










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de biodiversidad
On-line version ISSN 2007-8706Print version ISSN 1870-3453
Rev. Mex. Biodiv. vol.85 n.3 México Sep. 2014
https://doi.org/10.7550/rmb.35806
Ecología
Composición y estructura de la ictiofauna del río Hondo, México-Belice, con base en el uso del arpón
Composition and structure of the ichthyofauna in the Hondo River, Mexico-Belize, using a harpoon
Jesús M. López-Vila1*, Martha E. Valdéz-Moreno1, Juan J. Schmitter-Soto1, Manuel Mendoza-Carranza2 y Roberto L. Herrera-Pavón1
1 El Colegio de la Frontera Sur. Av. Centenario km 5.5, 77900 Chetumal, Quintana Roo, México. * jlopezv@ecosur.edu.mx
2 El Colegio de la Frontera Sur. Carretera a Reforma km 15.5, Ranchería Guineo, segunda sección, 86280 Villahermosa, Tabasco, México.
Recibido: 13 febrero 2013
Aceptado: 10 marzo 2014
Resumen
Se determinó la composición y estructura de la ictiofauna del río Hondo y la variación espacial de éstas en función de los parámetros ambientales. Se realizaron 3 muestreos durante marzo, abril y mayo de 2012, en 6 sitios a lo largo del río. Los datos usados para determinar la estructura se obtuvieron con ayuda de un arpón de 58.5 cm, utilizado durante 2 h de buceo libre en cada punto de muestreo. Para complementar la lista de especies se utilizaron redes agalleras, palangres, nasas y anzuelos. Además, se realizó una revisión bibliográfica y se consultó material de la colección de peces de ECOSUR en Chetumal. La lista sistemática se conformó por 40 especies en 33 géneros, 18 familias y 11 órdenes. La familia mejor representada fue Cichlidae, incluida la especie exótica Oreochromis niloticus. Los parámetros ambientales que mejor describieron la variación de la distribución y abundancia en los peces en el río Hondo fueron: la anchura del río, distancia a la boca, transparencia, profundidad, concentración de oxígeno disuelto y conductividad. La relación entre las especies y las variables ambientales fue alta en ambos ejes (0.89 y 0.79). Ambientalmente, el río se puede zonificar en 3 partes, aunque su ictiofauna puede dividirse en 2 conjuntos principales.
Palabras clave: diversidad, distribución, ecología de ríos, peces, variables ambientales, Quintana Roo.
Abstract
Composition and structure of the Hondo River ichthyofauna and its spatial variation were determined as a function of environmental parameters. Six sites along the river were sampled in March, April and May 2012. Data for estimating structure of fish assemblages was obtained by means of a 58.5 cm harpoon during 2 hours of free diving at each sampling site. The species list for the study area was completed with the aid of gillnets, longlines, double-cone traps, and hook-and-line. In addition, a bibliographical research performed as well as the ichthyological collection of ECOSUR at Chetumal was checked. The systematic list includes 40 species in 33 genera, 18 families and 11 orders. The richest family was Cichlidae, including the exotic Oreochromis niloticus. The environmental parameters that best described distribution and abundance of fish in the Hondo River were river width, distance to mouth, transparency, depth, dissolved oxygen concentration, and conductivity. Correlation between species and environmental parameters was high in both axis (0.89 and 0.79). The river was classified into 3 environmental zones, although its fish fauna could be divided in 2 main assemblages.
Key words: diversity, distribution, river ecology, fishes, environmental parameters, Quintana Roo.
Introducción
El río Hondo es el único río epicontinental en el estado de Quintana Roo y se ubica dentro de la provincia ictiológica del Usumacinta, la cual se caracteriza por poseer un gran número de especies endémicas de las familias Poeciliidae y Cichlidae, así como muchas especies marinas que viven o penetran en agua dulce (Miller, 1982). Debido a la alta diversidad biológica que este río alberga, la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio) lo declaró, en 1998, como Región Hidrológica Prioritaria Núm. 110 (Arriaga-Cabrera et al., 1998).
A pesar de lo anterior, el conocimiento de la diversidad íctica del cauce principal del río Hondo, se ha limitado en gran medida a listas faunísticas (Gamboa-Pérez, 1991, 1994; Espinosa-Pérez et al., 1993; Schmitter-Soto y Caro, 1997; Arriaga-Cabrera et al., 1998; Schmitter-Soto, 1998; Scholz y Vargas-Vázquez, 1998; Esselman, 2009; Schmitter-Soto et al., 2009; Ruiz-Cauich, 2011) que en su conjunto han aportado 38 especies hasta el momento, pero sin entrar en detalles sobre cómo está conformada la estructura de la comunidad de peces. La mayoría de estos trabajos se enfocaron en muestrear en partes someras del río, lo cual permitió obtener registros, principalmente, de tallas y especies pequeñas, no así de las tallas y especies grandes, las cuales son especies de importancia pesquera para esta zona.
