











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The poecilostomatoid family Kelleriidae was created by Humes and Boxshall (1996) to accommodate the genus Kelleria Gurney, 1927. The genus is distinguished by the characters described by Humes and Stock (1973) but the presence of 4 discrete elements on the endopodal segment of female maxilliped is the main character to distinguish it among other lichomolgoids (Humes & Boxshall, 1996). Kelleria is the only genus known in the family Kelleriidae and was established by Gurney (1927) to accommodate 2 poecilostomatoid copepods from the Suez Canal, K. regalis Gurney and K. purpurocincta Gurney and 2 species previously assigned to the genus Pseudanthessius Claus, 1889. Kelleria is known to contain 18 species worldwide (Gómez, 2006; Humes & Stock, 1973; Kim, 2006; Walter & Boxshall, 2014). Most of these records are from Africa, Australia, and the western Pacific (Gurney, 1927; Kim, 2006, 2009; Mulyadi, 2009; Scott, 1894).
Species of this genus are marine or brackish water forms and can be associated with crinoid echinoderms or live in loose association with various burrowing benthic invertebrates. Many species have been collected from plankton samples taken in shallow coastal waters (Boxshall & Halsey, 2004; Humes & Boxshall, 1996).
In Colombia, knowledge on the composition of the Kelleriidae and other poecilostomatoid is still lacking and until now, there are no previous records of species of Kelleria from the country. A recent biological survey of the aquatic fauna of a protected coastal lagoon system of northern Colombia yielded a single female specimen of K. reducta (Gómez, 2006). This species was originally described from a single locality of the Mexican Pacific and it is currently the only species of the genus known from the Americas. The aim of this paper is to document this new record of K. reducta for Colombia, expanding its distributional range and providing complementary morphological data and comparisons with other congeners with which it has not been hitherto compared.
Material and methods
Plankton samples were taken from the Navío Quebrado Lagoon, Colombia (11°25'N, 73°5'W) from April to December 2012, mainly in the littoral areas with vegetation (macrophytes and mangrove) but also from open water in areas close to oyster banks. Water salinity was measured with a WTW 3111 conductivity meter. Water samples were collected using a bucket of 25 L both in vegetation areas and in open water. Samples were filtered with a zooplankton net (45 μm) and preserved in 70% ethanol. The sample positive for K. reducta is located over oyster banks and it was sampled in August 2012.
Copepods were sorted from the original samples and then processed for taxonomic identification. The dissected specimen and its appendages were mounted in glycerine and sealed with Canada balsam. The specimen examined was measured in lateral position, from the anterior end of the rostral area to the posterior margin of the caudal ramus. The taxonomic identification of the genus and species recorded herein followed Humes and Stock (1973) and Gómez (2006). Drawings were made with the aid of a camera lucida mounted on an Olympus BX51 compound microscope equipped with Nomarski DIC. This specimen was deposited at the Museo de Colecciones Biológicas at the Universidad del Atlántico (UARC), Colombia. The morphological terminology follows Huys and Boxshall (1991). The following abbreviations are used in the descriptive section: P1-P6= first to sixth swimming legs, EXP= exopod, ENP= endopod. The comparative analysis includes the citation of the original descriptions together with their figures.
Complementary description
Order Poecilostomatoida
Family Kelleriidae Humes and Boxshall, 1996
Genus Kelleria Gurney, 1927
Kelleria reducta Gómez, 2006
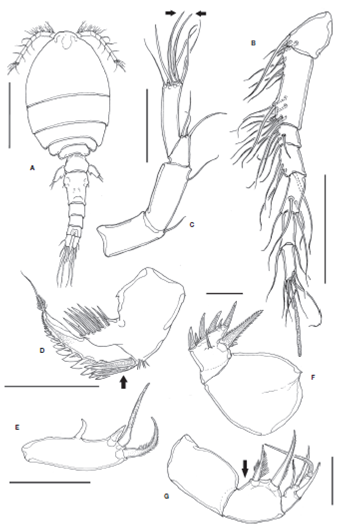
Figure 1 Kelleria reducta Gómez, 2006, adult female from Colombia. A, habitus, dorsal view; B, antennule; C, antenna (2 apical claw-like elements arrowed); D, mandible showing 2 enlarged proximal teeth (arrowed); E, maxillule; F, maxilla; G, maxilliped showing protuberant surface of basis (arrowed). Scale bars: A= 200 μm; B, C= 100 μm; D-G= 50 μm.
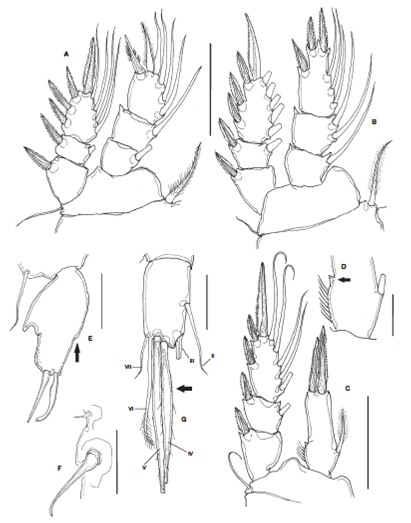
Figure 2 Kelleria reducta Gómez, 2006, adult female from Colombia. A, leg 1; B, leg 2; C, leg 4; D, Detail of spine-like process on inner margin of leg 4 endopod (arrowed); E, leg 5 showing inner process and external notch (arrowed); F, leg 6. G. caudal ramus, ventral view showing caudal setae II-VI and lightly serrate margin of caudal setae IV and V. Scale bars: A, B= 100 μm; C= 25 μm; E-G= 50 μm.
