










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de biodiversidad
versão On-line ISSN 2007-8706versão impressa ISSN 1870-3453
Rev. Mex. Biodiv. vol.85 no.4 México Dez. 2014
https://doi.org/10.7550/rmb.42105
Ecología
Estructura de la comunidad de peces en cuatro estuarios del Pacífico mexicano central
Fish community structure in four estuaries of the Central Mexican Pacific
Edgar Raúl Sandoval-Huerta1*, Xavier Madrigal-Guridi1, Luis H. Escalera-Vázquez2, Martina Medina-Nava1 y Omar Domínguez-Domínguez1
1 Laboratorio de Biología Acuática, Facultad de Biología, Universidad Michoacana de San Nicolás de Hidalgo. Edificio "R" planta baja, Ciudad Universitaria 58030, Morelia, Michoacán, México. * edgar_sh50@hotmail.com
2 Laboratorio de Vida Silvestre, Área de Ecología Acuática, Centro de Estudios de Desarrollo Sustentable y Aprovechamiento de la Vida Silvestre, Universidad Autónoma de Campeche. Av. Agustín Melgar S/N, Col. Buenavista, 24039 San Francisco de Campeche, Campeche, México.
Recibido: 27 septiembre 2013
Aceptado: 20 agosto 2014
Resumen
Se analizó la estructura de la comunidad de peces y se determinaron algunos parámetros fisicoquímicos durante 4 colectas nocturnas en la desembocadura de los estuarios de barra de Nexpa, Teolán, Mexcalhuacán y barra de Pichi, Michoacán, México. Se capturaron un total de 2 014 individuos con un peso de 10 393.42 g, pertenecientes a 31 especies y 20 familias. Barra de Nexpa registró la mayor riqueza con S= 21 y el mayor porcentaje de especies raras con 22.6%. La especies con mayor abundancia fueron Agonostomus monticola y Mugil curema, mientras que Dormitator latifrons y Lutjanus novemfasciatus registraron la mayor biomasa. La salinidad, temperatura, oxígeno disuelto y transparencia mostraron diferencias significativas por temporadas, influidas por la variación estacional de la precipitación. La composición y especies dominantes fueron distintas en cada sitio y en cada temporada, mientras que la abundancia, biomasa y riqueza no mostraron variación significativa. La comunidad de peces estuvo compuesta por especies marinas y dulceacuícolas que utilizan los estuarios como zonas de crianza, especies marinas que se integran al estuario de manera ocasional y especies estuarinas que forman parte de estos ecosistemas durante la mayor parte de su ciclo de vida.
Palabras clave: ecosistemas estuarinos, ictiofauna, variación temporal, Michoacán.
Abstract
Community structure of estuarine fish fauna were analyzed in estuaries of Barra de Nexpa, Teolán, Mexcalhuacán and Barra de Pichi, Michoacán, Mexico for four night collections at the mouth of every estuary. Also, some physicochemical parameters were determined. Were captured a total of 2 014 individuals with a weight of 10 393.42 g, belonging to 31 species and 20 families. Barra de Nexpa recorded the highest richness (S= 21) and the highest percentage of rare species (22.6%). The most abundant species were Agonostomus monticola and Mugil curema while Lutjanus novemfasciatus and Dormitator latifrons recorded the highest biomass. Salinity, temperature, dissolved oxygen and transparency, showed significant seasonal differences influenced by seasonal variation in precipitation. The composition and dominant species were different at each site and in each season while the abundance, biomass and richness showed no significant variation. The fish community was composed of marine and freshwater species that use estuaries as nursery areas, marine species that are incorporated into the estuaries occasionally and estuarine species that are part of these ecosystems for most of its life cycle.
Key words: estuarine ecosystems, fish fauna, seasonal variation, Michoacán.
Introducción
Los estuarios constituyen la desembocadura de los ríos hacia el océano, representando la transición del ambiente dulceacuícola al marino (Whitfield, 1999). A diferencia de otros ecosistemas estuarinos; por ejemplo, esteros, lagunas costeras, marismas, la salinidad de los estuarios es diluida en gran medida por el agua dulce aportada por los ríos (Pritchard, 1967; Yáñez-Arancibia et al., 1994); por lo que se describen como ecosistemas con características particulares tanto fisicoquímicas como biológicas, con una composición de especies que no se encuentra en ningún otro ambiente, marino o dulceacuícola (Ray, 2005; Elliott y Whitfield, 2011). Otra particularidad, es que debido al arrastre de sedimentos ricos en nutrientes que se depositan en el cuerpo de agua, la productividad primaria de los estuarios es muy alta durante la estación lluviosa (Pérez-Ruzafa et al., 2007), lo que implica una gran cantidad de recursos en un área protegida de potenciales depredadores. Por esta razón, los estuarios son utilizados como zonas de crianza para larvas y juveniles de un gran número de especies (Aceves-Medina et al., 2008; Woodland et al., 2012).
En los estuarios, los peces son una de las comunidades más importantes, ya que pueden determinar la composición, abundancia y distribución espacio-temporal de otras comunidades bióticas; por ejemplo, al reducir la densidad de zooplancton y fitoplancton (Borges et al., 2010). Muchas especies de peces dependen de los estuarios en alguna etapa de su vida, ya que funcionan como zonas de reclutamiento, reproducción y alimentación, principalmente para organismos de origen marino, por lo que estos ecosistemas son de gran valor para el mantenimiento de las pesquerías locales y regionales (Loneragan, 1999; Kimmerer, 2002).
La distribución espacio-temporal de peces adultos y juveniles está fuertemente influenciada por los cambios en los factores ambientales (como temperatura y salinidad; Day y Yáñez-Arancibia, 1985; Laroche et al., 1997). En los estuarios tropicales, la variación estacional está determinada principalmente por la variación en la precipitación y el régimen de las mareas, por lo que la dinámica en la estructura de la comunidad de peces está altamente relacionada con dicha variación (Lyons y Schneider, 1990; Torres-Orozco, 1994). Por otra parte, cada estuario presenta características morfológicas particulares como tipo de sustrato, distancia hacia el océano, pendiente, etc., factores que también influyen en la estructura de las comunidades de peces en una escala espacial (Álvarez-Arellano y Gaitán-Morán, 1994; Pérez-Ruzafa et al., 2007).
