










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de biodiversidad
versión On-line ISSN 2007-8706versión impresa ISSN 1870-3453
Rev. Mex. Biodiv. vol.85 no.2 México jun. 2014
https://doi.org/10.7550/rmb.34996
Manejo y aprovechamiento de recursos
Crecimiento de plantas jóvenes de Cordia dodecandra (Boraginaceae) en tres etapas sucesionales de vegetación en Calotmul, Yucatán
Growth of young plants of Cordia dodecandra (Boraginaceae) in three successional stages of vegetation in Calotmul, Yucatán
Carlos Yam-Chin1, Patricia Montañez-Escalante2* y Rocío Ruenes-Morales2
1 Instituto Tecnológico de Conkal. Km 16.3 antigua carretera Mérida-Motul, 97345 Mérida, Yucatán, México.
2 Facultad de Medicina Veterinaria y Zootecnia. Campus de Ciencias Biológicas y Agropecuarias. Universidad Autónoma de Yucatán. Km 15.5 Carretera Mérida-Xmatkuil, 97315 Mérida, Yucatán, México. * montanez@uady.mx
Recibido: 08 enero 2013
Aceptado: 03 diciembre 2013
Resumen
Las poblaciones naturales de Cordia dodencandra, siricote, en Yucatán se encuentran amenazadas por la sobreexplotación de su madera, apreciada por su dureza y veteado. Se requiere diseñar programas de reforestación y enriquecimiento de la vegetación en áreas perturbadas que consideren el crecimiento de esta especie en respuesta a los factores ambientales de selvas en diferentes grados de recuperación. El objetivo de este trabajo fue caracterizar las condiciones ambientales de temperatura, humedad y radiación fotosintéticamente activa en diferentes etapas sucesionales de vegetación: avanzada, media, temprana y sin vegetación, para determinar su influencia sobre el área foliar específica y el crecimiento de plantas jóvenes de siricote. En cada etapa sucesional se estableció una parcela con 60 plántulas de siricote, a distancias de 4 m entre línea de siembra y 2.7 m entre plantas. Después de 19 meses, los sitios en sucesión temprana son los más adecuados para el crecimiento de siricote. Existe una relación inversamente proporcional entre la incidencia de luz y el área foliar específica y una relación directamente proporcional entre humedad relativa y el área foliar específica. Se concluye que el siricote puede utilizarse en programas de enriquecimiento de la vegetación avanzada y para reforestar áreas en sucesión temprana.
Palabras claves: interacciones, microambiente, radiación.
Abstract
Natural populations of Cordia dodencandra (siricote) in Yucatán are threatened due to the overexploitation of their wood, valued by its hardness and grain. Reforestation, enrichment of vegetation in disturbed areas and commercial plantation programs of this species are needed. These programs must take into account the relation between plant growth and its response to the environmental factors occurring in forests in different stages of recovery. The aim of this study was to characterize the environmental conditions; temperature, humidity and photosynthetically active radiation; under different successional stages; advanced, intermediate, early and without vegetation; in order to determine the influence of specific leaf area and growth of siricote seedlings. On a patch of each successional stage, sixty seedlings of siricote were established in a plot. Distribution of seedlings was 4m between rows and 2.7 between plants. After 19 months, the early successional plot was the better site for siricote growth. We observed an inverse relationship between photosynthetically active radiation and specific leaf area and, a direct relationship between relative humidity and specific leaf area. According to our results siricote could be used in programs of enrichment of vegetation in advanced successional stages as well as for reforesting early successional areas.
Key words: interactions, microenvironment, radiation.
Introducción
La sucesión secundaria se caracteriza por cambios en la estructura y composición de la vegetación a través del tiempo producida por la acción de las actividades antropogénicas o los fenómenos naturales (Sánchez et al., 2007; Dahdouh-Guebas et al., 2002). Estos cambios modifican el régimen de luz, temperatura y humedad que penetran en el sistema y generan microclimas que afectan las funciones básicas de las plantas como la fotosíntesis y el crecimiento, esto puede favorecer las interacciones positivas y la incorporación de especies en los estratos inferiores (Gehlhausen et al., 2000; Sherman et al., 2005; Martínez-Ramos y García-Orth, 2007; Lambers et al., 2008; Mendoza-Arroyo et al., 2011).
