











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La extracción no planificada de organismos acuáticos para consumo humano ha derivado en la sobreexplotación de sus bancos naturales, la cual se incrementa anualmente debido al crecimiento poblacional (FAO, 2003). Por lo anterior y ante la creciente necesidad de productos de origen acuático, no es factible detener la extracción de materias primas que sirven para cubrir dicha demanda (Tacon, Hasan y Subasinghe, 2006). No obstante, la acuacultura es una excelente alternativa de producción de organismos acuáticos, esta es una actividad antropogénica sustentable, mediante la cual se generan diversas especies acuáticas a través de técnicas de cultivo (Castillo-Corella, Cisneros-López, Ortiz-Serrano y Jover-Cerdá, 2002). En ese sentido, se han desarrollado específicamente procedimientos de producción para especies de peces, moluscos bivalvos y algunos crustáceos (FAO, 2012). Aunque la mayor parte de ellas se enfocan en especies de importancia comercial nacional o internacional como el camarón (Litopenaeus vannamei ) y el ostión (Crassostrea gigas ) (Villarreal, 2000), dejando a un lado la tecnificación de especies nativas que no se comercializan ampliamente en el mercado, pero que registran un elevado potencial para ser cultivadas (Rodríguez-Serna, 1999). Un ejemplo es el acocil de río C. montezumae , crustáceo decápodo endémico de México perteneciente a la familia Cambaridae, localizado en la meseta central, en los estados de Hidalgo, México, Tlaxcala, Michoacán y Puebla (Villalobos, Cantú y Lira, 1993). Este acocil, independientemente de su pequeña longitud (de 30 a 50 mm), es ampliamente sobreexplotado, debido a la alta demanda de consumo alimentario por parte de los pobladores nativos de la zona, causando una merma importante en las poblaciones de C. montezumae . Además, es importante mencionar que en la actualidad sus poblaciones son afectadas también por la inserción de otras actividades antropogénicas que han llevado a la alteración de su hábitat (Álvarez y Rangel, 2007; Ángeles-Monroy et al., 2010; Latournerié-Cervera, Osorio, Cárdenas-Vázquez y Romero-Jarero, 2006; Rodríguez-Serna y Carmona-Osalde, 2002). Al respecto, Arredondo-Figueroa, Vásquez-González, Núñez-García, Barriga-Sosa y Ponce-Palafox (2011) mencionan que también la introducción de especies exóticas agresivas como la carpa común (Cyprinus carpio ) y la tilapia del género Oreochromis han mermado poblaciones de C. montezumae al consumirlo como alimento. Ante la disminución del número de individuos, su importante función en las cadenas tróficas y su valor nutrimental, se han realizado algunos estudios relacionados con su biología y ecología, los cuales dan indicios de un elevado potencial acuacultural, debido a las altas tasas de crecimiento, fácil reproducción y gran adaptación a los cambios ambientales (Latournerié-Cervera et al., 2006; Rodríguez-Serna y Carmona-Osalde, 2002). Se ha establecido que C. montezumae es un organismo capaz de vivir en ambientes reducidos y con altos contenidos de sedimentos y microorganismos, aspectos importantes en la acuacultura, debido a que la calidad química y microbiológica del agua dan la pauta para asegurar óptimas sobrevivencias y reducir los efectos negativos en la producción por desvió de energía para el proceso de adaptación a bajas condiciones fisicoquímicas del sistema (Martínez-Porchas, Martínez-Córdova y Ramos-Enríquez, 2009). Se sabe también que C. montezumae registra un elevado contenido de proteínas en su tejido somático; Sánchez-Meza, Romero, Negrete, López y Malpica (2009) mencionan que tiene un elevado porcentaje de proteínas, el cual varía dependiendo de las condiciones de cultivo y la alimentación. Ante las altas probabilidades de ser una especie cultivable en condiciones fisicoquímicas no tan exigentes y una fuente nutrimental importante para consumo humano, es necesario realizar estudios enfocados al desarrollo de una tecnificación sustentable, ya sea para su uso como materia prima de alto valor nutrimental en la elaboración de alimentos o en la obtención de crías con fines de repoblamiento de embalses y poblaciones naturales que más adelante se pudieran usar a través de pesquerías reguladas (Aguilar y Demecio, 2013; Ángeles-Monroy et al., 2010; Latournerié-Cervera et al., 2006). Así, uno de los principales retos en la tecnificación del cultivo de C. montezumae es el determinar qué tipo de alimento proporciona los requerimientos nutricionales que necesita esta especie para incrementar sus tasas de crecimiento y el valor nutrimental a nivel de tejido somático (Tacon, 1989). Lo anterior se refiere en estudios que demuestran que la calidad nutrimental del alimento influye no solo en el crecimiento y sobrevivencia, sino también en el contenido nutrimental del tejido somático de los organismos cultivados (Cerón-Ortiz, Cordero, Arredondo-Vega y Voltolina, 2009; Martínez-Córdova, Ezquerra-Brauer, Bringas-Alvarado, Aguirre-Hinojosa y Garza-Aguirre, 2002). Para esto último, se pueden aplicar alimentos utilizados en otros crustáceos de hábitos alimenticios y requerimientos nutricionales similares; aunque antes de recomendarlos para C. montezumae , tiene que comprobarse la influencia de los alimentos en su desarrollo, considerando que se localiza generalmente en el bentos y es un organismo omnívoro. El presente trabajo tiene el objetivo de conjuntar la evaluación de 2 variables básicas en la acuacultura: el tipo de alimento y la calidad de agua, sobre el crecimiento y sobrevivencia de este organismo. Además de identificar y sustentar la respuesta en el consumo y aceptación de un alimento vivo con un desplazamiento en toda la columna del agua y uno balanceado que permanece inerte en el fondo para esta especie de crustáceo; lo cual sentará las bases para trabajos ulteriores.