En México se han realizado un gran número de estudios enfocados a describir la estructura de la comunidad de peces estuarinos (Ayala-Pérez et al., 2003; Castillo-Rivera et al., 2005; González-Acosta et al., 2005; entre otros). Sin embargo, en el estado de Michoacán, estos estudios son escasos, considerando que en la costa michoacana desembocan más de 53 ríos y arroyos (Correa y Ayala, 2003). En Michoacán, la zona litoral cubre una superficie aproximada de 700 km2 (Garduño et al., 2003), presenta una topografía accidentada, una franja de costa angosta y los ecosistemas estuarinos de mayor tamaño se encuentran en la región sureste del litoral (Antaramián y Correa, 2003; Correa y Gómez, 2003). Esta región corresponde al municipio de Lázaro Cárdenas, el cual alberga 83% de la población humana que habita en la zona costera (Alcalá et al., 2003). Por consecuencia, los ecosistemas costeros de la región han sufrido un gran impacto debido al drenaje de descargas domésticas no tratadas y residuos producidos por las actividades mineras y agrícolas (Toledo y Bozada, 2002).
A pesar de la importancia que estos ecosistemas y su fauna habitante representan para los pobladores, existen pocos trabajos publicados tendientes a conocer la riqueza y abundancia de los peces que habitan los estuarios de la costa michoacana (Raz-Guzmán y Huidobro, 2002). Además, en la mayoría de los casos los estudios realizados carecen de un enfoque que integre la relación entre los factores ambientales y la comunidad íctica (Madrigal, 2006). Asimismo, desde el punto de vista social y productivo, es primordial evaluar los recursos pesqueros que existen en el estado, para generar proyectos sustentables sobre su aprovechamiento. Debido a lo anterior, es necesario realizar investigaciones orientadas al conocimiento sobre las comunidades de peces estuarinos y su relación con los factores ambientales. El presente trabajo tiene como objetivo evaluar la relación de la estructura de la comunidad íctica con la dinámica estacional, así como con las características ambientales, morfológicas y espaciales de cada estuario.
Materiales y métodos
Área de estudio. Se muestrearon 4 estuarios ubicados en el municipio de Lázaro Cárdenas en Michoacán: barra de Nexpa, Teolán, Mexcalhuacán y barra de Pichi (Fig. 1).
Esta zona del Pacífico mexicano presenta un régimen micromareal de tipo mixto semidiurno (Klein, 1985), con un intervalo entre las mareas alta y baja de 0 a 2 m (Toledo y Bozada, 2002; Correa y Gómez, 2003).
De acuerdo con el sistema de Köppen (1948) modificado por García (1981), la planicie costera de Michoacán presenta un clima Awi: tropical lluvioso, con lluvias en verano, temperatura media anual de 26.5° C con máxima de 39.5° C y mínima de 9.5° C. La precipitación media anual es de 1 295.3 mm, la época de lluvias corresponde a los meses de mayo a octubre, siendo agosto el mes con mayor precipitación (Correa y Gómez 2003).
Toma de muestras y recolecta. Debido a que durante la noche la comunidad de peces estuarinos presenta una mayor actividad (Castillo-Rivera et al., 2005), se realizaron 4 muestreos nocturnos, entre las 20:00 y 22:00 h, en las bocas de cada estuario en los meses de junio, septiembre, diciembre (2010) y febrero (2011). En cada sitio se registró la salinidad (ppm) con un refractómetro de mano Vital Sine SR-6 (Pentair); la temperatura (° C) y el oxígeno disuelto (mg/L) se midieron con una sonda multiparamétrica modelo 550ADO (YSI); la profundidad (m) fue medida con una sondaleza convencional, y la transparencia (%) se determinó mediante un disco de Secchi auxiliados por una linterna de mano de 30 W. En cada sitio, los peces se capturaron con una red de arrastre tipo chinchorro (10x2.5 m) con una luz de malla de 0.7 cm, realizándose 2 arrastres en cada colecta con recorridos lineales de 50 m. Los peces fueron sedados con aceite de clavo diluido al 10% en etanol y sacrificados inmediatamente, introduciéndolos en agua con hielo. Los organismos se fijaron con formol al 5% neutralizado con borato de sodio, posteriormente se lavaron en agua y se preservaron en alcohol etílico al 70%. La identificación taxonómica se basó en las claves de Castro-Aguirre et al. (1999), Fischer et al. (1995) y Miller et al. (2005). Los especímenes fueron depositados en la colección ictiológica del Laboratorio de Biología Acuática de la Universidad Michoacana de San Nicolás de Hidalgo, con los números de catálogo: 3286-3339; 3534-3583; 3801-3838; 4906-4952.
Análisis de datos. La estructura de la comunidad de peces fue analizada entre estuarios y entre temporadas. La riqueza de especies (S) fue considerada como el número de especies total en cada estuario. Se estimó el porcentaje de especies raras por sitio mediante la relación de especies encontradas únicamente en un estuario, entre las especies totales. Se comparó la riqueza de especies por estuario mediante un análisis de rarefacción (Colwell et al., 2004), realizado en Ecosim (Gotelli y Ellison, 2013) generándose intervalos de confianza por medio de 1 000 repeticiones.
La abundancia se estimó con base en el número de individuos, mientras que para la biomasa se consideró el peso (g) total de los organismos. La abundancia relativa se determinó con base en la proporción de individuos de cada especie en relación con el total de individuos capturados en cada estuario.
Las especies dominantes en cada estuario y en cada mes, se determinaron mediante un análisis gráfico entre la abundancia relativa de cada especie y el porcentaje de frecuencia de aparición basado en la prueba de Olmstead-Tukey (Sokal y Rohlf, 1985). Este análisis permite establecer una clasificación cuantitativa de las especies en el estuario (González-Acosta et al., 2005).
Análisis estadísticos. Las variables ambientales y los valores de riqueza, abundancia y biomasa, se compararon entre estuarios y entre meses de muestreo, mediante un análisis de varianza de una vía (Anova). Los resultados con diferencias significativas, se evaluaron con una prueba post hoc de Tukey-Kramer.
