 6CT). Por su parte, los análisis de conglomerado de los valores del índice de Bray-Curtis por parcela y época (Tabla 2B; Fig. 7BS) indican una clara separación entre los 2 sitios en la época de secas; sin embargo, en la época de lluvias (Fig. 7BL) y para el conjunto de los muestreos (Tabla 3; Fig. 7BT), las 3 parcelas del sitio tradicional formaron un grupo que incluye además, a la parcela 3 del sitio tecnificado.
Tabla 2 Valores del índice de similitud de Chao-Sorensen (2A) y de Bray-Curtis (2B), por parcela (P1 a P3) de cada sitio (TE: sitio tecnificado, TR: sitio tradicional) y por época. Los valores por arriba de la diagonal (indicada con XX) corresponden a la época de secas y por debajo de la diagonal a la época de lluvias.

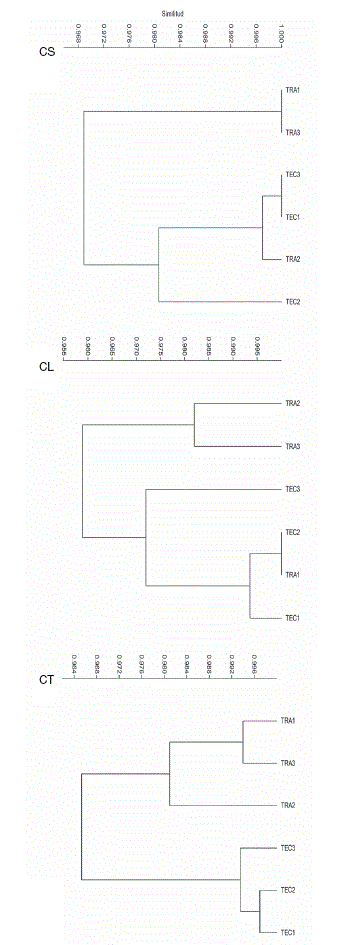
Figura 6 Análisis de conglomerados de los valores de similitud cualitativa (índice de Chao-Sorensen) entre las parcelas de ambos sitios para cada época y para el conjunto. CS, similitud para la época de secas; CL, para la época de lluvias; CT, para el conjunto de las 2 épocas.
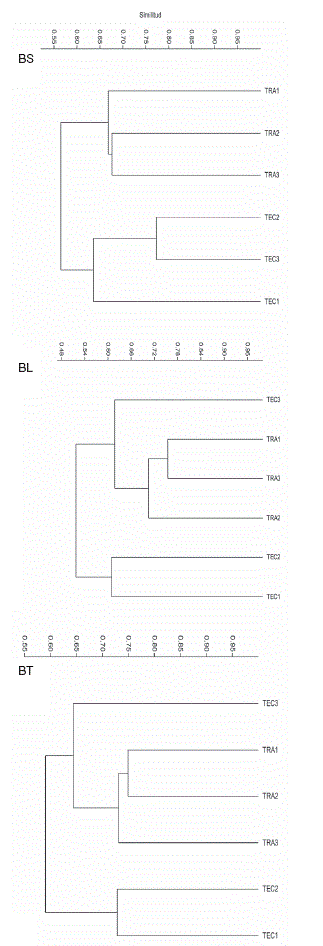
Figura 7 Análisis de conglomerados de los valores de similitud cuantitativa (índice de Bray-Curtis) entre las parcelas de ambos sitios para cada época y para el conjunto. BS, similitud para la época de secas; BL, para la época de lluvias; BT, para el conjunto de las 2 épocas.