Materiales y métodos
Diseño experimental
El trabajo se efectuó en las instalaciones acuícolas del Centro de Estudios Tecnológicos en Aguas Continentales Núm. 02 (CETAC Núm. 02), bajo un diseño factorial completamente al azar, de 2 variables independientes: calidad de agua y tipo de alimento, y 2 dependientes: crecimiento y sobrevivencia. La calidad del agua es definida por la cantidad de sólidos suspendidos aunado al contenido y tipo de microorganismos conforme a las Normas Oficiales Mexicanas (NOM-092-SSA1 [Secretaría de Salud, 1994]; NOM-112-SSA1 [Secretaría de Salud, 1994]; NOM-116-SSA [Secretaría de Salud, 1994]; NOM-143-SSA1 [Secretaría de Salud, 1995]) (NMP Escherichia coli en 100 ml igual a 2,400). La calidad de agua baja denominada así en la presente investigación, hace referencia a la presencia de microorganismos en un número mayor al mínimo permitido que establecen las normas antes referidas y al contenido de sólidos suspendidos superior a los 500 mg/l. La calidad de agua alta indica la presencia de microorganismos en una cantidad inferior a la norma correspondiente o a la ausencia de los mismos; respecto a los sólidos suspendidos se debían de registrar por debajo de los 500 mg/l. Así, los tratamientos se identificaron como M1: calidad de agua alta y alimento vivo; M2: calidad de agua alta y alimento balanceado; M3: calidad de agua baja y alimento vivo; M4: calidad de agua baja y alimento balanceado. Los tratamientos se probaron en 3 repeticiones por cada uno, con una densidad inicial de 13 organismos por unidad experimental. Cada una de ellas consistió en un acuario de vidrio con dimensiones de 50340330 cm lleno de agua, dependiendo del tratamiento correspondiente solamente hasta un volumen de 40 l). Las postlarvas de acocil de río (C. montezumae ) se obtuvieron del stock de reproductores del CETAC Núm. 02, con un peso de 0.14 g (±0.002) y longitud de 1.49 cm (±0.07). En las unidades experimentales se colocaron plantas artificiales que fungieron como zonas de resguardo durante la muda. Los parámetros fisicoquímicos se monitorearon diariamente para mantener un registro del comportamiento de los mismos, mediante kits colorimétricos individuales para alcalinidad, dureza, amonio y oxígeno (Hanna Instruments, TX US); la temperatura se midió a través de un termómetro de vidrio de inmersión total (Taylor, TX US) y la presencia de detergente con el dispositivo desmineralizador DEMI-10 (HANNA Instruments, TX US). El agua utilizada en los tratamientos M3 y M4 se obtuvo de zonas naturales en Tezontepec de Aldama, Hidalgo, con características antes referidas de calidad de agua baja. En los tratamientos M1 y M3 se utilizó agua corriente de la llave previa filtración a través de cartuchos de 10 y 5 micras. Los datos obtenidos se analizaron en el paquete estadístico Statistica 7 (Statsoft, Austin TX US), donde se verificó su normalidad distributiva (Wilk-Shapiro) y homocedasticidad de Bartlett, posteriormente al determinar que los datos cumplieron ambos supuestos se aplicó un análisis de varianza. Al registrar diferencias significativas, se aplicó una prueba HSD (Honestly-significant-difference) de Tukey. En los datos referentes a la supervivencia, se calculó la media aritmética y la desviación estándar.
Alimento
Se evaluaron 2 tipos de alimentos: un alimento balanceado (Camaronina 35 de la marca Purina) y uno consistente en organismos vivos (D. pulex con una talla de 2 mm), los cuales se proporcionaron en 2 raciones (por la mañana y la tarde) a una tasa de alimentación por día del 15% del peso seco total (PST) de los organismos, modificándose semanalmente de acuerdo con la densidad de organismos registrada en las unidades experimentales y al incremento en peso húmedo (PHum), previa obtención al inicio del experimento de su equivalente en PST utilizando el principio del método de peso constante de Sorokin (1973). Además, el método antes referido también se aplicó antes de comenzar los bioensayos de alimentación para obtener los datos de PHum, PST y peso seco orgánico (PSO), con el propósito de ajustar en su base seca el contenido de ambos alimentos. El método consistió en obtener primeramente el PHum al pesar las muestras en una balanza analítica modelo Adventurer con precisión de 0.0001 g (Ohaus, NJ US), posteriormente se colocaron en una estufa a 60 oC y las muestras se extrajeron cada 60 min para pesarse en la misma balanza, repitiéndose este paso hasta la obtención del peso seco constante. En seguida se calcinaron en una mufla de la marca Felisa modelo FE-361 (MEX) a 490 oC por 12 h, se dejaron enfriar dentro de la misma hasta que alcanzó los 50 oC, finalmente las muestras se pesaron para la determinación de cenizas y obtener el PSO por diferencia entre el peso seco total y las cenizas.
Los bioensayos de alimentación se desarrollaron a lo largo de 14 semanas, durante este periodo se realizó un control diario en el consumo de alimento, retirando el remanente antes de agregar de nueva cuenta la ración correspondiente. El peso seco del alimento remanente se consideró para determinar la cantidad de alimento consumido por los organismos en cada tratamiento, la cuantificación se realizó cada 4 días mediante muestreos al azar en donde se aplicó el método de Sorokin (1973) antes descrito, considerando como muestra al alimento remanente para la obtención del peso seco y por relación de proporcionalidad obtener el porcentaje de consumo. El cultivo de los organismos vivos (D. pulex ) se desarrolló en estanques de concreto de 3 m3 de capacidad a temperatura ambiente y cielo abierto.
Evaluación del crecimiento
Al inicio y al final de los experimentos se sacrificó un lote de 20 organismos de C. montezumae para obtener el PHum, PST y PSO a través del método de Sorokin (1973), resultados que se consideraron a lo largo de los bioensayos para calcular la tasa de alimentación con base en el PST de los organismos. Lo anterior, debido a que la obtención del PST y PSO implican el sacrificio de organismos y esto tendría un impacto negativo en el número de animales en las unidades experimentales.
El crecimiento de las postlarvas de acocil (C. montezumae ) mantenidas en cada unidad experimental se obtuvo semanalmente al considerar el total de organismos en cada tratamiento con base en el incremento longitudinal (desde el cefalotórax hasta el telson medido con un vernier de precisión de 0-6"/150 mm ajuste fino tipo pie de rey) y PHum corporal (en una balanza analítica con las especificaciones referidas en párrafos anteriores) de los mismos con respecto al tiempo. Asimismo, se determinaron las tasas específicas de crecimiento (TEC) por semana en longitud, mediante la fórmula modificada de Hernández, Rouse, Olvera-Novoa y Davis (2003) expresada como TECL= ((LnLF-LnLI)/t)3100, donde Ln= logaritmo natural, LF= longitud final, LI= longitud inicial, t= tiempo. Respecto a la ganancia en peso reflejada en la tasa de crecimiento específico se aplicó bajo una modificación de la ecuación propuesta por Mimbela-López (2000), quedando como TECW= (LnWf-LnWi)/t)*100 donde Pg= peso ganado, Wf= peso final, Wi= peso inicial, t= tiempo.
La sobrevivencia se calculó semanalmente utilizando el dato registrado diariamente sobre organismos muertos y vivos en las unidades experimentales, al dividir el número final de organismos a la semana sobre el número inicial de los mismos, posteriormente se multiplicó por 100 (Mimbela-López, 2000).