La radiación solar, además de ejercer influencia sobre la temperatura y la humedad presentes en un ecosistema, se le considera como el factor más importante para el crecimiento y desarrollo de una planta (Jaramillo-Robledo, 2005). Por lo general, la ausencia o el exceso generan cambios morfológicos en la hoja, sobre todo, en el grosor y en el área foliar (Maxwell et al., 1999; Close et al., 2009; Cruz-Rodríguez y López-Mata, 2010). Estos cambios pueden manifestarse en las plantas durante periodos continuos o estacionales de crecimiento, dependiendo de la intensidad lumínica que reciban las plantas (Lavender, 1984; Chapin et al., 1987; Puntieri, 2005).
Varios estudios demuestran el efecto de la cantidad de luz sobre el crecimiento de especies vegetales. Romo (2005), en plántulas de Dipterix micrantha, obtuvo mayor crecimiento en plantaciones mixtas con sombra que en claros. Moreno (1998) realizó estudios con el sajo (Capnosperma panamensis) a través de luz controlada con sombras de 10, 32 y 45%, obteniendo mayor crecimiento donde la intensidad lumínica era baja en comparación con otras parcelas donde las plantas se encontraban más expuestas a la luz.
El grado de tolerancia de las especies forestales a los factores ambientales es uno de los criterios más importantes a considerar en las actividades relacionadas con la recuperación de la selva, en los procesos de reforestación y en el enriquecimiento de la vegetación nativa (Khurana y Sing, 2001; Hau y Corlett, 2003; Rao et al., 2008). El enriquecimiento de la vegetación es una estrategia común utilizada en la recuperación de las selvas, aprovechando los espacios generados por la dinámica natural de la selva para incorporar especies nativas de valor comercial, como fuentes de productos maderables y de servicios ambientales (Zheng et al., 2000; Lozada et al., 2003; Souza y Valio, 2003; Griscom et al., 2005; Khumbongmayum et al., 2005).
Por lo anterior, es importante conocer cómo responden las plantas a las diferentes condiciones ambientales que se presentan durante los procesos de recuperación de las selvas, considerando que hay una relación muy estrecha entre el crecimiento y los factores ambientales (Harold y Hocker, 1984; Montgomery y Chazdon, 2002; López-Torres y Tamarit-Urias, 2005; Ekinci, 2006). El objetivo de este estudio fue caracterizar las condiciones ambientales en términos de temperatura, humedad y radiación fotosintéticamente activa en parcelas de selva en diferentes etapas sucesionales, para evaluar su influencia sobre el área foliar específica y el crecimiento en altura, así como el diámetro basal de plantas jóvenes de C. dodecandra (ciricote). Esta especie es apreciada por la dureza y el veteado de su madera. Se distribuye en el sur de México (Pennington y Sarukhán, 2005), y se encuentra como parte de la agrodiversidad de los huertos familiares en la comunidades rurales (Benjamín et al., 2001; Moreno-Casasola y Paradowska, 2009; Zamora et al., 2009), también se puede encontrar poblaciones naturales en selvas medianas caducifolias y sudcaducifolias. Sin embargo, sus poblaciones se encuentra amenazadas por la sobreexplotación (Forster et al., 2002). Este estudio generará información sobre las condiciones adecuadas para el crecimiento de C. dodecandra en programas de reforestación, enriquecimiento de la vegetación en áreas perturbadas y plantaciones forestales comerciales.
Materiales y métodos
Área de estudio. El estudio se realizó en la finca forestal Chechen, localizado en el km 32 carretera Calotmul-Valladolid, municipio de Calotmul, Yucatán, México. El clima de la región está clasificado como cálido subhúmedo, con periodos de lluvias (junio a octubre), nortes (noviembre-enero) y de sequía (marzo-mayo). La temperatura media anual es de 26.3° C y la precipitación pluvial de 1 200 mm anuales (García, 1988).
El área se encuentra a una altitud promedio de 15 m snm. Su fisiografía es plana, constituida por una llanura de barrera, con piso rocoso escarpado con ligeros lomeríos, conformados por montículos de piedra (Duch, 1991). Los suelos son del tipo cambisol y litosol (Bautista et al., 2005). Pennington y Sarukhán (2005) clasifican la vegetación de la zona como selva mediana subcaducifolia.