Material examined
One adult female, dissected, semi-permanent slides mounted in glycerine. UARC411Z, UARC422Z. Plankton sample, Navío Quebrado Lagoon, Colombia (11°25'N, 73°5'W), August, 2012.
Description of female from Colombia
Body cyclopiform, robust, habitus as in Figure 1A. Body length, excluding caudal setae, 980 μm. Prosome tapering posteriorly. Genital double somite, slightly tapering posteriorly. Three postgenital somites naked. Caudal rami about 2 times as long as wide with 6 caudal setae, setae IV and V with proximal expansion with serrate margins and a few scattered setules (Fig. 2G) or fully setulate margins (Gómez, 2006, Fig. 2B).
Antennule
Seven-segmented; segments 1 and 3 wider than long; segment 2 longest, about 2.4 times as long as wide; segment 4 about 1.3 times as long as wide; segments 5-7 about twice as long as wide (Fig. 1B). Armature formula as described by Gómez (2006) except for long, thick distal aesthetasc, which is shorter and slender in the Mexican population (Gómez, 2006, Fig. 4A).
Antenna
Four-segmented. Coxobasis armed with single slender seta. Endopod 3-segmented; first segment rectangular, about 2 times as long as wide, with 1 subdistal seta. Second segment shortest, with distal suture oblique, segment armed with 3 setae, one of them shorter and 1 thicker than the other 2; third segment armed with 7 elements: 5 setae and 2 claws (arrowed in Fig. 1C). Seven elements, but 3 of them being subdistal, were depicted by Gómez (2006, Fig. 4B).
Mandible
Gnathal blade broad basally, with tapering, basally pinnate apical lash. Blade with inner row of 8 strong spinules. Outer margin with robust teeth, proximal 2 teeth more than twice as long as the rest, obliquely positioned (arrow in Fig. 1D); with cluster of spinules close to base of first 2 outer teeth.
Maxillule
Represented by a single subrectangular lobe armed with 1 smooth, curved pointed element arising from middle margin and 3 distal, robust pinnate spines (Fig. 1E).
Maxilla
Large, subquadrate syncoxa, unarmed. Basis with 1 smooth robust seta inserted on middle surface reaching beyond distal margin of segment plus inner strong pinnate spine and 6 unequal distal teeth (Fig. 1F).
Maxilliped
Three-segmented, syncoxa large, unarmed; basis protuberant (Fig. 1G) or flat (Gómez, 2006, Fig. 1F), with 2 pinnate spines, proximal one shorter; single endopodal segment with 2 pinnate spines, 1 long seta, and 1 smooth, curved element.
P1-P4 ENP and EXP 3-segmented, except for 1-segmented P4ENP (Figs. 2A-C). P4ENP with small outer pointed projection (arrowed in Fig. 2D), 1 inner seta and 2 terminal pinnate setal elements, inner one being longer than outer (Fig. 2C). Armature of P1-P4 and other characters as described by Gómez (2006, Table 1).
P5 free exopodal segment oval, about 1.7 times longer than wide, ornamented with single row of minute spinules along most of outer margin and along distal half of inner margin. Segment with pointed hook-like process midlength of inner margin and notch on outer margin (arrowed in Fig. 2E), armed with 2 subequal apical elements (Fig. 2E). P6 reduced, represented by 2 setae, 1 of these very small, inconspicuous (Fig. 2F).
Remarks
According to Gómez (2006), K. reducta can be easily differentiated from its congeners by its possession of 2 mediobasal teeth of mandibular blade which are remarkably larger than the others and by a maxillipedal claw with 1 long and 1 reduced accompanying seta. These 2 distinctive characters are present in the specimen from Colombia.
Currently, this genus is represented by 18 valid species (Walter & Boxshall, 2014): K. propinqua (Scott, T., 1894), K. pectinata (Scott, A., 1909), K. regalis , K. purpurocincta Gurney, 1927, K. andamanensis Sewell, 1949, K. camortensis Sewell, 1949, K. gradata Stock, K. rubimaculata Krishnaswamy, 1952, K. australiensis Bayly, 1971, K. corioensis Arnott & McKinnon, 1981, K. vaga Kim, 2000, K. portiviva Kim, 2006, K. reducta , K. undecidentata Kim, 2006, K. grandisetiger Kim, 2006, K. indonesiana Mulyadi, 2009, K. javaensis Mulyadi, 2009, K. multiovigera Kim, 2009. Kelleria reducta is the only species of the genus that has been reported from the Americas.