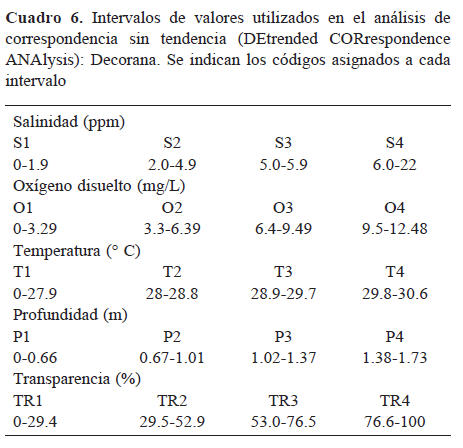
Análisis de ordenamiento. Se graficó la distribución y abundancia de especies aplicando el método de promedio recíproco, mediante el análisis de correspondencia sin tendencia Decorana (por sus siglas en inglés; Hill, 1979). El software utilizado fue Pc-ord (McCune y Mefford, 2005).
Resultados
Variables ambientales. Los valores registrados de cada variable están descritos en el cuadro 1. Se registraron valores de salinidad bajos (< 2 ppm) en todos los estuarios, excepto en junio (5-22 ppm). La salinidad más alta (22 ppm) se registró en Mexcalhuacán. El oxígeno disuelto mostró un incremento en relación con su valor inicial (x_= 3.34+2.47 mg/L) en junio, y alcanzó su valor máximo en febrero (x_= 9.03+1.75 mg/L). La concentración más alta de oxígeno (12.48 mg/L) se registró en barra de Pichi en el mes de diciembre, el valor más bajo (0.21 mg/L) se registró en junio en el mismo estuario. El intervalo de temperatura tuvo una diferencia de 3.5° C. La temperatura registró su valor más alto (30.6° C) en Teolán en junio y el más bajo (27.1° C) en barra de Nexpa en diciembre. La profundidad máxima (1.73 m) se registró en barra de Nexpa en septiembre y la mínima (0.3 m) en Teolán en junio. Los valores máximos de transparencia (100%) fueron registrados en diciembre y febrero, el porcentaje de transparencia más bajo (5.88%) se observó en barra de Nexpa en septiembre.
El Anova entre meses indicó diferencias significativas de todas las variables entre los diferentes sitios, excepto en profundidad. La prueba de Tukey-Kramer señaló que la temperatura (p= 0.037) y la salinidad (p= 0.028) fueron diferentes entre junio con respecto al resto de los meses. El oxígeno (p= 0.012) y la transparencia (p= 0.003) fueron diferentes entre diciembre y febrero en relación con junio y septiembre. Entre estuarios, se encontraron diferencias significativas únicamente para la profundidad (p= 0.017), entre Mexcalhuacán y barra de Nexpa con relación a Teolán y barra de Pichi.
Análisis de la ictiofauna. Se capturaron un total de 2 014 individuos, con un peso de 10 393.42 g. Las especies que presentaron mayor abundancia fueron Agonostomus monticola (618 individuos), Mugil curema (435 individuos), Dormitator latifrons (223 individuos) y Lile nigrofasciata (223 individuos), las cuales en conjunto representaron 74.43% de la abundancia relativa. Las especies que registraron la mayor biomasa fueron D. latifrons (4 402.18 g), Lutjanus novemfasciatus (1 428.48 g), M. curema (1 335.39 g), A. monticola (793.98 g) y Gobiomorus maculatus (725.33 g), que en conjunto representaron 83.56% de la biomasa relativa. La menor abundancia (2 individuos) se obtuvo en barra de Nexpa, mientras que la mayor abundancia (542 individuos) se registró en Teolán, ambas en el mes de septiembre. La biomasa mínima (1.34 g) se obtuvo en barra de Nexpa en septiembre, mientras que la máxima (2 639.07 g) correspondió a barra de Pichi en el mes de diciembre. Los valores totales de abundancia y biomasa por mes y sitio se describen en el cuadro 2. Entre estuarios, Teolán registró la mayor abundancia (759), mientras que barra de Pichi registró la mayor biomasa (4 992.74 g). Entre meses, septiembre fue el mes con la mayor abundancia (906) y febrero el mes con mayor biomasa (3 295.88 g). El Anova no encontró diferencias significativas para estas 2 variables.
Se identificaron 31 especies agrupadas en 28 géneros, 20 familias y 8 órdenes. Las familias con un mayor número de especies fueron Eleotridae, Gobiidae y Achiridae, cada una con 3 especies (cuadro 2). Los valores de riqueza de especies (S) por estuario y por mes, se resumen en el cuadro 3. El número total de especies registradas por estuario fue de: 21 en barra de Nexpa, 18 en Mexcalhuacán, 14 en Teolán y 12 en barra de Pichi (Cuadro 4). Junio fue el mes con el mayor número de especies (S= 22), mientras que en septiembre se registró la riqueza más baja (S= 15). El Anova no encontró diferencias significativas para la riqueza.
Barra de Nexpa fue el estuario que registró el mayor porcentaje de especies raras con 22.6%, sin embargo, fue también el único sitio donde se registró una especie exótica (Oreochromis sp.). Barra de Pichi registró un porcentaje de especies raras de 9.7%, seguido de Mexcalhuacán con 6.5% y Teolán con 3.2%.
El análisis de rarefacción indica que barra de Nexpa presenta una mayor riqueza, seguido de Mexcalhuacán, Teolán y, finalmente barra de Pichi (Fig. 2). La diferencia de número de especies fue significativa únicamente para barra de Nexpa, los intervalos de confianza (95%) en las curvas de rarefacción del resto de los sitios se sobrelapan.

Especies dominantes. El listado de especies dominantes entre sitios y meses se describe en el cuadro 5. Mugil curema fue dominante en todos los sitios y en 3 de los muestreos.