Cordia dodecandra-establecimiento. Las semillas de ciricote se obtuvieron de un banco semillero (Rancho Montara) en el municipio de Tzucacab, Yucatán. Las cuales se pusieron a germinar en la primera semana de agosto del 2009, 2 meses después se seleccionaron aquellas plántulas con altura de 20 cm y se implantaron en parcelas de 20 × 50 m. Fueron 60 individuos por parcela con un diseño de 4 m entre línea de siembra y 2.7 m entre plantas, con dirección este-oeste. Se realizaron deshierbes frecuentes durante el periodo de lluvias para favorecer el establecimiento.
Los tratamientos fueron parcelas en: 1) sucesión temprana (ST), compuestas de vegetación herbácea y arbustiva; 2) sucesión media (SM), de 5 -10 años con vegetación entre los 3 a 8 m de altura; 3) sucesión avanzada (SA), de 20 a 30 años con vegetación entre 10 a 15 m de altura y 4) sin vegetación (SV) plantación en monocultivo de C. dodecandra con control permanente de arvenses.
Caracterización de la vegetación. Se caracterizó la vegetación presente en cada estadio sucesional trazando 4 cuadrantes de 20 × 50 m (0.1 ha), se identificaron las especies con diámetro a la altura del pecho (DAP) ≥ 4cm, se les midió la altura total, DAP y cobertura. Para el arbolado con DAP < 4 cm se tomaron datos de número de individuos en subparcelas anidadas de 10 × 10 m. Para la parcela con sucesión temprana sólo se tomaron datos de las especies presentes y su altura, utilizando parcelas de 10 × 10 m. La composición y estructura se obtuvo a partir de los valores de importancia (IVI) (Zarco-Espinoza et al., 2010):
IVI= DeR + DoR + FR
donde DeR= densidad relativa, DoR= dominancia relativa, FR= frecuencia relativa.
Variables ambientales. Se midió la radiación fotosintéticamente activa (RFA, µmol m−2 s−1) con un ceptómetro (ACCUPAR LP-80, Decagon, Pullman, USA) a 1 m de altura sobre 30 plantas seleccionadas al azar en cada tratamiento; se realizaron 3 mediciones por día en horarios de 9:00 a 10:00, de 12:00 a 13:00 y de 15:00 a 16:00 h. Se registró la temperatura (°C) y la humedad relativa atmosférica (%) con sondas acopladas a un registrador de datos (HOBOu12, Onset, Pocasset, USA) cada 30 minutos durante 24 h por cada tratamiento. Todas las variables ambientales fueron registradas cada 30 días durante 8 meses, agosto de 2010 a marzo de 2011. Los datos fueron agrupados de acuerdo con los 3 periodos presentes en la zona: de lluvias (agosto-octubre), nortes (noviembre-enero) y sequía (marzo-abril).
Crecimiento de plantas de ciricote. A los 9 meses después de iniciado el experimento, en cada tratamiento se seleccionaron al azar 30 plantas de C. dodecandra a las cuales se les midió el crecimiento en altura (cm) (del suelo al ápice) y en diámetro basal a 3 cm del suelo. Las mediciones se realizaron en intervalos de 30 días entre cada muestreo, por un periodo de 10 meses (junio 2010 a marzo de 2011) con la intención de abarcar el periodo de lluvias, nortes y sequías
Área foliar específica (AFE). En septiembre de 2010 se registró el área foliar específica (AFE cm−2 g−1), se colectaron 10 hojas por planta en una muestra al azar de 10 individuos por tratamiento. El área foliar (AF) se registró con un medidor de área foliar en banda (LI 30, LI-COR Lincoln, Nebraska, USA). Las hojas fueron secadas a una temperatura constante de 72° C por 24 h y después se calculó el AFE: área de la hoja (cm2)/ g de peso seco.
Análisis estadístico. Para la evaluación del crecimiento de las plantas se utilizó un modelo de análisis de varianza (ANOVA) con medidas repetidas, y se aplicó la prueba de homogeneidad de varianzas de Levene (Correa et al., 2006). Se realizaron transformaciones logarítmicas de los datos de altura, diámetro y área foliar específica para lograr la normalidad, y se utilizó la prueba de Tukey para la comparación de medias (Montgomery, 1991). Para determinar la variación de la RFA, la temperatura y humedad relativa atmosférica en cada tratamiento, se realizaron análisis de varianza de una sola vía con una prueba de Tukey. Se aplicó un análisis de regresión múltiple para determinar la relación entre las variables ambientales y el crecimiento en altura y diámetro basal. Para la temperatura y la humedad se utilizaron los datos registrados de 6 a las 18 h, ya que demostraron ser más informativos al momento de correr el análisis de regresión. Para todos los análisis estadísticos se utilizó el software Statistica Six Sigma, versión 7, Stat Soft, USA.