Composición bioquímica de las dietas
La composición bioquímica del alimento inerte (Camaronina 35) se obtuvo de la etiqueta del empaque comercial (Purina, Son MEX): proteínas 35%, lípidos 8%, carbohidratos 5%, vitaminas (A, D-3, E, K, Tiamina, Riboflavina, Niacina, ácido pantoténico, piridoxina, B12, Biotina, ácido fólico, cloruro de colina, ácido ascórbico, inositol) y minerales (CaCO3, NaH2PO4 y/o Ca(H2PO4), NaCl, CoSO4, MgSO4, ZnSO4, FeSO4) en un 52%. Respecto a D. pulex , se aplicaron métodos colorimétricos para la obtención de proteínas (Lowry, Rosebrough, Lewis-Farr y Randall, 1951), carbohidratos (Dubois, Gilles, Hamilton, Rebers y Smith, 1956) y lípidos (Bligh y Dayer, 1959; Pande, Parvin-Khan y Venkitasubramanian, 1963). Para las curvas de calibración, se usaron como estándares albúmina de bovino en el caso de las proteínas, glucosa anhidra para los carbohidratos y tripalmitina para los lípidos. Asimismo, se consideraron valores promedio de diferentes autores que refieren la composición proximal de esta especie para aminoácidos (arginina 10.92%, histidina 2.69%, tirosina 4.27%, cistina 1.17%, metionina 3.45%), Ca 0.21 mg·g-1, Mg 0.12 mg·g-1, P 1.46 mg·g-1, Na 0.74 mg·g-1, K 0.72 mg·g-1, Fe 72.2 mg·g-1, Zn 12.8 mg·g-1, Mn 13.2 mg·g-1, Cu 1.2 mg·g-1 (Castro-Barrera, 2003; Muñoz-Gutiérrez, 2007; Torrentera y Tacon, 1989).
Composición bioquímica de Cambarellus montezumae
El impacto de la calidad nutrimental de los alimentos proporcionados se realizó considerando todo el cuerpo del organismo (C. montezumae ) a través de análisis proximales, por triplicado. Para lo cual, al inicio de los bioensayos se aplicó un muestreo completamente al azar de los organismos para seleccionar 3 muestras de 20 g con base en el peso húmedo para la determinación de su contenido proximal y 6 muestras de 2 g con base en el peso seco para los análisis de ácidos grasos. Finalmente, al final de los bioensayos en cada tratamiento se tomó de nueva cuenta la misma cantidad de biomasa para la aplicación de los análisis antes mencionados. Las muestras se analizaron en un laboratorio certificado con base en las normas mexicanas específicas para humedad (Nom-116-SSA1, 1994), cenizas (NMX-F-607-Normex, 2002), proteína (NMX-F-607-Normex [Secretaría de Economía, Segunda Sección, 2002]), proteína (NMX-F-608-Normex [Secretaría de Economía, 2011]), grasas (NMX-F-615-Normex [Secretaría de Economía, 2004]), fibra (NMX-F-613-Normex [Secretaría de Economía, 2003]) y carbohidratos no fibrosos por diferencia.
El contenido de ácidos grasos en el tejido de C. montezumae se realizó en el Centro de Investigaciones Biológicas del Noroeste, para lo cual, se almacenaron a -70 oC individuos de C. montezumae antes y después de los bioensayos hasta su utilización; posteriormente se liofilizaron y enviaron para análisis. La extracción de los ácidos grasos se realizó con hexano grado HPLC y la determinación por medio de cromatografía de gases-espectrometría de masas; GCMS GCD 1800B, software de integración WSEARCH, método PUFAC37 con mezcla de estándares de ácidos grasos ME: catálogo SUPELCO 47885-U.
Resultados
Durante los bioensayos se registró una estabilidad en los parámetros fisicoquímicos planteados en el proceso metodológico para cada una de las calidades de agua en las distintas unidades experimentales (Tabla 1). En donde se enmarca el mayor contenido microbiológico en los tratamientos M3 (calidad de agua baja y alimento vivo) y M4 (calidad de agua baja y alimento balanceado). En cuanto a la ingesta de alimento vivo y balanceado se observó que los acociles no consumieron la misma cantidad a lo largo del bioensayo indistintamente del tratamiento, registrando durante las primeras 3 semanas, aproximadamente el 10% de alimento remanente, posteriormente el porcentaje disminuyó conforme avanzaba el bioensayo hasta un valor cercano al 3%. Los resultados obtenidos en crecimiento (longitud y peso húmedo) registraron diferencias significativas entre tratamientos (p < 0.05) y al aplicar las pruebas de comparación múltiple se establece que es el tipo de alimento quien tiene el impacto significativo en el crecimiento, independientemente de la calidad de agua utilizada. Alcanzando los valores más altos en los tratamientos con alimento balanceado M2 y M4 respecto a los tratamientos con alimento vivo M1 y M3 con una p < 0.05 (Fig. 1).
Tabla 1 Promedio y desviación estándar (±) de los valores registrados en los diferentes parámetros fisicoquímicos y valor del contenido microbiológico en las unidades experimentales.
| Calidad de agua alta (M1 y M2) | Calidad de agua baja (M3 y M4) | |
|---|---|---|
| Temperatura | 21±1° C | 21±1° C |
| Detergentes | 0.00±0 mg.l-1 | 0.02±0.001 mg.l-1 |
| Amonio | 0.00±0 mg.l-1 | 0.03±0.009 mg.l-1 |
| Dureza | 150-160 mg.l-1 | 150-160 mg.l-1 |
| Alcalinidad | 40-80 mg.l-1 | 45-80 mg.l-1 |
| Oxígeno | 5±0.02 mg.l-1 | 5±0.04 mg.l-1 |
| Contenido microbiológico | NMP Escherichia coli en 100 ml "0" | NMP Escherichia coli en 100 ml "2 400" |
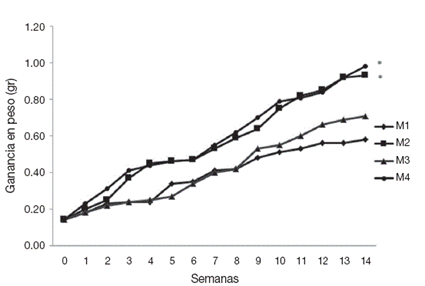
Figura 1 Tendencia en el crecimiento con base en el peso húmedo de postlarvas de Cambarellus montezumae cultivados en diferentes tratamientos. M1: calidad de agua alta y alimento vivo; M2: calidad de agua alta y alimento balanceado; M3: calidad de agua baja y alimento vivo; M4: calidad de agua baja y alimento balanceado. * indica p < 0.05 vs . M1 y M3.
Los incrementos en el peso húmedo muestran una tendencia al crecimiento discontinuo o por saltos, principalmente de la biometría 4 a la 6 y de la 11 a la 12 en los organismos de los tratamientos M2 y M4. Así como de la 2 a la 4 y de la 12 a la 13 en los tratamientos M1 y M3 (Fig. 1). Resaltando los pesos máximos en M2 y M4 a partir de la tercera biometría, alcanzando valores al final de los bioensayos de 0.93 g (±0.22) y 0.98 g (±0.15) respectivamente; con relación a M1 y M3 los valores máximos al final del experimento son de 0.58 (±0.08) y 0.71 g (±0.09). Respecto a las TECW semanal los valores registrados son de 0.102 g en M1, 0.135 g en M2, 0.116 g en M3 y 0.139 g en M4; las cuales refieren un incremento en peso del 414.28% en M1, 664.28% en M2, 507.14% en M3 y de 700% en M4 al final de los experimentos.