Análisis de ordenamiento. Los intervalos de variables ambientales (Cuadro 6) con mayor influencia dentro del análisis fueron: T2, T4 O3, TR1, S4 y P1. Se identificaron 4 agrupamientos principales (Cuadro 7). El Grupo I está determinado por P3 y P4; el Grupo II está definido por TR2, S2, S3, P2, T3 y O1; el Grupo III por T1, S1, TR1 y O2 y el Grupo IV por TR3, TR4, O4 y P1 (Fig. 3). Adicionalmente se encontró una especie que no correspondió a los grupos anteriores: Achirus klunzingeri, que se relacionó en gran medida con S4.

Discusión
La profundidad fue la única variable diferente entre sitios, lo que sugiere características particulares relacionadas con factores involucrados en la morfología del estuario, las características de los sedimentos y su velocidad de asentamiento (Van Rijn, 1993; Álvarez-Arellano y Gaitán-Morán, 1994). En contraparte, la profundidad fue la única variable sin diferenciación temporal significativa, ya que probablemente no es afectada por las variaciones estacionales en una escala anual. De esta manera, si existe variación en la profundidad de manera temporal, ésta no fue significativa. De la Lanza-Espino (1998) menciona que la acumulación de sedimentos puede reducir de forma importante la profundidad en esteros y lagunas costeras de latitudes tropicales del Pacífico mexicano, donde las lluvias aumentan la acumulación de sedimentos. Sin embargo, es posible que la tasa de deposición no sea suficiente para que exista diferencia significativa en un año, por lo que, en la presente investigación esta variable no se ve afectada de manera considerable. El incremento de la precipitación durante el mes de septiembre afectó directamente a la salinidad. Como resultado, el volumen de agua oceánica que ingresa a estos ecosistemas es diluida en gran medida por el agua dulce proveniente de la lluvia y los ríos (Valiela et al., 2012). La transparencia disminuyó también durante la temporada lluviosa, debido a las descargas fluviales que acarrean una gran cantidad de sedimentos con abundante materia orgánica (Pastor et al., 2011). La concentración de oxígeno disuelto también disminuyó durante esta estación, las bajas concentraciones pueden estar asociadas con una gran demanda de oxígeno, debido a un alto contenido de material orgánico aportado por los ríos (De la Lanza-Espino, 1994; Torres-Alvarado et al., 2006). En general, el oxígeno disuelto mostró valores mayores que los registrados en otros ecosistemas de características similares (Díaz-Ruiz et al., 2006; Mendoza et al., 2009). Sin embargo, en barra de Pichi se observaron valores muy bajos durante el mes junio (0.21 mg/L), lo que implica un impacto directo sobre la sobrevivencia de los peces. Siendo el oxígeno un elemento clave en los procesos metabólicos de los peces e invertebrados, su disminución es el factor de estrés más importante para los organismos acuáticos (Llansó, 1992). La hipoxia ocurre en concentraciones de oxigeno menores a 2 mg/L y es causante de mortalidad, cambios en el comportamiento, reducción del crecimiento, disminución de la biomasa y composición de especies (Wu, 2002; Eby y Crowder, 2004; Lui y Chen, 2012). Durante el muestreo de junio se observaron peces muertos a aproximadamente 50 m río arriba de donde se realizaron los arrastres. Una posible respuesta a la condición de hipoxia en barra de Pichi es la incorporación al estuario de un drenaje de aguas residuales de las poblaciones vecinas; por ejemplo, La Mira, 4.5 km al norte. Durante la temporada de estiaje una gran cantidad de desechos se acumulan, debido al diseño ineficiente del drenaje urbano (Morales y Pérez, 2006). Al inicio de la temporada de lluvias el incremento de la corriente arrastró los desechos hasta la costa causando eutrofización y procesos de descomposición que redujeron los niveles de oxígeno en el estuario.
La riqueza de especies reportada en otros ecosistemas estuarinos de Michoacán es muy similar a las 31 especies de este trabajo. En Santa Ana, un estuario 3 km al sureste de barra de Pichi, se registraron 24 especies (Madrigal, 2006), mientras que Raz-Guzmán y Huidobro (2002) indicaron 19 especies en la laguna de Salinas del Padre, en el extremo noroeste de la costa de Michoacán. Sin embargo, destaca la ausencia en este trabajo de algunas familias reportadas en los estudios mencionados; por ejemplo, Ariidae, Chanidae, Haemulidae, Polynemidae y Ephippididae. Con respecto a otros ecosistemas estuarinos del Pacífico mexicano tropical, se considera que la riqueza es equiparable a la de los 4 estuarios analizados. Por ejemplo, Mendoza et al. (2009) documentaron 33 especies en una laguna costera de Oaxaca, mientras que Díaz-Ruiz et al. (2006), reportaron 34 especies en 2 sistemas lagunares estuarinos del sur de Chiapas. Estos resultados coinciden con el promedio estimado para lagunas costeras del Pacífico mexicano, con un valor de 30 especies por sitio (Raz-Guzmán y Huidobro, 2002). Con respecto a la composición, destacan en estos trabajos la presencia de especies de la familia Dasyatidae (subclase: Elasmobranchii), ya que no existen registros similares en los ecosistemas estuarinos de Michoacán. En los estudios realizados en Oaxaca y Chiapas, se mencionan, además, las familias Characidae, Hemiramphidae, Albulidae y Synodontidae. La ausencia de especies de estas familias en los estuarios de Michoacán puede ser resultado de la poca heterogeneidad ambiental de cada ecosistema. Raz-Guzmán y Huidobro (2002) mencionan que las lagunas costeras de extensión reducida, carentes de vegetación sumergida y con valores bajos de salinidad, presentan una menor riqueza de especies que las lagunas con características opuestas. La variación en estas características se relaciona con el grado de influencia marina, circulación, ciclos biogeoquímicos y heterogeneidad de hábitat. Estos factores tienen implicación en la composición de especies que pueden pertenecer en menor o mayor grado al componente dulceacuícola, marino y estuarino (Caballero-Vázquez et al., 2005).