Resultados
Estructura de la vegetación. En la sucesión temprana (ST) se registraron 19 especies de 9 familias botánicas entre arbustos, herbáceas y plantas juveniles de especies arbóreas con alturas entre 0.9 y 1.5 m. La especie con mayor dominancia por su cobertura fue el pasto Brachiaria brizantha, estimada entre 60 y 70%. Las especies con mayor valor de importancia fueron Neomillspaugia emarginata, Gymnopodium floribundum y Mimosa bahamensis (Cuadro 1).
En el estadio sucesional medio (SM) se registraron 13 familias botánicas con 26 especies arbóreas, la altura promedio fue de 5.7±1.2 m. Las especies con mayor valor de importancia fueron: Gymnopodium floribundum, Bourreria pulchra, Havardia albicans y Mimosa bahamensis (Cuadro 1). Entre el arbolado joven < 4 cm de DAP se registraron 17 especies, las más abundantes fueron Gymnopdium floribundum, Lysiloma latisiliquum, Piscidia piscipula y Croton glabellus.
En la vegetación con estadio sucesional avanzado (SA) se registraron 14 familias con 30 especies arbóreas con diámetros ≥ 4 cm y altura de 8.7±1.8 m. Destacan por su valor de importancia Caesalpinia gaumeri, Lysiloma latisiquum, Bursera simaruba y Gymnopdium floribundum (Cuadro 1). El arbolado joven con diámetros < 4 cm, estuvo compuesto por 23 especies, las más abundantes fueron: Caesalpinia gaumeri, Eugenia sp. y Thouinia paucidentata.
Crecimiento de C. dodecandra. A los 19 meses del implante, el mayor crecimiento en altura se registró en las parcelas de SV (173.88±43 cm y 3.6±0.7 cm de diámetro). El mayor crecimiento en diámetro se presentó en el tratamiento de ST (3.3±2 cm y 148.3±49 cm en altura). El tratamiento de SM registró 121.4±47 cm en altura y 1.8±0.5 cm en diámetro, mientras que el tratamiento de SA, 206.3±80 cm de altura y 2.5±0.5 cm de diámetro (Fig. 1).

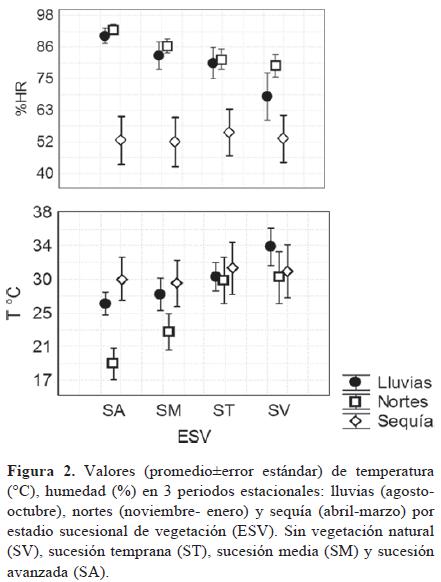
Temperatura y humedad relativa. Las condiciones ambientales fueron estadísticamente diferentes entre tratamiento en los periodos de lluvia y nortes (p> 0.01), mientras que durante la sequía no se encontraron diferencias significativas (Fig. 2). La temperaturas más altas se registraron en el tratamiento control sin vegetación (SV) con 35.59±1.7° C (media±error estándar) para el periodo de lluvias y 29.74±1.5° C para los nortes, y el más bajo fue en el tratamiento de sucesión avanzada (SA) con temperaturas de 26.49±0.06° C en lluvias y 18.94±0.9° C en el periodo de nortes (p> 0.01). La humedad relativa más alta se presentó en SA con un 90.41±1.3% en la temporada de lluvias, y en el periodo de secas la humedad no presentó diferencias significativas entre los tratamientos (p> 0.05).
Radiación fotosintéticamente activa (RFA). La cantidad de luz fue diferente entre los tratamientos (p> 0.01; Fig. 3). Como se esperaba, la mayor radiación incidente se registró durante la estación seca en la parcela control (SV), seguida por el tratamiento ST.