The specimen from Colombia bears the diagnostic features of K. reducta as described by Gómez (2006), but some subtle difference were observed in our specimen, thus allowing an expansion of the morphological knowledge of this species: 1) the caudal rami length/width ratio; it is about 2.0 in the Colombian specimen while in the Mexican specimens the figure is about 2.5 (Gómez, 2006, Fig. 2B); 2) the ornamentation of the caudal setae IV and V differs in the Colombian specimen, margins are finely serrate along the proximal expanded section, mostly naked except for a few scattered setules (Fig. 2G), whereas these setae are regularly setulated in the Mexican specimens (Gómez, 2006, Fig. 2A, B ); 3) the basis of the maxilliped is flat in the Mexican Pacific population (Gómez, 2006, Fig. 4F) and it is clearly protuberant in the Colombian specimen (Fig. 1G); 4) the size of the female specimen from Colombia (980 μm) is outside the lowest end of the range previously known for this species (990-1100 μm) (Gómez, 2006), thus expanding the known size range of the species.
After the publication of K. reducta , several species of Kelleria were described from Korea (Kim, 2006, 2009) and Indonesia (Mulyadi, 2009); hence, these were not included in Gómez (2006). We provide a complementary comparison of K. reducta with respect to the recently described congeners. The structure of the free segment of the female P5 and the armature of the maxilliped and maxilla are variable among species of Kelleria and can are useful for separating K. reducta from its congeners: Kelleria multiovigera , K. javaensis, K . undecidentata , K. grandisetiger , and K. portiviva . In K. reducta the female P5 has a single inner pointed process (Gómez, 2006, present data), thus differing from K. javaensis , in which 2 processes occur, 1 is a small pointed process and the other is a digitiform protuberance (Mulyadi, 2009, Fig. 6H). A P5 process is absent in K . undecidentata (Kim, 2006, Fig. 4G), K. grandisetiger (Kim, 2006, Fig. 2H), K. portiviva (Kim, 2006, Fig. 6G), and K. multiovigera (Kim, 2009, Fig. 33H). Also, in both K. reducta and K. javaensis (Mulyadi, 2009, Fig. 6G) the terminal segment of the maxilliped is long and with 2 pinnate spines, 1 long seta, and 1 smooth, long curved element (Gómez, 2006, present data) while in K . undecidentata (Kim, 2006, Fig. 4B), K. grandisetiger (Kim, 2006, Fig. 2B), K. portiviva (Kim, 2006, Fig. 6B),and K. multiovigera (Kim, 2009, Fig. 33B) this segment is short, armed with 3 small and 1 smooth long setae. The second maxillary segment bears 6 distal teeth in K. reducta (Gómez, 2006, present data), thus diverging from K. javaensis (Mulyadi, 2009, Fig. 6F) and K. grandisetiger (Kim, 2006, Fig. 2A) with 8 such teeth, K . undecidentata (Kim, 2006, Fig. 4A) with 11, K. portiviva (Kim, 2006, Fig. 6A) with 10,and K. multiovigera (Kim, 2009, Fig. 33A) with 7 teeth.
Kelleria reducta most closely resembles K. indonesiana Mulyadi, 2009 in the armature formula of P1-P4 and the armature and structure of the maxilla, but these species can be separated by the following characters: 1) the last segment of the antennary ENP has 5 setae in K. reducta (Gómez, 2006, Fig. 4A, present data)and only 3 in K. indonesiana (Mulyadi, 2009, Fig. 2D); 2) the P4ENP reaches the end of P4EXP2 in K. reducta (Gómez, 2006, Fig. 6B, present data) whereas in K. indonesiana the P4ENP reaches midlength of P4EXP3 (Mulyadi, 2009, Fig. 3D); 3) the shape of P5 diverges in these 2 species, it has a single inner marginal process in K. reducta (Gómez, 2006, Fig. 3C, present data) and 2 lobes in K. indonesiana (Mulyadi, 2009, Fig. 2I); 4) the mandibular blade of K. reducta has 2 mediobasal teeth of mandibular blade much larger than the others (Gómez, 2006, Fig. 4C, present data)while in K. indonesiana only one such teeth is larger than the others (Mulyadi, 2009, Fig. 2E). Also, in K. indonesiana the row of spines on the inner margin of the blade is represented by 7 spines (vs. 8 in K. reducta ) and 4 of them are clearly thicker and longer than the other 3 (Mulyadi, 2009, Fig.2E ), thus contrasting with the subequal condition of these spines in K. reducta (Figs. 1D, 4C; Gómez, 2006).
Distribution and ecology
Hitherto, this species has been reported only from an estuarine system in the state of Sinaloa, in the Mexican Pacific (Gómez, 2006). This record from Colombia allows a significant expansion of the known distributional range of this species, from the eastern Pacific to the western Atlantic. This is also the first record of both the genus and the species from Colombia and from the Caribbean basin. In the surveyed area K. reducta was found in the plankton of the limnetic area where salinity was highest (28 psu). This value is within the range reported by Gómez (2006) for this species. The Sinaloa population was recorded from sieved sediment samples (Gómez, 2006), thus contrasting with the finding of our specimen in the water column.