La mayoría de los peces que integraron la comunidad fueron juveniles, lo que sugiere el uso de estos hábitats como zonas de crianza (Rakocinski et al., 1992). Teolán fue el estuario con la mayor abundancia, debido al gran número de individuos juveniles de A. monticola y M. curema capturados durante el mes de septiembre. De esta manera, septiembre fue el mes con mayor abundancia, un resultado que puede relacionarse con las características fisicoquímicas y biológicas que ocurren durante este mes y que se relacionan con el aumento de la precipitación. Mugil curema se alimenta de detritus y restos vegetales (Franco y Bashirullah, 1992), mientras que A. monticola se alimenta en mayor medida de insectos y algas (Torres-Navarro y Lyons, 1999). Ambas especies aprovechan las condiciones de los estuarios durante la época de lluvias, donde el aumento del material orgánico arrastrado por los ríos y el aumento de la producción primaria, facilita el desarrollo de los juveniles (Kostecki et al., 2012). La presencia de un gran número de individuos de tallas pequeñas sugiere una estrategia del tipo catádromo para A. monticola y anádromo para M. curema, comportamiento que ya ha sido documentado (Castro-Aguirre et al., 1999; Eslava-Eljaiek y Díaz-Vesga, 2011).
Barra de Pichi mostró el mayor valor de biomasa, representada principalmente por D. latifrons y G. maculatus, con tallas mayores a las del resto de las especies. Estas especies son consideradas habitantes permanentes de los ecosistemas estuarinos, donde transcurre todo su ciclo de vida (Castro-Aguirre et al., 1999); presentan, además, una alta tolerancia a la variación ambiental y a la capacidad para sobrevivir en ambientes deficientes de oxígeno y con altas variaciones en salinidad y temperatura (Ancieta y Landa, 1977).
Barra de Nexpa registró la mayor riqueza y el mayor porcentaje de especies raras. Además, fue el único estuario que el análisis de rarefacción determinó como significativamente diferente, en relación con la riqueza. El número de especies dominantes fue mayor y con una composición diferente a la del resto de los estuarios. La riqueza y composición de especies raras puede relacionarse a características del estuario, como el grado de conexión con el océano y el volumen del cuerpo de agua (Mendoza et al., 2009). Se apreció que este estuario cuenta con una extensión mayor en comparación con los otros, además de que se observaron características particulares que no se encontraron en los otros cuerpos de agua. La presencia de vegetación sumergida y una mayor complejidad en la morfología del estuario reflejada en ramificaciones en su afluente, pequeños meandros, barras y zonas de inundación aisladas, son factores que pueden influir en la riqueza y composición de especies (Raz-Guzmán y Huidobro, 2002; Pérez-Ruzafa et al., 2007).
Barra de Pichi fue el ecosistema con la menor riqueza, lo que puede ser resultado de las condiciones de deterioro que ha sufrido este cuerpo de agua. Sin embargo, en este estuario se registró un porcentaje de especies únicas del 9.7%, como Anchoa lucida, Pliosteostoma lutipinnis y Sphoeroides annulatus. Cabe señalar, que estas especies se registraron en septiembre y febrero, donde las condiciones de oxígeno disuelto y transparencia aumentaron con respecto a junio. Lo que indica que estos organismos evitarían las condiciones de perturbación.
La variación temporal de la estructura de la comunidad mostró un patrón influenciado por la estacionalidad de la precipitación. La riqueza fue mayor en junio, al inicio de la temporada de lluvias y menor en septiembre, durante el máximo de la estación lluviosa. En febrero se registró un mayor número de especies dominantes, lo que indica una mayor estabilidad del número de individuos por especie. La variabilidad en la composición de las especies dominantes puede ser consecuencia de diferencias en la salinidad, así como condiciones ambientales desfavorables para los movimientos migratorios de especies que no son componentes habituales del estuario y que sólo ingresan en temporadas específicas (Laroche et al., 1997).
El Decorana mostró ser una herramienta útil para determinar la relación entre las variables ambientales con respecto a la frecuencia y abundancia de especies. Los conjuntos de especies indican que la composición está determinada por condiciones ambientales que varían espacial y temporalmente. Existen especies con preferencias de hábitat específicas relacionadas con la biología de la especie, la tolerancia a la salinidad, los hábitos alimenticios y el ciclo de vida. El Grupo I, definido por el Decorana, estuvo asociado a condiciones donde la profundidad fue mayor; particularmente Ctenogobius sagittula, Dactyloscopus amnis junto con Microdesmus dorsipunctatus y Pseudophallus starksii se colectaron solamente en un muestreo, lo que sugiere la preferencia de estas especies por características fisicoquímicas específicas. En especial, la riqueza y densidad de algunas especies estuarinas se asocian positivamente a la profundidad (Thomas y Connolly, 2001). Como se ha observado en otros trabajos, estos estuarios son utilizados en gran medida por habitantes temporales que buscan zonas para alimentación o crianza (Selleslagh y Amara, 2008). El recambio en la composición es una característica común en estuarios tropicales y templados. Este comportamiento puede estar relacionado con los periodos de reproducción y reclutamiento, así como la variación estacional en las densidades de zooplancton (Kimmerer, 2002; González-Acosta et al., 2005). El Grupo II se asoció con valores intermedios de todas las variables ambientales; a excepción de Poecilia butleri, el resto de las especies de este grupo pertenecen al componente marino, por lo que se consideran visitantes ocasionales de los ecosistemas estuarinos que ingresan al ecosistema cuando se presenta una conexión con el océano (Torres-Orozco, 1994; Yáñez-Arancibia et al., 1994). Las especies de este grupo presentaron preferencia por condiciones ambientales con poca variación, por lo que podría considerárseles como estenoicas. El Grupo III estuvo definido por valores bajos de salinidad, temperatura y transparencia y valores intermedios de oxígeno; estas condiciones se asocian con un aumento en la precipitación, ya que el agua dulce aportada por los ríos disminuyó la salinidad y la temperatura, mientras que la disminución de la transparencia es resultado del aumento del material orgánico. En este grupo se conjuntaron especies de consumidores primarios como L. nigrofasciata (filtrador), A. monticola (fitófago y detritívoro) y M. curema (detritívoro) (Franco y Bashirullah, 1992; Torres-Navarro y Lyons, 1999). El resto de las especies de este grupo pertenecen al medio marino sin que exista un patrón claro que defina las características de su agrupamiento. El Grupo IV estuvo definido por condiciones de alta transparencia, elevada temperatura y poca profundidad. En este grupo se conjuntaron especies que pertenecen en su mayoría al componente estuarino. Awaous transandeanus, D. latifrons, Eleotris picta, Gobionellus microdon y G. maculatus son consideradas como especies que habitan permanentemente los ecosistemas estuarinos (Castro-Aguirre et al., 1999). En general, la comunidad de peces estuvo compuesta por especies marinas y dulceacuícolas que utilizan los estuarios como zonas de crianza, especies marinas que ingresan a los estuarios de manera ocasional y especies estuarinas que permanecen en estos ecosistemas durante la mayor parte de su ciclo de vida.