Área foliar específica. Las plantas de los tratamientos SA y SM presentaron mayor área foliar específica que en los tratamientos ST y SV (144.4±5.03 y 149.06±7.4 cm vs. 106±5.38 y 98.16±5.38 cm, respectivamente (p< 0.01; Fig. 4).

Análisis de regresión múltiple. Las variables asociadas con el crecimiento en altura fueron el estado sucesional de vegetación (ESV) y la radiación fotosintéticamente activa (RFA), con valores de R2 de 85 y 86%, respectivamente (p< 0.01). Para el crecimiento en diámetro basal la interacción con la RFA el valor de la R2 sólo explica el 27% (p< 0.05), la temperatura el 29% (p< 0.05) y la humedad el 86% (p< 0.01). Respecto al AFE, la única variable asociada es la humedad relativa con una R2 de 86% (p< 0.05).
Discusión
Estructura y composición florística. El análisis de vegetación demuestra que la familia Fabaceae es la más representativa en todos los estadios sucesionales de la selva estudiada en Yucatán, y concuerda con lo mencionado por Palacio et al. (2002). En la sucesión temprana, la vegetación está compuesta en su mayor parte por pasto (Bracharia brizantha), además de herbáceas y arbustos. Presentó especies que se encuentran en la sucesión media y temprana, lo que indica que existe una dinámica sucesional que permite la incorporación de especies pioneras (Neomillspaughia emarginata, Mimosa bahamensis, Coccoloba spicata) y en menor densidad especies tolerantes a la sombra (Caesalpinia gaumeri y Piscidia piscipula). Esto puede explicarse con base en el modelo de sucesión por facilitación, el cual propone que las especies pioneras juegan un papel importante en la modificación del hábitat, haciéndolo más adecuado para la incorporación de especies de estadios sucesionales más avanzados (Bush y Van, 1995; Zahawi y Augspurger, 1999; Wrigth, 2002, Guevara, 2005).
En la sucesión media la composición florística está integrada por Gymnopodium floribundum, Mimosa bahamensis, Havardia albicans, Acacia gaumeri, consideradas como especies pioneras, presentando, además, densidades altas de árboles jóvenes con DAP< 4 cm. La presencia de hemisciófitas, por ejemplo: Caesalpinia gaumeri, Lysiloma latisiliquum y Piscidia piscipula, en el arbolado joven indica que la vegetación se encuentra en un proceso de desplazamiento de las especies pioneras. Esta dinámica sucesional viene acompañada de múltiples respuestas de las especies que inciden sobre el crecimiento y desarrollo de otros organismos que interactúan con ellas (Fotelli et al., 2001; Wright, 2002; Wacker et al., 2009).
La estructura y composición florística de la vegetación avanzada se encuentra dominada por el estrato arbóreo compuesto por Caesalpinia gaumeri, Lysiloma latisiliquum y Bursera simaruba, especies de sol, pero tolerantes a la sombra (Sánchez et al., 2007). Gymnopodium floribundum ocupa el cuarto lugar en valor de importancia, Jiménez et al. (2010) y Mendoza-Arroyo et al. (2011) la consideran como un elemento importante en las primeras etapas de la sucesión secundaria y mencionan que al encontrarse en vegetación avanzada es un indicador de perturbación. La baja diversidad de especies encontradas en la sucesión avanzada del área de estudio sugieren que en los estadios sucesionales más avanzados se ha dado un manejo selectivo de especies, debido a las prácticas agrícolas tradicionales de la zona (Toledo et al., 2008).