Por otro lado, a pesar de que se han realizado estudios que documentan algunos datos sobre los parámetros ambientales del río Hondo (Schmitter-Soto y Gamboa-Pérez, 1996; De Jesús-Navarrete y Góngora-Pech, 2009; Pacheco-Díaz, 2011; Schmitter-Soto et al., 2011) ninguno de ellos ha analizado la relación entre estas variables ambientales y la composición y abundancia de los peces. Por tal motivo, el objetivo de este trabajo fue el de evaluar la estructura de la ictiofauna del cauce principal del río Hondo en términos de su riqueza, distribución, abundancia y diversidad específica; utilizando un arpón para poder capturar tallas y especies grandes; la composición se determinó de manera bibliográfica, revisando la colección ictiológica de ECOSUR y con registros obtenidos en este trabajo. Así también se evaluaron los factores ambientales que determinan dicha estructura.
Materiales y métodos
El río Hondo se localiza en el estado de Quintana Roo, entre las coordenadas 18°45' y 17°43' de latitud norte y 88°35' y 89º15' de longitud oeste (Magnon-Basnier, 1996). Nace en Guatemala, donde se le conoce con el nombre de río Azul (Gamboa-Pérez 1991; Herrera-Sansores y Heredia-Escobedo, 2011) y desemboca en la bahía de Chetumal. Tiene una longitud de 145 km, 50 m de ancho en promedio, una profundidad media de 10 m y su cuenca tributaria tiene un área de 13 465 km2 (Herrera-Sansores y Heredia-Escobedo, 2011). Este río se alberga en una falla geológica (Álvarez-Legorreta, 2011) y su cauce marca una frontera natural entre México y Belice. El clima de la región es cálido subhúmedo con lluvias en verano, la temperatura media es de 26° C y la precipitación media de 1 350 mm. La vegetación ribereña es bastante diversa: selvas mediana subcaducifolia, baja perennifolia, baja inundable, sabana, cañaveral, pastizal cultivado y manglar (Arriaga-Cabrera et al., 1998).
Se realizaron 3 muestreos mensuales en la época de secas e inicio de lluvias; marzo, abril y mayo de 2012; en 6 sitios del río distribuidos desde la boca hasta la parte alta de éste (Fig. 1). El principal arte de pesca, y en el que se basó la parte cuantitativa del estudio, debido a la ineficacia de las artes de captura pasiva (redes agalleras), fue un arpón de una liga con una varilla de acero de 58.5 cm. El esfuerzo de muestreo se estandarizó con un tiempo de pesca de 2 h por sitio, tiempo en el cual el pescador nadó paralelo a la orilla del río pescando a una profundidad de entre 1 y 6 m entre las raíces de la vegetación ribereña; la actividad se realizó durante el día. Para capturar especies asociadas al fondo, se colocaron 3 trampas tipo nasa; 90 cm de largo, 45 cm de diámetro y apertura de 13 cm; anzuelos de diferentes tamaños y un palangre con 10 anzuelos número 11. Los organismos capturados fueron contados, pesados y medidos en campo; la mayoría de éstos fueron identificados in situ, aquellos que no pudieron serlo se transportaron en una hielera al laboratorio para su determinación taxonómica. Para completar la lista sistemática de las especies se revisó bibliografía especializada y la colección ictiológica de ECOSUR-Chetumal (ECOCH). Los peces se identificaron con las claves taxonómicas de Greenfield y Thomerson (1997), Schmitter-Soto (1998) y Miller et al. (2009) y la revisión sistemática reciente de McMahan et al. (2011). El arreglo sistemático a nivel de familia siguió a Nelson (2006) y la actualización de nomenclatura a Eschmeyer y Fricke (2012).

En cada sitio de muestreo, con ayuda de una sonda multiparamétrica marca Horiba modelo U-10, se midieron las variables físico-químicas del agua: temperatura (°C), salinidad (ups), conductividad (mS/cm), oxígeno disuelto (mg/l) y pH, tanto en el fondo como en la superficie del río. También se registraron las siguientes características del río en cada sitio de muestreo: anchura del río —distancia perpendicular entre ambas riberas del río— con un GPS marca Garmin y el programa Google Earth; distancia a la boca del río de cada lugar de muestreo; transparencia del agua con un disco Secchi y profundidad máxima, la cual se midió en la parte medial del canal utilizando una plomada. Además, se registró de manera cualitativa el tipo de vegetación.
Para analizar la estructura de la comunidad íctica del río Hondo, capturada con el arpón, se tomó en cuenta la composición y riqueza de las especies; la dominancia se calculó con base en el índice de valor de importancia relativa (IVIr), el cual toma en cuenta la biomasa, abundancia y frecuencia relativa de las especies; la diversidad, con base en el índice de Shannon (H'n) y la equidad, con el índice de Pielou (J'n) (Magurran, 1998). La distribución espacial de las especies se analizó con un cuadro de presencia-ausencia, con el cual se obtuvo un dendrograma con base en el índice de similitud de Bray-Curtis y la estrategia de agrupamiento UPGMA, para así conocer el grado de asociación entre sitios.
Con el fin de conocer las variables que mejor reflejaron la dinámica del río, se realizó un análisis de componentes principales (ACP) con el paquete estadístico Permanova 6 + Primer, versión 1.0.1 (Clarke y Warwick, 2006). La relación entre las variables ambientales y los peces se evaluó mediante un análisis de correspondencia canónica (ACC) con la ayuda del programa Canoco, versión 4.5 (Ter-Braak y Smilauer, 2002); no se tomaron en cuenta las especies representadas por 1 o 2 individuos. Los datos del ACP se normalizaron y, posteriormente, se transformaron logarítmicamente; los datos del ACC sólo se transformaron logarítmicamente. En el caso de las variables que presentaron colinealidad, se optó por una de cada par y aquellas variables con baja capacidad de explicación tampoco se tomaron en cuenta. De este modo, para el ACP y ACC se utilizaron las siguientes variables: anchura del río, transparencia, profundidad, distancia a la boca, conductividad superficial y oxígeno disuelto en la superficie.