Se obtuvieron resultados similares en la ganancia de longitud, los cuales muestran una tendencia al crecimiento discontinuo o por saltos, principalmente en las biometrías 1, 2 y de la 11 a la 12 en los organismos de los tratamientos M2 y de la 7 a la 10 para M4. Así como de la 2 a la 4 y de la 7 a la 10 en el tratamiento M1 y de la 8 a la 12 en M3 (Fig. 2). Resaltando las longitudes máximas en M2 y M4 alcanzando valores al final del bioensayo de 2.97 (±0.48) y 2.97 cm (±0.21), respectivamente; con relación a M1 y M3 los valores máximos al final del experimento son de 2.61 (±0.31) y 2.51 cm (±0.43) respectivamente. Asimismo, se registró la TCEL semanal de 0.040 cm en M1, 0.049 cm en M2, 0.037 cm en M3 y 0.049 cm en M4; las cuales refieren un incremento en longitud del 175.16% en M1, 199.32% en M2, 188.45% en M3 y de 199.32% en M4 al final de los experimentos.
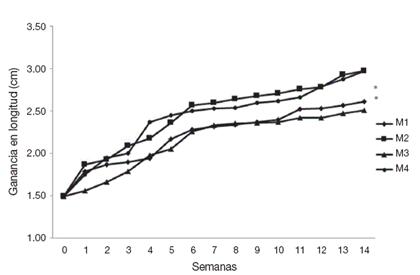
Figura 2 Tendencia en el crecimiento con base en el peso húmedo de postlarvas de Cambarellus montezumae cultivados en diferentes tratamientos. M1: calidad de agua alta y alimento vivo; M2: calidad de agua alta y alimento balanceado; M3: calidad de agua baja y alimento vivo; M4: calidad de agua baja y alimento balanceado. * indica p < 0.05 vs . M1 y M3.
El contenido de humedad en los individuos evaluados tanto al inicio como al final de los bioensayos se encuentra entre los 78.24 y el 78.48%, independientemente del tratamiento (p > 0.05). Se registraron tendencias similares en el contenido de cenizas, alcanzando un intervalo de valores de 31 al 33% en los 4 tratamientos tanto en el inicio como al final de los bioensayos, de las cuales se identificaron valores en el contenido de sodio en un intervalo de entre 8.30 y 8.60 mg/100 g.
Respecto a la modificación del contenido proximal en el tejido de C. montezumae al comparar los valores iniciales y finales, se registraron diferencias significativas entre tratamientos (p < 0.05) y al aplicar las pruebas de comparación múltiple se establece que es el tipo de alimento lo que tiene un impacto significativo en la composición bioquímica de los organismos, independientemente de la calidad de agua utilizada en los bioensayos (Tabla 2). De manera destacada se registraron incrementos en el contenido de proteínas en los organismos, independientemente del tratamiento al final de los bioensayos, registrando los mayores valores en los organismos mantenidos en los tratamientos M2 y M4 (p < 0.05). En carbohidratos solamente, los organismos mantenidos en el tratamiento M3 mantuvieron los valores iniciales y al comparar estadísticamente los datos se identificaron diferencias significativas, en donde los valores más altos se registraron en M1 y M3 (p < 0.05); de los cuales entre el 0.08 y 0.09% son fibra cruda. Tendencias similares se observaron en el contenido lipídico de los organismos, en donde el único tratamiento que registró incrementos en los valores finales fue M3 con respecto al inicio del bioensayo y al final del mismos al compararlos con los otros tratamientos (p < 0.05).
Tabla 2 Promedio y desviación estándar (±) del contenido proximal considerando todo el cuerpo del organismo (Cambarellus montezumae ) al inicio y final de los bioensayos.

Los resultados obtenidos respecto al contenido de ácidos grasos al final del experimento reflejan un incremento en los porcentajes totales de los ácidos saturados, una estabilidad en los monoinsaturados y una tendencia a la disminución en los poliinsaturados (Tabla 3). Asimismo, se registraron diferencias significativas entre los tratamientos (p < 0.05), los valores más altos del total de ácidos grasos saturados se registraron en los organismos cultivados bajo el tratamiento M3; en cuanto a los poliinstaurados en M4 y en los monoinsaturados la diferencia se establece entre M1 y M2 con respecto a M4.
Tabla 3 Promedio y desviación estándar (±) del contenido de ácidos grasos considerando todo el cuerpo del organismo (Cambarellus montezumae ) al inicio y final de los bioensayos.

De manera específica, se registraron 9 tipos de ácidos grasos saturados, cuya cantidad varió dependiendo del tipo de ácido; en donde el ácido graso saturado 16:00 tiene los máximos valores en un intervalo del 21 al 29.18% (Fig. 3A). Con relación a los ácidos grasos monoinsaturados se identificaron 7, de los cuales el 18:1 n9 c registra los valores más altos (15.25 a 24.10%) (Fig. 3B).
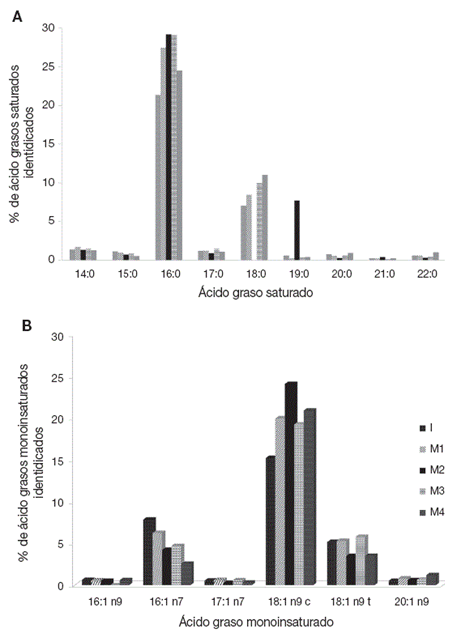
Figura 3 Porcentaje de ácidos grasos saturados identificados (A) y Porcentaje de ácidos grasos monoinsaturados identificados (B) en el acocil de río Cambarellus montezumae mantenidos en diferente tratamiento. I: contenido al inicio de los bioensayos; M1: calidad de agua alta y alimento vivo; M2: calidad de agua alta y alimento balanceado; M3: calidad de agua baja y alimento vivo; M4: calidad de agua baja y alimento balanceado.