La información obtenida en este estudio ofrece un panorama general sobre la riqueza y composición de la comunidad de peces estuarinos de Michoacán. La abundancia de peces juveniles encontrada, implica que estos estuarios ofrecen recursos para el abastecimiento de las pesquerías locales. Estos resultados pueden ser útiles para la investigación en Michoacán en materia de conservación de peces y manejo de recursos pesqueros. Los registros de especies no reportadas para México, como en el caso Atherinella panamensis (Sandoval-Huerta et al., 2012a), contribuyen al conocimiento de la biodiversidad del país. Por otra parte, se propone desarrollar un plan de manejo integral para el estuario de barra de Pichi, con el objetivo de establecer medidas de conservación y restauración. El mejoramiento de la calidad ambiental del cuerpo de agua afectará positivamente la población que explota sus recursos pesqueros y la biodiversidad que se encuentra en el estuario y en los ecosistemas costeros vecinos. En el caso de la especie exótica del género Oreochromis, se asume que su presencia es resultado de su liberación accidental o deliberada por parte de las granjas de tilapia que se ubican río arriba. Se recomienda la implementación de medidas para su control que eviten su dispersión en otros cuerpos de agua del estado, como ha ocurrido con los loricáridos sudamericanos del género Pterygoplichthys en el estuario del río Coahuayana (Sandoval-Huerta et al., 2012b). Las especies invasoras representan uno de los problemas más severos y menos controlados en México (Contreras-MacBeath et al., 2014), por lo que su erradicación representa un escenario complicado, debido a que requiere de la integración de proyectos por parte de las instituciones correspondientes, quienes lejos de establecer medidas de control, fomentan prácticas que favorecen la introducción de especies exóticas.
Agradecimientos
Los autores agradecen a la Conabio HJ021 y a la Coordinación de la Investigación Científica de la Universidad Michoacana, por el financiamiento para este estudio. Al Dr. Adrián Felipe González-Acosta por su ayuda en la identificación de los organismos y al M. C. Rubén Hernández-Morales por su apoyo en los análisis de datos. Así como a todas las personas que colaboraron en las jornadas de campo en el proyecto Ictiofauna de la Costa de Michoacán.
Literatura citada
Aceves-Medina, G., R. Saldierna-Martínez, A. Hinojosa-Medina, S. P. A. Jiménez-Rosenberg, M. E. Hernández-Rivas y R. Morales-Ávila. 2008. Vertical structure of larval fish assemblages during diel cycles in summer and winter in the southern part of Bahía de La Paz, Mexico. Estuarine, Coastal and Shelf Science 76:889-901. [ Links ]
Alcalá, G., G. Marín, A. Toledo, J. A. Cano, S. Farreras y J. Viqueira. 2003. Espacios y actividades costeras en Michoacán. Aproximaciones varias. El Colegio de México/Centro de Investigación Científica y de Educación Superior de Ensenada/El Colegio de Michoacán, Zamora. 210 p. [ Links ]
Álvarez-Arellano, A. D. y J. Gaitán-Morán. 1994. Lagunas costeras y el litoral mexicano: Geología. In Lagunas costeras y el litoral mexicano, E. G. De la Lanza-Espino y C. Cáceres-Martínez (eds.). Universidad Autónoma de Baja California Sur, La Paz. p. 13-74. [ Links ]
Ancieta, D. F. y A. Landa. 1977. Reseña taxonómica y biológica de los peces cultivados en el área andina incluyendo la costa del Perú. La acuicultura en América Latina. FAO, Informes de Pesca 2:106-113. [ Links ]
Antaramián, H. E. y G. Correa. 2003. Fisiografía. In Geografía física. Atlas Geográfico del Estado de Michoacán, H. E. Antaramián (coord.). SEEM / UMSNH/EDISSA, S. A. de C. V. Morelia. p. 42-46. [ Links ]
Ayala-Pérez, L. A., J. Ramos y D. Flores. 2003. La comunidad de peces de la Laguna de Términos: estructura actual comparada. Revista de Biología Tropical 51:783-794. [ Links ]
Borges, P. A., S. Train, J. D. Dias y C. C. Bonecker. 2010. Effects of fish farming on plankton structure in a brazilian tropical reservoir. Hydrobiologia 649:279-291. [ Links ]
Caballero-Vázquez, J. A., H. C. Gamboa-Pérez y J. J. Schmitter-Soto. 2005. Composition and spatio-temporal variation of the fish community in the Chacmochuch lagoon system, Quintana Roo, Mexico. Hidrobiológica 15:215-225. [ Links ]
Castillo-Rivera, M., R. Zárate y S. Ortiz. 2005. Variación nictímera y estacional de la abundancia, riqueza y especies dominantes de peces, en un ambiente salobre de sustrato blando. Hidrobiológica 15:227-238. [ Links ]
Castro-Aguirre, J. L., H. S. Espinosa y J. J. Schmitter-Soto. 1999. Ictiofauna estuarino-lagunar y vicaria de México. Colección de Textos Politécnicos. Instituto Politécnico Nacional y Limusa Noriega Editores, México D. F. 711 p. [ Links ]
Colwell, R. K., C. X. Mao y J. Chang. 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 85:2717-2727. [ Links ]
Contreras-MacBeath, T., M. T. Gaspar-Dillanes, L. Huidobro-Campos y H. Mejía-Mojica. 2014. Peces invasores en el centro de México. In Especies acuáticas invasoras en México, R. Mendoza y P. Koleff (coords.). Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, D. F. p. 413-424. [ Links ]