Resultados
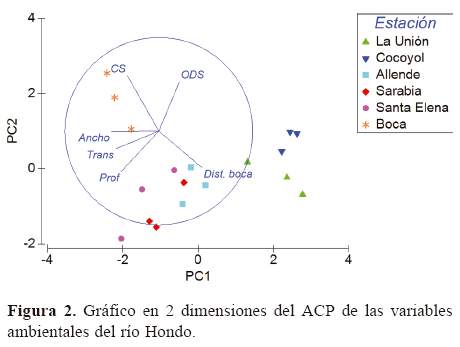
Parámetros ambientales y de estructura de hábitat que dominan en el río Hondo. De acuerdo con el ACP, el 83% de la variación fue explicada por los 2 primeros componentes. El eje 1 expresó un 55.8% y fue dominado por las siguientes variables: transparencia, profundidad, anchura del río y distancia a la boca. Por otro lado, el eje 2 aportó un 22.7% de la variación, y las variables más importantes tienen que ver con conductividad y oxígeno disuelto, así como otro parámetro de estructura del río: la profundidad. El gráfico en 2 dimensiones (Fig. 2) agrupa los sitios de muestro en 3 conjuntos que dividen al río en 3 secciones: alta (Cocoyol y La Unión), media-baja (Allende, Santa Elena y Sarabia) y desembocadura (boca). Además, muestra un gradiente marcado por la distancia a la boca y la conductividad superficial, desde La Unión hasta la desembocadura. La conductividad permitió diferenciar la región estuarina de la dulceacuícola, debido a que los valores más altos de esta variable se midieron en Santa Elena y boca, especialmente en el fondo, lo que hizo evidente la presencia de una cuña salina que llega hasta Santa Elena en la época de sequía.
Composición y estructura de la ictiofauna. Se capturó un total de 353 peces agrupados en 6 órdenes, 11 familias, 17 géneros y 18 especies (Cuadro 1), entre éstas una exótica: Oreochromis niloticus. Los organismos capturados acumularon una biomasa de 111 312 g. A estas especies se le suman 22 más como resultado de la revisión bibliográfica y de la colección ictiológica de ECOSUR-Chetumal, de modo que la ictiofauna del río Hondo está conformada por 11 órdenes, 18 familias, 33 géneros y 40 especies (Cuadro 1).
De las 19 especies registradas en el muestreo, sólo 3 no fueron capturadas con arpón: Astyanax cf. aeneus, Rhamdia laticauda y Megalops atlanticus, esta última se encontró flotando muerta en la orilla del río; las 16 restantes fueron incluidas en los análisis cuantitativos de la estructura de la ictiofauna del río Hondo. Con base en el material recolectado en campo la familia mejor representada fue Cichlidae, con 9 especies, mientras que las familias Ariidae, Belonidae, Characidae, Gerreidae, Lutjanidae, Poeciliidae y Tetraodontidae estuvieron representadas por una especie cada una. La especie más importante por su alta abundancia a lo largo de todo el río fue Paraneetroplus melanurus —antes conocida como Cichlasoma synspilum—, ya que los 186 organismos de esta especie contribuyeron con el 55.69% de la abundancia total, así también lo fue en biomasa con 40 245 g, lo que representó un 51.76% de la biomasa total. De las especies capturadas destacan las siguientes por ser consumidas por los habitantes de la ribera del río: Cichlasoma urophthalmus, Eugerres plumieri, Ictalurus meridionalis, Lutjanus griseus, O. niloticus, P. melanurus y Petenia splendida.
El sitio con la mayor abundancia de peces, promedio de las 3 expediciones, fue boca (24.33±3.21 DE), mientras que el sitio Cocoyol presentó el valor promedio más bajo (4.33±2.41). Por otro lado, el valor promedio más alto de biomasa se registró en Santa Elena (7 878.33 g±2 162.17) y el más bajo en Cocoyol (869.33 g±364.77). La mayor riqueza promedio de especies se encontró en La Unión (6.66±1.15) y la menor en Cocoyol (3±1) y Santa Elena (3±0). Los sitios con mayor diversidad (H'n) fue La Unión (1.54±0.31), mientras que Santa Elena obtuvo el valor promedio más bajo de diversidad (0.62±0.19). Por su parte, la equidad (J'n) tuvo su valor promedio más alto en Cocoyol (0.908±0.15) y el menor en boca (0.52±0.21). Las especies dominantes durante la época de secas en el río Hondo, de acuerdo con el IVIr, fueron P. melanurus (41.09%), O. niloticus (14.16%). E. plumieri (13.24%) y C. urophthalmus (9.04%). En conjunto estas especies contribuyeron con el 77.5% del total del IVIr. El resto de las especies estuvieron por debajo del 7% de este índice.