Finalmente, se identificaron 6 ácidos grasos del tipo poliinsaturados en los organismos (Fig. 4). En donde los valores más altos se registran en los ácidos grasos 18:2 n6 c, 20:4 n6 y 20:5 n3 en los diferentes tratamientos (M1, M2, M3 y M4). Asimismo, se obtuvieron diferencias significativas entre el tipo de ácidos grasos y su contenido en los organismos mantenidos en los diferentes tratamientos (p < 0.05). Al respecto, el contenido de los ácidos grasos 18:3 n3 y 20:4 n6 fue mayor en los análisis realizados al inicio de los bioensayos (I) que al final de los mismos, independientemente de los tratamientos (M1, M2, M3 y M4). Los valores máximos de los ácidos grasos 20:5 n3 y 22:6 n3 se identificaron en los organismos mantenidos en el tratamiento M4.
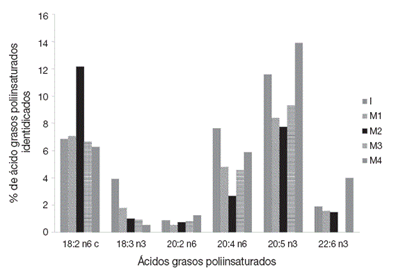
Figura 4 Porcentaje de ácidos grasos poliinsaturados identificados en el acocil de río Cambarellus montezumae mantenidos en diferente tratamiento. I: contenido al inicio de los bioensayos; M1: calidad de agua alta y alimento vivo; M2: calidad de agua alta y alimento balanceado; M3: calidad de agua baja y alimento vivo; M4: calidad de agua baja y alimento balanceado.
Los valores de supervivencia de los organismos durante los bioensayos registraron diferencias significativas entre tratamientos (p < 0.05), registrándose los más altos valores en los organismos mantenidos en el tratamiento M2 (64%) y M4 (62%) a diferencia del M1 (46%) y M3 (41%). Aunque en general, se establece una tendencia a disminuir conforme avanza el tiempo, identificándose puntos específicos de mortalidades sobresalientes, principalmente entre las semana 5 y 6 del cultivo con valores entre los 10 y 15% en M1 y M3 y de 5 a 10% en M2 y M4.
Discusión
El incremento en las medidas morfométricas de los crustáceos de -manera natural se registra de forma discontinua o por saltos a diferencia de los peces, lo anterior es debido a que en los crustáceos el crecimiento está relacionado al proceso de muda; en donde existe un cambio del exoesqueleto del organismo ante la ganancia en peso y talla, por lo cual, debe eliminar la estructura quitinosa para dar paso a una nueva con las dimensiones necesarias para cubrir el nuevo tejido somático. La generación de la nueva muda durante el periodo de crecimiento se relaciona directamente con la calidad nutrimental del alimento, las condiciones fisicoquímicas del cultivo, la etapa de desarrollo de los organismos y la aceptación del alimento por parte de los mismos (Devaraj y Natarajan, 2006; Martínez-Porchas et al., 2009). Los resultados obtenidos con C. montezumae en el presente estudio lo ubican también en las tendencias de un crecimiento discontinuo característico de los crustáceos.
En cuanto al efecto del tipo de alimento en el incremento en peso y talla, se encontró una relación con el tipo de tratamiento (M2 y M4 mayor, con respecto a M1 y M3), como pudo apreciarse en los resultados. En ese sentido, algunos estudios indican que la preferencia y aceptación del tipo de alimento, vivo o inerte, por parte de los organismos y su relación con el crecimiento y sobrevivencia depende en gran medida de su estadio de desarrollo y hábitos alimentarios (Cerón-Ortiz et al., 2009; Velasco y Lawrance, 2000). Carlos-Mallo (2005) en 2 experimentos por separado con el langostino argentino (Pleoticus muelleri , Bate) registró en el primero de ellos que al alimentar a postlarvas de PL1 a PL10 (3.9 a 4.1 mm de longitud y 1.7 a 1.8 mg de peso inicial) los mejores crecimientos y sobrevivencias se registraban en los organismos alimentados con nauplios de Artemia sp. como alimento vivo (6.6 a 7.8 mm de longitud y 3.4 a 5.7 mg de peso final). En el segundo experimento de PL11 a PL25 (7.2 a 7.3 mm de longitud y 7.3 a 7.6 mg de peso inicial) registró los mejores crecimientos al proporcionar dietas inertes microencapsuladas (8.2 a 9.9 mm de longitud y 8.0 a 10.2 mg de peso final). Al respecto, en el presente estudio se sustentan a través de una metodología científica tendencias similares en el crecimiento y sobrevivencia de postlarvas de C. montezumae , datos que no habían sido determinados en algún otro trabajo y donde los organismos alimentados con una dieta inerte (Camaronina 35) presentaron los mejores pesos, longitudes y sobrevivencias, en comparación con el alimento vivo (D. pulex ). Lo anterior es uno de los factores que era necesario probar para determinar si efectivamente como se pudiera inferir que los mejores crecimientos se registrarían con el alimento inerte por los hábitos bentónicos de las postlarvas de C. montezumae , o bien, es en conjunto con la calidad del alimento, las variables que influyen en el desarrollo de este crustáceo. Al respecto, los resultados obtenidos en condiciones controladas permiten establecer que postlarvas con pesos a partir de los 0.14 g de peso y 1.49 cm de longitud ingieren más fácilmente el alimento inerte, Camaronina 35, lo que permitió alcanzar valores de 0.98 g y 2.97 cm al final del estudio. Lo cual se relaciona también a la posición del alimento en la columna del agua (Isiordia-Pérez y Puello-Cruz, 2007), si se considera que el alimento balanceado se dirigía inmediatamente al fondo de las unidades experimentales permitiendo más fácilmente su consumo, a diferencia de D. pulex que se localizaba en su mayoría en la parte media de la columna de agua y en menor proporción nadaban cerca el fondo. Lo cual puede aseverarse con las observaciones realizadas durante los primeros 15 min posteriores a la adición de los 2 tipos de dietas y en el alimento remanente a las 24 h en las unidades experimentales correspondientes, en donde se apreció que Camaronina 35 se consumía más prontamente que la pulga de agua. Además, es importante hacer mención que, aunque es recomendado realizar la evaluación del balance energético cuando se trabaja con bioensayos de alimentación para identificar la proporción de la energía destinada por el organismo al crecimiento, respiración, excreción, e incluso a la captura del alimento (Martínez-Porchas et al., 2009). El objetivo del presente estudio se refería a detectar el efecto que el tipo de alimento y la calidad de agua tenían en el crecimiento y sobrevivencia de postlarvas de C. montezumae , obteniendo resultados relevantes. Lo anterior es importante, ya que en la actualidad en algunas de las zonas naturales en el estado de Hidalgo donde se encuentra C. montezumae , la pulga de agua y otros organismos invertebrados que le sirven de alimento, se distribuyen en toda la columna de agua, aportando elementos que indican que en un cultivo bajo condiciones controladas lo mejor es proporcionar un alimento balanceado con las características fisicoquímicas de Camaronina 35. Se requieren estudios posteriores, que como el presente, contribuyan a generar el conocimiento que hace falta específicamente para esta especie, y así, identificar la ecuación del balance energético de este individuo durante los procesos de alimentación con el alimento balanceado.