Tabla 3 Valores del índice de similitud de Chao-Sorensen (por arriba de la diagonal, indicada con XX) y de Bray-Curtis (por debajo de la diagonal), por parcela (P1 a P3) de cada sitio (TE: sitio tecnificado; TR: sitio tradicional) para el conjunto de las 2 épocas.
| Parcelas | TE-P1 | TE-P2 | TE-P3 | TR-P1 | TR-P2 | TR-P3 |
| TE-P1 | XX | 0.997 | 0.991 | 0.965 | 0.972 | 0.957 |
| TE-P2 | 0.728 | XX | 0.996 | 0.971 | 0.971 | 0.954 |
| TE-P3 | 0.610 | 0.617 | XX | 0.965 | 0.967 | 0.966 |
| TR-P1 | 0.624 | 0.625 | 0.632 | XX | 0.982 | 0.994 |
| TR-P2 | 0.585 | 0.622 | 0.606 | 0.749 | XX | 0.980 |
| TR-P3 | 0.509 | 0.532 | 0.695 | 0.748 | 0.715 | XX |
Discusión
Las familias Theridiidae, Araneidae, Salticidae y Anyphaenidae sobresalieron en ambos sitios de estudio por su riqueza de especies, lo cual es similar a lo encontrado en el dosel de árboles de otros estudios efectuados con arañas en regiones tropicales (Álvares, Machado y Acevedo, 2004; Flores, 1999; Ibarra-Núñez, Maya-Morales y Chamé-Vázquez, 2011; Russell-Smith y Stork, 1995; Sorensen, 2004). Esto también concuerda parcialmente con un estudio sobre las 4 principales familias de arañas tejedoras en árboles de cacao en la región del Soconusco, donde Theridiidae y Araneidae son las 2 familias con mayor riqueza de especies (Ibarra-Núñez et al., 2004), y con otro estudio en Tabasco, donde las familias Salticidae, Araneidae y Theridiidae son las de mayor riqueza de especies en un cacaotal (Pérez-De la Cruz y De la Cruz-Pérez, 2003).
Los resultados de este trabajo incrementan el número de taxones de arañas registrados en México y en el estado de Chiapas, además de presentar los primeros registros de diversas especies de arañas errantes arborícolas para cacaotales de México. Entre los taxones determinados se detectaron 1 género y 9 especies como nuevos registros para el país y 1 género y 4 especies como nuevos registros para el estado de Chiapas ( Anexo ).
Para México, se registra por primera vez al género Pippuhana (Anyphaenidae) (registrado previamente en EUA, Panamá y Brasil; Brescovit, 1997) y a las especies Hamataliwa circularis (Kraus, 1955) (Oxyopidae) (en El Salvador; Brady, 1970), Castianeira dentata Chickering, 1937 (Corinnidae) (en Panamá; Reiskind, 1969), Nops largus Chickering, 1967 (Caponiidae) (en Panamá; Chickering, 1967) y cuya presencia en el país había sido supuesta a partir de ejemplares juveniles (Medina-Soriano, 2005), 4 especies del género Wulfila: W. innoxius Chickering, 1940, W. modestus Chickering, 1937, W. sublestus Chickering, 1940 y W. tantillus Chickering, 1940 (Anyphaenidae) (en Panamá; Platnick, 2013), Trachelas spinulatus F. O. P. Cambridge, 1899 (Corinnidae) (en Guatemala, Nicaragua y El Salvador; Platnick y Shadab, 1974) y Dipoena atlantica Chickering, 1943 (Theridiidae) (de Panamá hasta Paraguay; Platnick, 2013).
Como nuevos registros de arañas para el estado de Chiapas se detectaron Myrmecotipus fuliginosus O. P Cambridge, 1894 (Corinnidae) (previamente en Tabasco; Resikind, 1969), Thallumetus octomaculellus (Gertsch y Davis, 1937) (Dictynidae) (en San Luis Potosí; Gertsch y Davis, 1937). Apollophanes longipes (O. P. Cambridge, 1896) (Philodromidae) (en Guerrero; Dondale y Redner, 1975) y Goeldia tizamina (Chamberlin e Ivie, 1938) (Titanoecidae) (en Yucatán; Reddell, 1971). Del género Gertschosa (Gnaphosidae) se han descrito 4 especies, 2 para México, pero ninguna de Chiapas (Platnick y Shadab, 1981). Los organismos adultos recolectados y determinados de este género no corresponden a ninguna de las 4 especies descritas, por lo que seguramente representan una especie nueva para la ciencia.