Distribución. Las especies ampliamente distribuidas a lo largo de todo el río fueron C. urophthalmus y P. melanurus, seguidas de O. niloticus y P. splendida, las últimas 2 se encontraron en 5 de 6 sitios del río. Las especies que se restringieron a la región estuarina del río y a la zona de influencia de ésta fueron E. plumieri, M. atlanticus, I. meridionalis y Sphoeroides testudineus. El ordenamiento espacial de las especies (Cuadro 2) muestra una transición gradual desde La Unión hasta la desembocadura del río. Con base en el análisis de agrupamiento de las especies encontradas en cada sitio, el río se zonificó en 2 regiones (Fig. 4). Una sección corresponde a la parte baja del río y la otra a la parte alta.


Relación entre las especies y los parámetros ambientales y de estructura de hábitat. El ACC del río Hondo evidenció un gradiente asociado a las variables ambientales y a las relacionadas con la forma del río (Fig. 3). El gradiente expresado sobre el eje 1 muestra claramente el aumento de profundidad y de anchura del río desde la parte alta de éste hasta su desembocadura; la conductividad también presentó este aumento. El oxígeno mostró un patrón contrario a la conductividad, ya que los sitios con mayor cantidad de oxígeno se presentaron en la parte alta del río. Esta variable también demostró un gradiente relacionado con la profundidad, debido a que conforme la profundidad aumentaba el oxígeno disminuía.
A lo largo del gradiente mencionado sobre el eje 1, las especies que se relacionaron con sitios someros y con elevados valores de oxígeno y poca conductividad, ubicados en la parte alta y estrecha del río, fueron Amphilophus robertsoni, Cichlasoma salvini, Cryptoheros chetumalensis y Thorichthys meeki. Las especies C. urophthalmus y P. splendida tuvieron relación con sitios ricos en oxígeno, aguas poco profundas y con cierto grado de conductividad. Contrario a éstas estuvieron E. plumieri y Sciades assimilis, las cuales tuvieron preferencia por sitios poco profundos y con valores bajos de oxígeno disuelto. Al centro del gráfico se observó a P. melanurus y O. niloticus, las cuales se distribuyeron ampliamente a lo largo del río, lo que refleja la plasticidad de estas especies ante la variación de las variables tomadas en cuenta para este análisis. La correlación entre las especies y las variables ambientales fue alta en ambos ejes (0.89 para el eje 1 y 0.79 para el eje 2).
Discusión
El conjunto de variables abióticas que le confieren identidad al río Hondo durante la época de secas: son ancho del río, profundidad, transparencia, conductividad, oxígeno disuelto y distancia a la boca, esta última variable, si bien, no influye en las características morfológicas del río, sí lo hace en el comportamiento de las variables físico-químicas. Estas variables zonificaron al río en 3 regiones: 1) alta-media (La Unión y Cocoyol); 2)baja (Allende, Sarabia y Santa Elena) y 3) la región de la desembocadura (boca). De las variables anteriores, se ha señalado a la distancia al océano como mucho más importante que el tamaño o diversidad de hábitats al momento de explicar la distribución y estructura de la ictiofauna en un río (Lyons y Schneider, 1990).
En cuanto a la composición, las 40 especies que se dan a conocer en la lista sistemática de especies del río Hondo representan un 32% de la ictiofauna registrada en las aguas continentales de Quintana Roo, la cual se compone de 125 especies (Schmitter-Soto, 1998). Además, 19 de las 40 especies también se han registrado como componentes de la ictiofauna de la bahía de Chetumal (Pimentel-Cadena, 2001). Si se toman en cuenta las dimensiones del río Hondo; longitud, anchura y profundidad del río; se esperaría que la ictiofauna de éste fuera mayor a la encontrada, ya que otros ríos de la misma provincia ictiográfica con longitudes menores a la del río Hondo alberguen un mayor número de especies. Tal es el caso del río Champotón (Campeche) que con 48 km de longitud aloja 53 especies (López-López et al., 2009) y el del río Lacanjá (Chiapas) que ostenta 44 especies en 120 km de río (Rodiles-Hernández et al., 1999).
Este menor número de especies puede deberse a que las aguas del río Hondo son poco productivas (Camarena-Luhrs y Cobá-Cetina, 1991; Gamboa-Pérez, 1994), susceptibles a perturbaciones (Ruiz-Cauich, 2011) y reciben todos los residuos contaminantes derivados del cultivo de caña en la ribera del río. Por otro lado, la juventud geológica de la península, así como el drenaje superficial poco desarrollado en esta región, han generado una ictiofauna
relativamente pobre (Hubbs, 1936; Miller, 1991).
Con base en la lista de especies, las familias mejor representadas fueron Cichlidae y Poeciliidae con 12 y 9 especies, respectivamente; ambas poseen el mayor número de especies en el sur de Quintana Roo (Schmitter-Soto y Gamboa-Pérez, 1996), así como en el resto de la provincia ictiogeográfica del Usumacinta (Miller, 1982; Morales-Román y Rodiles-Hernández, 2000; Soria-Barreto y Rodiles-Hernández, 2008; López-López et al., 2009; Miller et al., 2009; Esselman y Allan, 2010).