Correa, G. P. y M. Ayala. 2003. Hidrografía e hidrología. In Geografía física. Atlas Geográfico del Estado de Michoacán, H. E. Antaramián (coord.). SEEM/UMSNH/EDISSA, S. A. de C. V. Morelia. p. 47-50. [ Links ]
Correa, G. P. y M. Gómez R. 2003. Litoral. In Geografía física. Atlas Geográfico del Estado de Michoacán, H. E. Antaramián (coord.). SEEM/UMSNH/EDISSA, S. A. de C. V. Morelia. p. 42-46. [ Links ]
Day, J. W. y A. Yáñez-Arancibia. 1985. Coastal lagoons and estuaries as an environment for nekton. In Fish community ecology in estuaries and coastal lagoons: towards an ecosystem integration, A. Yáñez-Arancibia (ed.). Universidad Nacional Autónoma de México. México D. F. p. 17-34. [ Links ]
De la Lanza-Espino, G. 1994. Química de las lagunas costeras y el litoral mexicano. In Lagunas costeras y el litoral mexicano, G. De la Lanza-Espino y C. Cáceres-Martínez (eds.). Universidad Autónoma de Baja California Sur, La Paz. p. 127-198. [ Links ]
De la Lanza-Espino, G. 1998. Aspectos fisicoquímicos que determinan la calidad del agua. In Ecología de los sistemas acuícolas, C. L. R. Martínez (ed.). AGT, México, D. F. p. 1-26. [ Links ]
Díaz-Ruiz, S., A. Aguirre-León y E. Cano-Quiroga. 2006. Evaluación ecológica de las comunidades de peces en dos sistemas lagunares estuarinos del sur de Chiapas, México. Hidrobiológica 16:197-210. [ Links ]
Eby, L. A. y L. B. Crowder. 2004. Effects of hypoxic disturbances on an estuarine nekton assemblage across multiple scales. Estuaries 27:342-351. [ Links ]
Elliott, M. y A. K. Whitfield. 2011. Challenging paradigms in estuarine ecology and management. Estuarine, Coastal and Shelf Science 94:306-314. [ Links ]
Eslava-Eljaiek, P. y R. Díaz-Vesga. 2011. Reproducción de Joturus pichardi y Agonostomus montícola (Mugiliformes: Mugilidae) en ríos de la sierra Nevada de Santa Marta, Colombia. Revista de Biología Tropical 59:1717-1728. [ Links ]
Fischer, W., F. Krupp, W. Schneider, C. Sommer, K. E. Carpenter y V. H. Niem. 1995. Guía FAO para la identificación de especies para los fines de la pesca: Pacífico centro-oriental. Rome, FAO. Volúmenes II y III:647-1813. [ Links ]
Franco, L. y K. M. B. Bashirullah. 1992. Alimentación de la lisa (Mugil curema) del golfo de Cariaco-estado Suche, Venezuela. Zootecnia Tropical 10:219-238. [ Links ]
García, E. 1981. Modificaciones al sistema climático de Köppen (para adaptarlo a la República Mexicana). Universidad Nacional Autónoma de México. México, D. F. 493 p. [ Links ]
Garduño, M. V. H., C. P. Corona y A. I. Israde. 2003. Geología. In Geografía física. Atlas geográfico del estado de Michoacán, H. E. Antaramián (coord.). SEEM/UMSNH/EDISSA, S. A. de C. V. Morelia. p. 37-41. [ Links ]
González-Acosta, A. F., G. De la Cruz-Agüero, J. De la Cruz-Agüero y G. Ruiz-Campos. 2005. Seasonal pattern of the fish assemblage of El Conchalito mangrove swamp, La Paz Bay, Baja California Sur, Mexico. Hidrobiológica 15:205-214. [ Links ]
Gotelli, N. J. y A. M. Ellison. 2013. Null modeling software for ecologists. EcoSimR 1.00. Disponible en http://www.uvm.edu/~ngotelli/EcoSim/EcoSim.html; última consulta: 02.IX.2013. [ Links ]
Hill, M. O. 1979. Decorana. A fortran program for detrended correspondence analysis and reciprocal averaging. Cornell University, New. York. [ Links ]
Kimmerer, W. J. 2002. Effects of freshwater flow on abundance of estuarine organisms: physical effects or trophic linkages? Marine Ecology Progress Series 243:39-55. [ Links ]
Klein, G. de V. 1985. Sandstone depositional models for exploration for fossil fuels. International Human Resources Development Corporation, Boston. 114 p. [ Links ]
Kostecki, C., J. M. Roussel, N. Desroy, G. Roussel, J. Lanshere, H. Le Bris y O. Le Pape. 2012. Trophic ecology of juvenile flatfish in a coastal nursery ground: contributions of intertidal primary production and freshwater particulate organic matter. Marine Ecology Progress Series 449:221-232. [ Links ]
Köppen, W. 1948. Climatología. Fondo de Cultura Económica, México, D. F. 478 p. [ Links ]
Laroche, J., E. Baran y N. B. Rasoanandrasana. 1997. Temporal patterns in a fish assemblage of a semiarid mangrove zone in Madagascar. Journal of Fish Biology 51:3-20. [ Links ]
Llansó, R. J. 1992. Effects of hypoxia on estuarine benthos: the lower Rappahannock river (Chesapeake bay), a case study. Estuarine, Coastal and Shelf Science 35:491-515. [ Links ]
Loneragan, N. R. 1999. River flows and estuarine ecosystems: implications for coastal fisheries from a review and a case study of the Logan river, southeast Queensland. Australian Journal of Ecology 24:431-440. [ Links ]
Lui, H. K. y C. T. A. Chen. 2012. The nonlinear relationship between nutrient ratios and salinity in estuarine ecosystems: implications for management. Current Opinion in Environmental Sustainability 4:227-232. [ Links ]
Lyons, J. y D. W. Schneider. 1990. Factors influencing fish distribution and community structure in a small coastal river in south-western Costa Rica. Hydrobiologia 203:1-14. [ Links ]