En relación con el consumo, este varió a lo largo de los bioensayos conforme se incrementaba el peso de los organismos, registrando un consumo inicial por organismo durante las primeras 3 semanas en ambos tratamientos de 0.005 g con base en su peso seco. Posteriormente, se registró una diferencia en el consumo de las 2 dietas, con valores de 0.021 g/día en los tratamientos que se adicionaba pulga de agua y de 0.040 g/día, en los cuales se les daba Camaronina 35, estos valores son menores a los 0.16 g/día de consumo individual de alimento balanceado (Camaronina 35) expresados por Arredondo-Figueroa et al. (2011) con la misma especie. Lo cual se puede explicar en el hecho de que ellos alimentaron cada tercer día y trabajaron con reproductores, a diferencia de este estudio que se realizó con postlarvas y el alimento se proporcionó diariamente. Asimismo, se observan diferencias entre los resultados obtenidos en el presente estudio y los registrados por Latournerié-Cervera et al. (2006) en crías de la misma especie, registrando mejores crecimientos en el presente estudio si se compara el incremento en el crecimiento en un tiempo específico, lo cual es debido a la diferencia en la talla inicial, el tipo de alimento proporcionado y el tiempo de duración del experimento, ya que los autores antes referidos utilizaron detritus de Egeria densa durante 6 semanas en organismos con pesos iniciales de 9.93 mg. Lo cual concuerda con lo sugerido por Mimbela-López (2000) respecto a que la diferencia entre los resultados obtenidos en distintos estudios aun con la misma especie son adjudicados a la variedad en las metodologías utilizadas en cada uno de ellos, incluyendo diferencias en la calidad nutrimental del alimento y la etapa de crecimiento de las especies estudiadas.
En otros crustáceos se han evaluado diversos tipos de alimento durante el proceso de engorda, los cuales, dependiendo de la especie y la cantidad de biomoléculas o bioelementos presentes han derivado en distintas tasas de crecimiento. En poblaciones naturales el detritus comprende entre el 13.2-21.6% del peso total del alimento ingerido, su valor nutrimental se enfoca a la proporción de carbono: nitrógeno que se registra durante su descomposición (Momot, Gowing y Jones, 1978), cuya relación de entre 17:1 o menor es indicativo de una fuente satisfactoria de alimento y que en conjunción con E. densa acelera el crecimiento en crías del acocil C. montezumae debido a la presencia de un mayor número de setas en sus apéndices bucales que incrementan la recolección de pequeñas partículas de alimento (Latournerié-Cervera et al., 2006). Si bien es cierto, el contenido de carbono y nitrógeno es importante durante la cadena trófica de los sistemas acuáticos, es bien sabido que durante la etapa de crecimiento la biomolécula utilizada por los organismos para incrementar la masa muscular es la proteína; también son aprovechadas metabólicamente en funciones estructurales, enzimas, formación de hormonas y reparación del tejido muscular (Kureshy y Davis, 2002; Lee, Cho y Kim, 2000; Lehninger, 1995). El contenido de proteínas en el alimento que se otorgará a los organismos cultivados depende de la etapa en la que se encuentren dentro del cultivo, la especie y su hábitos alimenticios (Vega-Villasante, Martínez-López, Espinosa-Chaurand, Cortés-Lara y Nolasco-Soria, 2011). Por ejemplo, los organismos herbívoros requieren una menor cantidad de proteína que los omnívoros y carnívoros, debido a que estos 2 últimos obtienen una mayor cantidad de su energía a partir de los aminoácidos (Martínez-Porchas et al., 2009). En la etapa de engorda los valores recomendados de proteína oscilan entre el 20 y 45% (Castillo-Corella et al., 2002; Muzinic et al., 2004). Sin embargo, existen estudios que registran una menor eficiencia en la asimilación de la proteína si esta se adiciona en cantidades superiores al 36%, y otros autores como Cortés (2003) mencionan que el excedente de proteína en Cherax quadricarinatus probablemente se utiliza como energía metabólica. Algo similar pudo ocurrir en el presente estudio con el acocil de río C. montezumae al consumir pulga de agua, ya que al contener valores por encima del 50% probablemente asimiló una cantidad menor y la otra parte se destinó como energía química. Resultado contrario a lo que ocurrió con los organismos alimentados con Camaronina 35 (M2 y M4), quienes registraron mejores crecimientos y no invirtieron como parte de su proceso metabólico la transformación de proteínas en energía, tal como lo demuestran los resultados, en donde los tratamientos presentaron valores más altos.
Sin embargo, no solamente la cantidad de proteínas en un alimento es importante, también se debe de considerar su calidad, la cual se relaciona con el contenido de aminoácidos esenciales que conforma la estructura primaria de todas las proteínas (Badui, 2006). Kanazawa (2000) determinó 10 aminoácidos (arginina, histidina, isoleucina, leucina, lisina, metionina, fenilalanina, treonina, triptófano y valina) considerados esenciales para los crustáceos y recomienda que no debe de faltar en el alimento. Por lo cual, no solo la cantidad, sino la calidad de las mismas son aspectos básicos en los sistemas de engorda dentro de la acuacultura; lo cual hace referencia a los resultados obtenidos en el presente estudio, en donde a pesar de ser el alimento vivo quien registró en su composición proximal la mayor cantidad de proteínas, son los organismos alimentados con la dieta balanceada Camaronina 35 los que obtuvieron el mejor crecimiento (M2 y M4, p ≤ 0.05). Indicando que probablemente este alimento contiene en mayor cantidad los aminoácidos esenciales para el crustáceo C. montezumae a diferencia de D. pulex , probablemente debido a la composición proximal de las materias primas utilizadas en la formulación de Camaronina 35 y las existentes en el alimento consumido por D. pulex . Lo cual concuerda con lo que establece D'Abramo y New (2000) respecto a que una dieta desbalanceada desde el punto de vista de la ausencia de aminoácidos esenciales, o bien, un déficit de los mismos producen un crecimiento limitado en los crustáceos.
Otra de las biomoléculas importantes en el proceso de engorda de los crustáceos son los lípidos, principalmente los que contienen en su estructura química a los ácidos grasos debido a la función estructural, por ser precursores de la actividad enzimática, la flexibilidad y permeabilidad de la membrana biológica, incluso su falta inhibe la producción de esteroles afectando el crecimiento y sobrevivencia (Company, Calduch-Giner, Pérez-Sánchez y Kaushik, 1999; Lehninger, 1995). El contenido recomendado en el alimento para crustáceos se encuentra entre el 6 y 11% (Ramírez et al., 2010; Webster, Thompson, Muzinic, Rouse y Manomaitis, 2002), intervalo en los que se encontraron dentro de las formulaciones usadas en este estudio; y donde el tratamiento M3, de acuerdo con los resultados se mantuvo en las cantidades iniciales, y en conjunto con el M1, registraron diferencias significativas respecto al alimento inerte (M2 y M4).