El 69% de las especies pertenece al componente dulceacuícola y el 31% al componente marino. Las primeras son dominantes en toda la península de Yucatán (Hubbs, 1936; Miller, 1982; Miller, 1991; Miller et al., 2009), mientras que las segundas se han dado a conocer como relativamente escasas en la región del río Hondo (Schmitter-Soto y Gamboa-Pérez, 1996), lo cual coincide con lo encontrado en este trabajo. La baja presencia de especies marinas en el río podría deberse a que éste desemboca en la bahía de Chetumal y no directamente en el mar, por lo que un gran número de especies marinas encuentran en la bahía un hábitat más favorable que la propia región estuarina del río y, por consiguiente, la intrusión de especies marinas en el río es menor.
Oreochromis niloticus fue la única especie exótica recolectada, que junto con O. mossambicus, fue introducida al río en 1988, posiblemente con fines de acuicultura (Schmitter-Soto y Caro, 1997). Recientemente se registró una especie de Pterygoplichthys en el río Bravo, tributario del Hondo (R. Quintana, com. pers.), pero no fue capturada en el cauce principal durante este estudio ni se conocen todavía ejemplares en Quintana Roo. Caranx hippos, Centropomus undecimalis y M. atlanticus han sido registradas como especies comunes en el río Hondo (Schmitter-Soto et al., 2011), aunque en el presente estudio sólo se observó un ejemplar muerto de M. atlanticus. La presencia de Thorichthys aureus registrada por Scholz y Vargas-Vázquez (1998) es dudosa, debido a que no se conocen ejemplares y es difícil distinguirla de T. meeki, que es la especies de Thorichthys común en el río. L. griseus y S. testudineus se dan a conocer por primera vez para el río, aunque ya se conocían de la bahía de Chetumal (Pimentel Cadena, 2001).
Tanto la abundancia numérica como la biomasa mostraron un gradiente en aumento desde La Unión hasta la boca del río. Esto concuerda con lo propuesto por Vannote et al. (1980), quienes explican que la materia orgánica parcialmente procesada que no es utilizada por las comunidades que habitan en la cabecera de los ríos es transportada río abajo. La energía que no es utilizada río arriba, junto con los aportes locales del sistema, es aprovechada por las comunidades que se encuentran en la parte baja del río.
La especie más importante en el aporte de organismos y en biomasa fue P. melanurus, como lo es también en el río Lacanjá (Morales-Román y Rodiles-Hernández, 2000). Esta especie también estuvo presente a lo largo de todo el río Hondo, al igual que en el río Champotón (López-López et al., 2009). Su relevancia en el IVIr, así como su plasticidad demostrada en el ACC respalda la idea de Esselman y Allan (2010), quienes señalan a P. melanurus como una especie altamente exitosa en los ríos de la plataforma yucateca.
La riqueza de especies no mostró un aumento de la cabecera hasta la desembocadura tal y como se ha visto en otros ríos (Edds, 1993; Rodiles-Hernández et al., 1999; Smith et al., 2009; McGarvey, 2011), sino que los valores aumentaron y disminuyeron entre los sitios de muestreo. La Unión tuvo el mayor número de especies, debido a que este sitio se vio influenciado por un arroyo que aportó especies pequeñas o en estado juvenil, mientras que el bajo número de especies en Cocoyol fue resultado de la poca visibilidad dentro del agua, lo que afectó el desempeño del pescador. La ausencia del gradiente esperado también fue expuesta por Rojas y Rodríguez (2008) y Trujillo-Jiménez et al. (2010) en los ríos Grande de Térraba (Costa Rica) y Amacuzac (Morelos), respectivamente. En ambos ríos la influencia de actividades humanas, contaminación, principalmente, estaría provocando la alteración de dicho gradiente. El río Hondo no está exento de este problema, ya que en sus aguas se han detectado hidrocarburos aromáticos policíclicos y plaguicidas organiclorados como consecuencia de la actividad agrícola en la ribera de México y Belice (Álvarez-Legorreta, 2009). Otros estudios han demostrado que el mayor número de especies se encuentra en los sitios cercanos a la boca, debido a la confluencia de especies marinas, estuarinas y dulceacuícolas (Lyons y Schneider, 1990; Gómez-González, 2011), pero en el en río Hondo no fue así. La disminución de especies en los sitios cercanos a la boca se ha asociado con el aumento de los valores de salinidad (Barletta et al., 2005; Sosa-López et al., 2007). Los valores de diversidad y equidad fueron más altos río arriba; este patrón también lo encontraron Smith et al. (2009) en el río Sorocaba, Brasil, como consecuencia de la perturbación del hábitat y deforestación río abajo. En el río Hondo se apreció un mayor grado de conservación en la parte alta, lo cual también ocurre en sus tributarios (Schmitter-Soto et al., 2011). La dominancia numérica de las especies E. plumieri, O. niloticus y principalmente P. melanurus, ayudó a explicar el porqué de los valores bajos de los índices de diversidad.
Mientras que ambientalmente el río muestra una zonificación de 3 partes, en términos de su ictiofauna el río puede dividirse en 2 regiones o conjuntos, tal y como lo expresó el dendrograma con base en la distribución de las especies a lo largo del río. De este modo se aprecia una región media alta (Cocoyol, Sarabia y La Unión) y una región media-baja (boca, Santa Elena y Allende).