Respecto a las especies más abundantes de cada sitio, resalta el que las especies del género Leucauge ocupen el tercero (L. venusta ) y el quinto (L. argyra ) lugares de abundancia en el sitio tecnificado, mientras que en el sitio tradicional estas 2 especies son mucho menos abundantes, ocupando, respectivamente, los sitios 12 y 18. Ambas especies tienen una amplia distribución en el continente americano y se pueden considerar como oportunistas por ser abundante en áreas naturales perturbadas, así como en varios cultivos (Dimitrov y Hormiga, 2010; Hénaut, Pablo, Ibarra-Núñez y Williams, 2001; Ibarra-Núñez y García-Ballinas, 1998; Ibarra-Núñez et al., 2004). La relativa mayor abundancia de estas especies en el sitio tecnificado pudiera estar relacionada con la mayor intensidad de manejo agronómico en ese sitio.
El nivel de integridad alcanzado en los muestreos de este estudio se puede considerar como exhaustivo (mayor al 90%), según el criterio propuesto por Cardoso (2009), y como casi exhaustivo para el único caso inferior al 90%, que fue el muestreo correspondiente al sitio tecnificado en la época de lluvias (89.7%). Estos niveles de integridad indican que los muestreos proporcionaron datos que son confiablemente representativos de los ensambles estudiados, lo que contribuye a dar confianza en los resultados de los distintos análisis realizados, apoyando a la robustez de las conclusiones que se derivan de estos.
Las curvas de rarefacción sugieren que las diferencias de manejo agronómico entre los sitios afectan la riqueza de especies encontradas tanto para el conjunto como para cada época. Sin embargo, al analizar las épocas por separado se hace evidente que las condiciones climáticas de cada época podrían haber ocasionado efectos diferentes sobre los ensambles de arañas, ya que se encontró una mayor riqueza de especies en el sitio tradicional en la época de secas y a la inversa en la época de lluvias. Al revisar los cambios en riqueza para cada sitio, en función de la época, se observa que hubo una ligera disminución de la riqueza en el sitio tradicional (de secas a lluvias), mientras que en el sitio tecnificado (y en la misma transición temporal) hubo un notable aumento de la riqueza, evidenciando que los cambios de época afectan en forma diferente a cada sitio. Las variaciones temporales en la riqueza de especies ya han sido registradas para diferentes tipos de hábitats (Lubin, 1978; Maya-Morales, Ibarra-Núñez, León-Cortés e Infante, 2012; Rubio, Corronca y Damborsky, 2008). El que estas diferencias ocurran en sentido opuesto entre los 2 sitios, parece indicar que las diferencias en manejo agronómico afectan de forma distinta su capacidad para integrar a un mayor o menor número de especies en función de las condiciones ambientales. En el caso de los sistemas manejados por el ser humano, se ha señalado que la intensidad de manejo (el grado de intervención humana a través del conjunto de prácticas agrícolas o forestales) afecta la riqueza de especies de diferentes artrópodos (Bos, Steffan-Dewenter y Tscharntke, 2007; Coyle, 1981; Delabie et al., 2007; Floren y Deeleman-Reinhold, 2005; Hore y Uniyal, 2008; Philpott y Armbrecht, 2006; Pinkus-Rendón, Ibarra-Núñez, Parra-Tabla, García-Ballinas y Hénaut, 2006; Tsai, Huang y Tso, 2006; Willett, 2001). Las prácticas agronómicas de los 2 sitios estudiados involucran diferentes niveles de intensidad de manejo en los aspectos de riego, podas y uso de agroquímicos. Trabajos futuros se deberían enfocar a determinar los efectos de las diferentes variables agronómicas sobre las arañas, lo que requiere diseñar un estudio con un registro más detallado de esas variables y de su variación temporal.