Dentro de la clasificación de los lípidos totales los denominados lípidos saponificables contienen como parte de su estructura química ácidos grasos, componentes nutrimentales que tienen un efecto primordial en el desarrollo de los seres vivos (Fidalgo, Cid, Torres, Sukenik y Herrero, 1998). Los ácidos grasos pueden a su vez clasificarse de acuerdo con las saturaciones del doble enlace en su cadena hidrocarbonada, en ácidos grasos saturados, monoinsaturados y poliinstaurados (Martin, 1982). Existen estudios que han demostrado que los crustáceos tienen una baja capacidad de alargar y desaturar cadenas de ácidos grasos, principalmente los altamente insaturados n-3, por lo cual, deben de proporcionarse directamente con la dieta (Kanazawa, Teshima y Ono, 1979). Los ácidos grasos saturados son utilizados como aporte energético, constituyentes de los fosfolípidos que se encuentran en las membranas celulares y algunos de ellos se asocian con el correcto funcionamiento de proteínas. Estudios en crustáceos refieren valores de 35.93 a 51.42% de ácidos grasos saturados en su composición bioquímica, los cuales son similares a los registrados para este grupo en el presente estudio con un intervalo de 36.7 a 46.8%, y que se presentaron con mayor abundancia en el tratamiento M3, en el cual se identificaron 9 diferentes. Por otra parte, los ácidos grasos monoinsaturados tienen importancia a nivel de la estructura lipídica de las membranas, principalmente en la mielina del sistema nervioso; los cuales se han registrados en crustáceos con porcentajes del 21 al 25.49, aunque en C. montezumae se determinaron porcentajes de hasta el 33.5 (Ramírez et al., 2010).
En cuanto a los ácidos grasos poliinsaturados, la demanda e importancia de ellos en crustáceos son los de la familia linolénica (20-22) quienes han producido mejores resultados en el crecimiento (Carlos-Mallo, 2005). Además de que los ácidos grasos linoleico (18:2 n3), linolénico (18:3 n3), eicosapentanoico (20:5 n3, EPA) y docosahexaenoico (22:6 n3,) son esenciales en el metabolismo de los lípidos y carbohidratos en esta clase de organismos (D'Ábramo y New 2000; González-Félix, Gatlin III, Lawrence y Pérez-Velázquez, 2002). Además, se observó que los ácidos grasos EPA y el ácido araquidónico (20:4 n6, ARA) influyen en el fortalecimiento del sistema inmunitario, lo cual ayuda a disminuir las enfermedades provocadas por estrés durante el cultivo (Koven et al., 2001; Zenteno-Savín, Cortés-Jacinto, Vázquez-Medina y Villarreal-Colmenares, 2008). Aunado a lo anterior, se sabe que el ácido graso ARA se relaciona con organismos de agua dulce como lo es C. montezumae , en los cuales actúa como precursor de eicosanoides fisiológicamente activos como las prostaglandinas utilizadas en la regulación hormonal (Won y Orth, 1994).
Además de que la carencia de los ácidos grasos linoleico (18:2 n6) y α- linolénico (18:3 n3) provocan una falta de crecimiento, lesiones cutáneas, menor pigmentación de la piel, aumento en el metabolismo basal, alteraciones en la permeabilidad de las células, trastornos en el balance de agua y aumento en la susceptibilidad a las infecciones (Torrentera y Tacon, 1989). Características que no se observaron en el presente estudio, por lo cual, y debido a la cantidad de estos ácidos grasos presentes en el cuerpo de C. montezumae se puede considerar que cubrieron las demandas necesarias en el periodo de engorda. Y aunque en las dietas proporcionadas en el estudio no se determinó el perfil de ácidos grasos al estar presentes en el tejido muscular de C. montezumae al final de los bioensayos y ser ácidos grasos esenciales, se puede inferir que se encontraban en los 2 alimentos. Lo cual, al igual que en el grupo de camarones peneidos (Jones, Yule y Holland, 1997) influyeron positivamente en el crecimiento y la sobrevivencia de C. montezumae (crustáceos de agua dulce) y su integración en el tejido somático del mismo, lo que se demuestra en los resultados obtenidos, donde en el tratamiento M4 es donde mayormente se presentan, encontrándose un total de 6 tipos diferentes. Así, los valores registrados para este grupo de ácidos grasos se encontraron por encima de los intervalos referidos para otras especies de organismos acuáticos, lo cual incrementa el valor nutrimental de esta especie como materia prima alimentaria de consumo humano, principalmente de los ácidos grasos eicosapentanoico (20:5 n3), docosahexanoico (22:6 n3) y araquidónico 20:4 n6. Asimismo, se confirma para C. montezumae lo expuesto por otros autores respecto a que en organismo acuáticos cuyo hábitat registra temperaturas templadas los valores de ácidos grasos saturados es mayor que los insaturados (Cerón-Ortiz, 2006; Flores-Vergara, Cordero-Esquivel, Cerón-Ortiz y Arredondo-Vega, 2004).
La presencia de lípidos insaponificables como el colesterol en el alimento también es importante debido a su función moderadora en los estados intermedios de fluidez de la membrana celular en presencia de los ácidos grasos poliinsaturados como el 20:4 n6 y 20:5 n3 (Parker-Barnes et al., 2000). Además, que la presencia del colesterol es básico en la producción de la hormona de la muda, evento que se produce en el cambio de exoesqueleto durante el crecimiento (Kanazawa, 2000). Finalmente, se aprecia en los resultados del presente trabajo que el contenido inicial de lípidos tiene modificaciones en todos los tratamientos al termino del experimento. En donde los organismos alimentados con M1, M2 y M4 registraron una disminución de los lípidos aproximada al 1% al final de los bioensayos y un incremento de 1% en los alimentados con M3. Lo anterior refiere el uso de esta biomolécula en algunos procesos metabólicos de C. montezumae .