A pesar de haber utilizado diversas artes de pesca: redes agalleras, nasas, anzuelos, palangres y arpón, tan sólo se capturaron 19 de 40 especies, ésto debido a diversos factores: 1) la presencia de árboles sumergidos, restos de árboles y ramas que el río Hondo arrastra, no permitieron el correcto funcionamiento de algunas artes como redes y atarrayas; 2) el río ofrece pocos espacios en la ribera como para utilizar chinchorros; 3) parte de los antecedentes (Schmitter-Soto, 1998; Scholz y Vargas-Vázquez, 1998; Esselman, 2009; Schmitter-Soto et al., 2009; Ruiz-Cauich, 2011) registraron varias especies de tallas pequeñas (pecílidos en particular) y parte de este material se encuentra en la colección ictiológica de ECOSUR-Chetumal, por lo que el trabajo se centró en especies de tallas grandes para complementar la información existente; 4) no todas las especies de la lista fueron observadas durante el muestreo; 5) el arpón fue el arte de pesca más efectivo y que nos permitió capturar ejemplares de tallas grandes, cuyos registros eran muy pocos o nulos para algunas especies, dejando de lado las especies pequeñas.
El arpón permitió el registro de tallas mayores en las especies capturadas, así como estimar por primera vez su abundancia relativa, especialmente de aquellas que mantienen la pesquería de subsistencia en el río Hondo (C. urophthalmus, E. plumieri, I. meridionalis, L. griseus, O. niloticus, Paraneetroplus melanurus y Petenia splendida), así como especies grandes que no se consumen por los pobladores (Strongylura timucu y S. testudineus). Por otro lado, las especies pequeñas; especialmente pecílidos, aterínidos, carácidos; quedaron fuera del alcance de este arte, debido al tamaño de los ejemplares. Para poder haber tenido un buen registro de éstas, en éste trabajo, debieron haberse usado otros artes como chinchorritos y electropesca, pero las características de los sitios de muestreo no permitieron el buen uso y efectividad. Asimismo, tanto los antecedentes como la colección ictiológica de ECOSUR-Chetumal, tienen un buen registro de las especies pequeñas, no así para especies de tallas mayores, las cuales se pudieron capturar gracias al uso del arpón.
Es importante señalar que el monitoreo debe incrementarse en el cauce principal del río, tanto en artes de pesca, temporadas del año y esfuerzo de muestreo; lo cual nos permitiría, por ejemplo, prever posibles invasiones de especies exóticas como la del pez diablo (Pterygoplichthys sp.), evaluar el efecto de la presencia de la tilapia (O. niloticus) y la presencia de contaminantes que provienen del cultivo de caña en los márgenes del río. Todos estos factores son un riesgo para los peces que habitan en el río Hondo, especialmente porque el río se encuentra en una región con alto grado de endemismos, especialmente para los géneros Atherinella, Cryptoheros y Thorichthys (Schmitter-Soto y Gamboa-Pérez, 1996). Es por esto, que los esfuerzos de monitoreo deben incrementarse en el cauce principal del río, además de explorar mejor las partes profundas, cuya ictiofauna se desconoce en gran medida.
Finalmente, este trabajo sienta las bases para la futura administración de los recursos acuáticos ícticos con los que el río cuenta, ya que aquí se presenta una lista actualizada de las especies que habitan su cauce, principalmente las de mayor talla, que son las que sostienen la pesquería local en la ribera del río, además de dar una estimación de la proporción en la que se encuentran las especies que se capturaron en campo con el uso del arpón, principal arte de pesca de los pescadores, y la cual debe tomarse con las reservas necesarias al ser un arte de pesca selectiva.
Agradecimientos
Los autores agradecen al Conacyt, al Colegio de la Frontera Sur, a Teresa Álvarez-Legorreta, Adán Enrique Gómez-González, Peter C. Esselman, Rodrigo Pacheco-Díaz, Janneth Adriana Padilla-Saldívar, Edgardo López y Horacio Cárdenas-Puc.