Madrigal, G. X. 2006. Distribución espacial y temporal de la ictiofauna del estero se Santa Ana, Michoacán, México. Tesis maestría, Posgrado de Ciencias Biológicas, Universidad Nacional Autónoma de México. México D. F. 77 p. [ Links ]
McCune, B. y M. J. Mefford. 2005. PC-ORD, multivariate analysis of ecological data. Version 5 for Windows edition. MjM Software Design, Gleneden Beach, Oregon. [ Links ]
Mendoza, E., M. Castillo-Rivera, R. Zárate-Hernández y S. Ortiz-Burgos. 2009. Seasonal variations in the diversity, abundance, and composition of species in an estuarine fish community in the Tropical Eastern Pacific, Mexico. Ichthyological research 56:330-339. [ Links ]
Miller, R. R., W. L. Minckley y S. M. Norris. 2005. Freshwater fishes of Mexico. The University of Chicago Press, Chicago. 490 p. [ Links ]
Morales, J. y J. L. Pérez. 2006. Crecimiento poblacional e instrumentos para la regulación ambiental de los asentamientos humanos en los municipios costeros. Gaceta Ecológica 79:53-77. [ Links ]
Pastor, L., B. Deflandre, E. Viollier, C. Cathalot, E. Metzger, C. Rabouille y A. Grémare. 2011. Influence of the organic matter composition on benthic oxygen demand in the Rhône river prodelta (NW Mediterranean sea). Continental Shelf Research 31:1008-1019. [ Links ]
Pérez-Ruzafa, A., M. C. Mompean y C. Marcos. 2007. Hydrographic, geomorphologic and fish assemblage relationships in coastal lagoons. Hydrobiologia 577:107-125. [ Links ]
Pritchard, D. W. 1967. What is an estuary: physical view point. Estuaries 83:3-5. [ Links ]
Rakocinski, C. F., D. M. Baltz y J. W. Fleeger. 1992. Correspondence between environmental gradients and the community structure of marsh-edge fishes in a Louisiana estuary. Marine Ecology Progress Series 80:135-148. [ Links ]
Ray, G. C. 2005. Connectivities of estuarine fishes to the coastal realm. Estuarine, Coastal and Shelf Science 64:18-32 [ Links ]
Raz-Guzmán, A. y L. Huidobro. 2002. Fish communities in two environmentally different estuarine systems of Mexico. Journal of Fish Biology 61:182-195 [ Links ]
Sandoval-Huerta, E. R., X. Madrigal-Guridi, A. García-Meraz, N. I. Dimas-Mora, O. Domínguez-Domínguez y A. F. González-Acosta. 2012a. First records of Atherinella panamensis in the central mexican Pacific coasts. Cybium, International Journal of Ichthyology 36:587-588. [ Links ]
Sandoval-Huerta, E. R., X. Madrigal-Guridi, A. García-Meraz, N. I. Dimas-Mora y O. Domínguez-Domínguez. 2012b. Nuevo registro de Pterygoplichthys disjunctivus (Actinopterygii: Loricariidae) en la desembocadura del río Coahuayana, Coahuayana, Michoacán. Revista Mexicana de Biodiversidad 83:294-297. [ Links ]
Selleslagh, J. y R. Amara. 2008. Environmental factors structuring fish composition and assemblages in a small macrotidal estuary (Eastern English Channel). Estuarine, Coastal and Shelf Science 79:507-517. [ Links ]
Sokal, R. R. y F. J. Rohlf. 1985. Biometría. Glume. Barcelona. 587 p. [ Links ]
Thomas, B. E. y R. M. Connolly. 2001. Fish use of subtropical saltmarshes in Queensland, Australia: relationships with vegetation, water depth and distance onto the marsh. Marine Ecology Progress Series 209:275-288. [ Links ]
Toledo, A. y L. Bozada. 2002. El delta del río Balsas. Medio ambiente, pesquerías y sociedad. Instituto Nacional de Ecología, El Colegio de Michoacán, A. C., México. D. F. 294 p. [ Links ]
Torres-Alvarado, M. del R., F. Fernández P., I. de los A. Barriga-Sosa y F. Ramírez-Vives. 2006. Dinámica de las bacterias anaeróbicas en las fases terminales de la mineralización de la materia orgánica en el sedimento de los ecosistemas Carretas-Pereyra y Chantuto-Panzacola. Hidrobiológica 16:183-196. [ Links ]
Torres-Navarro, C. I. y J. Lyons. 1999. Diet of Agonostomus monticola (Pisces: Mugilidae) in the Río Ayuquila, Sierra de Manantlán Biosphere Reserve, México. Revista de Biología Tropical 47:1087-1092. [ Links ]
Torres-Orozco, R. 1994. Los peces. In Lagunas costeras y el litoral mexicano, G. De la Lanza-Espino y C. Cáceres-Martínez (eds.). Universidad Autónoma de Baja California Sur, La Paz. p. 269-304. [ Links ]
Valiela, I., L. Camilli, T. Stone, A. Giblin, J. Crusius, S. Fox y C. Harris. 2012. Increased rainfall remarkably freshens estuarine and coastal waters on the Pacific coast of Panama: magnitude and likely effects on upwelling and nutrient supply. Global and Planetary Change 92:130-137. [ Links ]
Van Rijn, L. C. 1993. Principles of sediment transport in rivers, estuaries, and coastal seas. Aqua Publications, Amsterdam. 614 p. [ Links ]
Whitfield, A. K. 1999. Ichthyofaunal assemblages in estuaries: a South African case study. Reviews in Fish Biology and Fisheries 9:151-186. [ Links ]
Woodland, R. J., D. H. Secor, M. C. Fabrizio y M. J. Wilberg. 2012. Comparing the nursery role of inner continental shelf and estuarine habitats for temperate marine fishes. Estuarine, Coastal, and Shelf Science 99:61-73. [ Links ]
Wu, R. S. 2002. Hypoxia: from molecular responses to ecosystem responses. Marine Pollution Bulletin 45:35-45. [ Links ]
Yáñez-Arancibia, A., A. L. Lara-Domínguez y D. Pauly. 1994. Coastal lagoons as fish habitat. In Coastal lagoon processes, K. Kjerfve (ed.). Elsevier, Amsterdam. p. 363-376. [ Links ]