A diferencia de la variación en la riqueza, la heterogeneidad fue siempre más alta para el sitio tecnificado tanto globalmente como para cada una de las épocas. Sin embargo, al descomponer el valor de H' correspondiente, se observa que la componente de riqueza es más alta en el sitio tradicional que en el tecnificado en la época de secas, y a la inversa en la época de lluvias, así como para el conjunto, lo que coincide con los resultados obtenidos con las curvas de rarefacción. A pesar de presentar una menor riqueza en la época de secas, la mayor equitabilidad del sitio tecnificado en esa época es lo que promueve un valor más alto de H' para ese sitio, condición que posiblemente está relacionada con las diferencias en manejo agronómico, pero que con los datos disponibles no es posible determinar.
Los análisis de conglomerados de la similitud cualitativa (índice de Chao-Sorensen) de las parcelas para cada época por separado indican que los 2 sitios tienen una composición de especies parcialmente similar. La similitud en la composición de especies entre sitios puede ser derivada de la composición a nivel regional o paisajístico, donde la conectividad entre fragmentos favorece el mantenimiento de un acervo común de especies, pese a las diferencias en vegetación (Rinaldi y Trinca, 2008). En este estudio, cada cacaotal corresponde a un fragmento que forma parte de una matriz paisajística, mayormente agrícola, con la que comparte un acervo de especies, lo cual es común en zonas con cultivos perennes o en zonas de bosques en regeneración alejados de zonas con vegetación primaria (Floren y Deeleman-Reinhold, 2005). La similitud de especies entre los sitios de estudio puede verse favorecida por la dispersión de las arañas a partir de las fuentes de colonización cercanas, que son los distintos cultivos y los fragmentos de vegetación secundaria (Samu, Sunderland y Szinetár, 1999). Esta tendencia de similitud entre fragmentos ha sido observada en otros grupos de artrópodos con una alta capacidad de dispersión (Marín, León-Cortés y Stefanescu, 2009). Sin embargo, el análisis de conglomerados de la similitud cualitativa para las 2 épocas combinadas muestra una clara separación entre los sitios que parece indicar que las diferencias entre los 2 sistemas de manejo influyen sobre la composición de especies de arañas, generando 2 conjuntos globalmente diferenciados, como se observó en un estudio sobre arañas de un bosque mesófilo de montaña de esta misma región, donde la similitud (en composición de especies) entre los sitios, uno conservado y otro perturbado, fue solamente moderada, a pesar de estar separados por menos de 1 km (Maya-Morales et al., 2012). Asimismo, los resultados de los análisis de heterogeneidad y de similitud cuantitativa (medidas a través de los índices de Shannon y de Bray-Curtis, respectivamente) indican diferencias en las abundancias relativas de las especies que los integran y en consecuencia en la estructura de los ensambles de arañas. Derivado de esto, es entonces posible que las diferencias en manejo afecten su desempeño como enemigos naturales de los insectos asociados a esos cultivos, como se ha observado en cacaotales de Sulawesi, Indonesia (Klein, Steffan-Dewenter y Tscharntke, 2002). Falta por determinar cuáles son los factores que contribuyen a estas diferencias en composición y abundancia relativa. Estudios que incluyan un mayor número de cacaotales con diferentes sistemas de manejo agrícolas permitirían tener una visión a nivel de paisaje del efecto del cambio del uso del suelo en la diversidad de arañas de cacaotales.
Agradecimientos
A José Álvaro García-Ballinas por su ayuda en el trabajo de campo. A la Dra. Marisela Salgado por su apoyo en la ubicación de los sitios de trabajo; al Ing. Roberto Monterrosas, dueño del Rancho San Antonio, por las facilidades para la realización del trabajo en su propiedad. A los 2 revisores anónimos y al editor asociado por sus valiosas sugerencias al manuscrito. Al Conacyt por la beca otorgada al primer autor para sus estudios de maestría, de los cuales se deriva este trabajo.