Literatura citada
Álvarez-Legorreta, T. 2009. Contaminación acuática. In El sistema ecológico de la bahía de Chetumal/Corozal: costa occidental del mar Caribe, J. Espinoza-Ávalos, G. A. Islebe y H. A. Hernández-Arana (eds.). El Colegio de la Frontera Sur, Chetumal, Quintana Roo. p. 205-217. [ Links ]
Álvarez-Legorreta, T. 2011. Uso y manejo de recursos hídricos. In Riqueza biológica de Quintana Roo. Un análisis para su conservación. Tomo 1, vol. 1, C. Pozo, N. Armijo-Canto y S. Calmé (eds.). El Colegio de la Frontera Sur, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Gobierno del Estado de Quintana Roo y Programa de Pequeñas Donaciones, México, D. F. p. 121-126. [ Links ]
Arriaga-Cabrera, L., V. Aguilar-Sierra, J. Alcocer-Durán, R. Jiménez-Rosenberg, E. Muñoz-López y E. Vázquez-Domínguez. 1998. Regiones hidrológicas prioritarias: fichas técnicas y mapa (escala 1:4,000,000). Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, D. F. 142 p. [ Links ]
Barletta, M., A. Barletta-Bergan, U. Saint-Paul y G. Hubold. 2005. The role of salinity in structuring the fish assemblages in a tropical estuary. Journal of Fish Biology 66:45-72. [ Links ]
Camarena-Luhrs, T. y L. Cobá-Cetina. 1991. Aguas continentales e hipohalinas. In Estudios ecológicos preliminares de la zona sur de Quintana Roo, T. Camarena-Luhrs y S. Salazar-Vallejo (eds.). CIQRO, Chetumal, Quintana Roo. p. 161-166. [ Links ]
Clarke, K. R. y R. M. Warwick. 2006. Change in marine communities: an approach to statistical interpretation. PRIMER-E Ltd, Plymouth, Devon. 172 p. [ Links ]
De Jesús-Navarrete, A. y Z. V. Góngora-Pech. 2009. Distribución y abundancia de los nematodos en el gradiente salino río Hondo-Bacalar Chico. In El sistema ecológico de la bahía de Chetumal/Corozal: costa occidental del mar Caribe, J. Espinoza-Ávalos, G. A. Islebe y H. A. Hernández-Arana (eds.). El Colegio de la Frontera Sur, Chetumal, Quintana Roo. p. 69-79. [ Links ]
Edds, D. R. 1993. Fish assemblage structure and environmental correlates in Nepal's Gandaki River. Copeia 1993:48-60. [ Links ]
Eschmeyer, W. N. y R. Fricke. 2012. Catalog of Fishes electronic version. http://research.calacademy.org/research/ichthyology/catalog/fishcatmain.asp; última consulta: 28.XII.2012. [ Links ]
Espinosa-Pérez, H., M. T. Gaspar-Dillanes y P. Fuentes-Mata. 1993. Listados faunísticos de México. III. Los peces dulceacuícolas mexicanos. IBUNAM, México, D. F. 98 p. [ Links ]
Esselman, P. C. 2009. Fish communities and conservation of aquatic landscapes in northeastern Mesoamerica. Tesis, The University of Michigan. Ann Arbor, Michigan. 120 p. [ Links ]
Esselman, P. C. y J. D. Allan. 2010. Relative influences of catchment- and reach-scale abiotic factors on freshwater fish communities in rivers of northeastern Mesoamerica. Ecology of Freshwater 19:439-454. [ Links ]
Gamboa-Pérez, H. C. 1991. Ictiofauna dulceacuícola en la zona sur de Quintana Roo. In Estudios ecológicos preliminares de la zona sur de Quintana Roo, T. Camarena-Luhrs y S. Salazar-Vallejo (eds.). CIQRO, Chetumal. p. 186-198. [ Links ]
Gamboa-Pérez, H. C. 1994. Peces continentales de la frontera México-Belice: río Hondo y cuerpos de agua adyacentes. In Estudio integral de la frontera México-Belice. Tomo IV: recursos naturales, E. Suárez-Morales (ed.). CIQRO, Chetumal. p. 143-154. [ Links ]
Gómez-González, A. E. 2011. Comunidad de peces del sistema Chantuto-Panzacola, Reserva de la Biosfera La Encrucijada. Tesis, El Colegio de la Frontera Sur. San Cristóbal de las Casas, Chiapas. 78 p. [ Links ]
Greenfield, D. W. y J. E. Thomerson, 1997. Fishes of the continental waters of Belize. University Press of Florida, Gainesville. 311 p. [ Links ]
Herrera-Sansores, J. C. y J. D. Heredia-Escobedo. 2011. Hidrología superficial. In Riqueza biológica de Quintana Roo. Un análisis para su conservación. Tomo 1, C. Pozo, N. Armijo-Canto y S. Calmé (eds.). El Colegio de la Frontera Sur, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Gobierno del Estado de Quintana Roo y Programa de Pequeñas Donaciones , México, D. F. p. 42-49. [ Links ]
Hubbs, C. L. 1936. Fishes of the Yucatán peninsula. Carnegie Insitute, Washington D. C. 454: 261-295. [ Links ]
López-López, E., J. E. Sedeño-Díaz, F. López-Romero y P. Trujillo-Jiménez. 2009. Spatial and seasonal distribution patterns of fish assemblages in the río Champotón, southeastern Mexico. Reviews in Fish Biology and Fisheries, 19:127-142. [ Links ]
Lyons, J. y D. W. Schneider. 1990. Factors influencing fish distribution and community structure in a small coastal river in southwestern Costa Rica. Hydrobiologia 203:1-14. [ Links ]
Magnon-Basnier, C. 1996. Diagnóstico para el manejo y preservación de la cuenca hidrográfica del río Hondo. Tesis, El Colegio de la Frontera Sur. Chetumal, Quintana Roo. 52 p. [ Links ]