Los carbohidratos son una biomolécula necesaria en la acuacultura y, a pesar de no conocer el intervalo de las cantidades recomendadas en la composición de dietas para crustáceos como sí lo existe en lípidos y proteínas (Ramírez et al., 2010), estos tienen una actividad biológica fundamental para los organismos de esta clase, ya que son la principal fuente de energía y la cual se reserva en forma de glucógeno para la síntesis de la quitina, en la formación de esteroides y ácidos grasos (D'Abramo y Sheen, 1993). La falta de valores porcentuales de carbohidratos recomendados para el crecimiento de crustáceos es debido a sus hábitos alimenticios y su capacidad para sintetizar esta biomolécula a partir de las proteínas y lípidos (gluconeogénesis) para cubrir sus demandas energéticas (Robinson y Wilson, 1985), o bien, al hecho de que algunas especies de crustáceos como L. setiferus , L. vannamei y L stylirostris utilizan directamente a las proteínas como fuente de energía para el crecimiento (Rosas et al., 2000). Aunque esta respuesta de los crustáceos a la falta de carbohidratos deriva en un decremento en la ganancia en peso, talla y sobrevivencia. Al respecto, Cuzon y Guillaume (1997) mencionan que si la energía extraída de las proteínas debido a la falta de carbohidratos es muy alta se pone en restricción el uso de las mismas en producción de tejido somático y, por ende, se limita el crecimiento; aunado a lo anterior, observaron que una dieta baja en carbohidratos podría tener efectos negativos en el catabolismo de sustratos bioquímicos, dando como resultado un bajo nivel de eficiencia proteica. Lo antes mencionado explica lo registrado en el presente estudio con C. montezumae , en donde el contenido de carbohidratos en la dieta (siendo los tratamientos M1 y M3, los que mejores resultados presentaron) se encuentra por encima de los registrado por otros autores (Ramírez et al., 2010); lo cual es un indicativo junto al incremento en crecimiento de que la cantidad de esta biomolécula fue suficiente para proporcionar la energía metabólica necesaria sin afectar su ganancia en peso y talla por el uso de otras biomoléculas. Además, al considerar el decremento en el contenido de carbohidratos y el incremento de proteínas en el cuerpo de los organismos al final del experimento comparado con los valores iniciales sustenta precisamente el uso de los mismos por el organismo permitiendo la dirección de las proteínas al crecimiento y no como fuente de energía. La diferencia entre el contenido de carbohidratos en el alimento y el cuerpo completo de C. montezumae registrada al final de los bioensayos está relacionado a la presencia del carbohidrato aminado quitina en su exoesqueleto, compuesto orgánico necesario para la generación de la muda de los crustáceos.
Los resultados antes mencionados avalan el hecho de que el contenido nutrimental del alimento proporcionado tiene una relación directa en el contenido bromatológico del tejido del animal (Cerón-Ortiz, 2006). En ese sentido, se verificó que el contenido de proteína en el tejido somático de C. montezumae aumentó al final de los bioensayos, y aunque los organismos alimentados con alimento balanceado independientemente de la calidad del agua registraron las mayores cantidades de esta biomolécula, es importante mencionar que los otros 2 tratamientos que incluían el alimento con pulga de agua también denotaron cierto incremento en las proteínas al compáralas con el valor inicial. Estas diferencias establecen la importancia de adicionar en el alimento de esta especie un contenido de proteínas de 35%, ya que, posteriormente a estos porcentajes sus niveles al final del proceso de engorda pueden ser inferiores, lo cual implicaría un gasto innecesario por parte del productor al buscar proporcionar un alimento con alto contenido de proteína. Asimismo, si se analiza el hecho de que en el presente estudio los mejores crecimientos se alcanzaron con el alimento balanceado Camaronina 35 (marca Purina), el cual contiene el menor contenido de proteínas (35%), por lo que se recomienda la realización de perfiles de aminoácidos a los 2 alimentos probados, para identificar la presencia de aminoácidos esenciales y, posteriormente, establecer el impacto real de los mismos en el mantenimiento y crecimiento de esta especie, a la par de sentar las bases para generar un alimento propio para la misma. Lo anterior, permitirá probar si es la facilidad del consumo del alimento balanceado debido a los hábitos bentónicos de C. montezumae es quien establece los mejores crecimientos, o es también influenciado por el valor proteico que registra la dieta. Además de considerar la digestibilidad de los aminoácidos presentes en el alimento de acuerdo con la materia prima que se utiliza en su elaboración (Cruz-Suárez et al., 2009; Terrazas, Civera, Ibarra y Goytortúa, 2010). Sin dejar de lado, que la sugerencia se plantea, dada la falta de investigaciones al respecto con la especie.
En cuanto a la sobrevivencia existen estudios que indican que el tipo de dieta proporcionada durante el crecimiento afecta esta variable en el cultivo, principalmente si se considera el contenido de proteína en las formulaciones. Al respecto Castillo-Corella et al. (2002) registraron las máximas tasas de supervivencia en organismos de C. quadricarinatus alimentados con dietas que contenían el 31 y 43% de proteínas. Valores que concuerdan con otros autores que referencian que las supervivencias en crustáceos son bajas por el hecho de que al momento de estar en muda los organismos tienden al canibalismo, razón por la cual, se recomienda identificar los mejores escondrijos para cada especie de crustáceo. En el presente estudio se observó un efecto del contenido nutrimental del alimento en el porcentaje de supervivencia al final del estudio, en el cual los mejores resultados se registran en los organismos alimentados con el alimento balanceado, lo cual refuerza lo mencionado anteriormente respecto a que un contenido de proteínas del 35% y un contenido de ácidos grasos poliinsaturados incrementan las tasas de supervivencia en C. montezumae .
Respecto a los factores fisicoquímicos recomendados para el cultivo de C. montezumae se han indicado intervalos de temperatura óptimos entre los 10 y los 25 oC (Latournerié-Cervera et al., 2006). Aunque Rodríguez-Serna y Carmona-Osalde (2002) mencionan que a los 17 oC se registran en esta especie la mayor tasa de ingestión y eficiencia de consumo de alimento. En ese sentido, se observó que debido a que en el presente estudio se mantuvieron los bioensayos a una temperatura de 21±1 °C dentro de los intervalos antes referidos, la temperatura no afectó negativamente los procesos metabólicos y permitió la obtención de un crecimiento favorable en las postlarvas de C. montezumae . El resto de los parámetros fisicoquímicos se monitorearon diariamente para mantenerlos en condiciones controladas de temperatura (21 oC), pH (7-8), alcalinidad (<80 mg.l-1), dureza (<150 mg.l-1), amonio (<1 mg.l-1) y oxígeno (5 mg.l-1); valores similares a los registrados por otros autores en la misma especie, quienes refieren tasas de reproducción y crecimiento positivas (Pérez-Rodríguez, Malpica-Sánchez y Arana-Magallón, 1998). Aunque es importante referenciar estudios como el de Álvarez y Rangel (2007), quienes mencionan que esta especie en el lago de Xochimilco tiene la capacidad de ambientarse a cambios drásticos o fluctuantes en los parámetros ambientales o contaminantes sin afectar sus tasas de reproducción, crecimiento y sobrevivencia.
La calidad de agua con base en el contenido de microorganismos es una variable a considerar dentro de la acuacultura, principalmente si el producto generado será puesto para consumo fresco, ya que pudiera representar un foco de infección para los consumidores. Y aunque en el presente estudio el contenido de coliformes totales en el cultivo identificado como calidad de agua baja no tuvo un efecto significativo en el crecimiento a comparación de la calidad nutrimental del alimento, sí se registraron valores por encima de las Normas Oficiales Mexicanas 092, 112, 143-SSA1, 1994, 1994 y 1995. Lo cual establece la necesidad de realizar análisis microbiológicos específicos que determinen si la venta de este organismo en fresco debe establecerse un tratamiento de conservación bajo ciertos procesos de inocuidad.