Magurran, A. E. 1988. Ecological diversity and its measurement. Princeton University Press, Princeton, Nueva Jersey. 179. [ Links ]
McGarvey, D. J. 2011. Quantifying ichthyofaunal zonation and species richness along a 2 800-km reach of the río Chama and río Grande (USA). Ecology of Freshwater 20:231-242. [ Links ]
McMahan, C., C. M. Murray, A. D. Geheber, C. D. Boeckman y K. Piller. 2011. Paraneetroplus synspilus is a junior synonym of Paraneetroplus melanurus (Teleostei:Cichlidae). Zootaxa 2833:1-14. [ Links ]
Miller, R. R. 1982. Pisces. In Aquatic biota of Mexico, Central America and the West Indies, S. H. Hulbert y A. Villalobos-Figueroa (eds.). San Diego State University, San Diego. p. 486-501. [ Links ]
Miller, R. R. 1991. Taxonomic status of two nominal species of Mexican freshwater fishes (Cyprinidae). Copeia 1991:225-227. [ Links ]
Miller, R. R., W. L. Minckley y S. M. Norris. 2009. Peces dulceacuícolas de México. Conabio, Sociedad Ictiológica Mexicana, El Colegio de la Frontera Sur, Consejo de Peces del Desierto, México, D. F. p. 559. [ Links ]
Morales-Román, M. y R. Rodiles-Hernández. 2000. Implicaciones de Ctenopharyngodon idella en la comunidad de peces del río Lacanjá, Chiapas. Hidrobiológica 10:13-24. [ Links ]
Nelson, J. S. 2006. Fishes of the world. Fourth edition, John Wiley y Sons, Nueva York. 601 p. [ Links ]
Pacheco-Díaz, R. I. 2011. Efectos del paisaje sobre un índice biótico de integridad en el río Hondo. Tesis, El Colegio de la Frontera Sur. Chetumal, Quintana Roo. 87 p. [ Links ]
Pimentel-Cadena, E. 2001. Distribución y abundancia de la ictiofauna en la bahía de Chetumal, Quintana Roo. Tesis, Instituto Tecnológico del Mar Núm. 6. Bahía de Banderas, Nayarit. 44 p. [ Links ]
Rodiles-Hernández, R., E. Díaz-Pardo y J. Lyons. 1999. Patterns in the species diversity and composition of the fish community of the Lacanja River, Chiapas, Mexico. Journal of Freshwater Ecology 14:455-468. [ Links ]
Rojas, J. R. y O. Rodríguez. 2008. Diversidad y abundancia ictiofaunística del río Grande de Térraba, sur de Costa Rica. Revista de Biología Tropical 56:1429-1447. [ Links ]
Ruiz-Cauich, L. E. 2011. Un modelo trófico para la cuenca del río Hondo, México. Tesis, El Colegio de la Frontera Sur. Chetumal, Quintana Roo. 144 p. [ Links ]
Schmitter-Soto, J. J. 1998. Catálogo de los peces continentales de Quintana Roo. El Colegio de la Frontera Sur, San Cristóbal de las Casas. 239 p. [ Links ]
Schmitter-Soto, J. J. y C. I. Caro. 1997. Distribution of tilapia, Oreochromis mossambicus (Perciformes: Cichlidae), and water body characteristics in Quintana Roo, Mexico. Revista de Biología Tropical 45:1257-1261. [ Links ]
Schmitter-Soto, J. J. y H. Gamboa-Pérez, H. 1996. Composición y distribución de peces continentales en el sur de Quintana Roo, península de Yucatán, México. Revista de Biología Tropical 44:199-212. [ Links ]
Schmitter-Soto, J. J., L. E. Ruiz-Cauich, R. L. Herrera y D. González-Solís. 2011. An index of biotic integrity for shallow streams of the Hondo river basin, Yucatán peninsula. Science of the Total Environment 409:844-852. [ Links ]
Schmitter-Soto, J. J., L. Vásquez-Yeomans, E. Pimentel-Cadena, R. Herrera-Pavón, G. Paz y N. García-Téllez. 2009. Peces. In El sistema ecológico de la bahía de Chetumal/Corozal: costa occidental del mar Caribe, J. Espinoza-Ávalos, G. A. Islebe y H. A. Hernández-Arana (eds.). El Colegio de la Frontera Sur, Chetumal. p. 102-114. [ Links ]
Scholz, T. y J. Vargas-Vázquez. 1998. Trematodes from fishes of the Hondo river and freswater lakes of Quintana Roo, Mexico. Journal of the Helminthological Society of Washington 65:91-95. [ Links ]
Smith, W. S., M. Jr. Petrere y W. Barrella. 2009. The fish community of the Sorocaba river basin in different habitats (state of São Paulo, Brazil). Brazilian Journal of Biology 69:1015-1025. [ Links ]
Soria-Barreto, M. y R. Rodiles-Hernández. 2008. Spatial distribution of cichlids in Tzendales river, Biosphere Reserve Montes Azules, Chiapas, Mexico. Environmental Biology of Fishes 83:459-469. [ Links ]
Sosa-López, A., D. Mouillot, J. Ramos-Miranda, D. Flores-Hernández y T. Do-Chi. 2007. Fish species richness decreases with salinity in tropical coastal lagoons. Journal of Biogeography 34:57-61. [ Links ]
Ter-Braak, C. J. F. y P. Smilauer. 2002. Canoco reference manual and CanoDraw for Windows user's guide: software for canonical community ordination. [4.5]. 2002. Microcomputer Power. [ Links ]
Trujillo-Jiménez, P., E. López-López, E. Díaz-Pardo y J. A. Camargo. 2010. Patterns in the distribution of fish assemblages in río Amacuzac, Mexico: influence of abiotic factors and biotic factors. Reviews in Fish Biology and Fisheries 20:457-469. [ Links ]
Vannote, R. L., G. W. Minshall, K. W. Cummins, J. R. Sedell y C. E. Cushing. 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Science 37:130-137. [ Links ]