Condiciones ambientales. En los estadios avanzado (SA) y medio (SM), a pesar de las diferencias en estructura y composición de especies, la cantidad de radiación fotosintética que llega al sotobosque es similar. En la sucesión temprana, debido a la ausencia del estrato arbóreo, la RFA aumentó considerablemente. Los diferentes niveles de RFA, debido a la diferencia en cobertura en los estadios sucesionales ayuda a explicar las diferencias en composición y densidad de especies (Martínez-Ramos y García-Orth, 2007; Sánchez et al., 2007)
Las diferencias encontradas en la temperatura y humedad entre los estadios sucesionales demuestran la importancia de la vegetación en la generación de microambientes, sobre todo, en la conservación del agua atmosférica en el sistema (Joffre y Rambal, 1993; Rincón y Huante, 1993; Gamon et al., 1995; Wirth, 2001; Ceccon et al., 2006). Sun et al. (2008) mencionan que la estructura y la composición de la vegetación se encuentran estrechamente relacionadas con las características ambientales que regulan la evapotranspiración y la disponibilidad de agua para las plantas. La altura del estrato arbóreo crea un gradiente que ayuda a conservar la humedad en el ambiente y mantiene la temperatura más baja a diferencia de la sucesión media en donde el estrato arbóreo es más bajo. En la sucesión temprana (ST) las características de la vegetación permiten que la radiación solar llegue directamente hacia el nivel del suelo, traduciéndose en temperaturas más altas y reducción en la humedad ambiental (Davies-Colley et al., 2000). A pesar de los microambientes más estresantes para las plantas de los sitios de sucesión temprana, en contraste con los de sitios de sucesión más avanzada (Pausas y Austin, 2001), son los sitios utilizados más comúnmente en procesos de reforestación y restauración en los que se utilizan especies locales (Shankar et al., 2009).
Cordia dodecandra-crecimiento. El crecimiento en altura y diámetro de C. dodecandra establecidos en diferentes estadios sucesionales registró diferencias atribuibles a la interacción con la vegetación y sus condiciones ambientales prevalecientes. La plantación de C. dodecandra en la parcela sin vegetación natural (SV) con estricto control de arvenses presentó un crecimiento en altura y diámetro basal (173.8±7.8 cm y 3.7±0.20 cm, respectivamente) en plantas de 19 meses de edad mayores a los obtenidos por Reuter et al. (2008). Ellos documentaron que para plantas de ciricote de 21 meses se alcanzaron alturas de 80 cm en promedio y 1.64 cm de diámetro, en parcelas experimentales establecidas al norte de Yucatán con diferentes niveles de irrigación. En la zona norte de Veracruz, Montero (2009) obtuvo plantas con una altura promedio de 87.61 cm en 11 meses bajo condiciones de monocultivo. Rodríguez et al. (2009) registró alturas de 102 cm y diámetros de 2.8 cm en plantas de 2 años de edad en plantaciones mixtas con otras especies forestales.
Las plantas de C. dodecandra se establecieron sin problemas en el tratamiento de sucesión temprana (ST), aun con la dominancia del pasto Bracharia brizantha. Dañobeytia et al. (2007) encontraron que C. pentandra, Schizolobium parahyba y Swietenia macrophylla pueden establecerse en pastizales abandonados. De manera similar C. dodecandra ha demostrado ser una especie tolerante a las condiciones presentes en la sucesión temprana. Mendoza, et al. (1995) determinaron que en condiciones naturales los individuos juveniles de esta especie sólo se encuentran en los claros de las selvas bajas y medianas caducifolias, lo que sugiere que es una planta de sol, poco tolerante a la sombra.
Las especies leñosas pueden presentar múltiples adaptaciones en respuesta al estrés ambiental. Por ejemplo, el AFE se reduce como una estrategia de la planta para reducir la pérdida de agua por transpiración cuando la humedad atmosférica es baja (Milla et al., 2007; Xu et al., 2009). Además, el aumento en la humedad relativa asociada con la reducción en RFA y temperatura en el estadio en sucesión avanzada (SA) favorece el mayor crecimiento de C. dodecandra. Similarmente, microambientes con menor RFA y mayor humedad relativa también favorecen el crecimiento de diversas plántulas de árboles tropicales (Ekta y Singh, 2001) y de especies de clima templado como las del género Eucalyptus (Guarnaschelli et al., 2009). La intercepción de luz por el estrato arbóreo, estimula a las plantas a producir hojas con mayor área foliar que permiten interceptar más luz, aumentar la tasa fotosintética y a su vez, aumentar la tasa de crecimiento (Lambers et al., 2008).
En este estudio, los datos sugieren que los sitios en sucesión temprana son los más adecuados para el crecimiento y desarrollo de C. dodecandra. Sin embargo, para fines de producción, es necesario hacer análisis de volumen y densidad de madera en un periodo de tiempo mayor. Existe una relación inversamente proporcional entre la incidencia de luz y el área foliar específica, y una relación directamente proporcional entre la humedad relativa y el área foliar específica. Debido a su plasticidad en la tolerancia a diversos factores ambientales, C. dodecandra puede utilizarse en programas de enriquecimiento de la vegetación avanzada y en la reforestación y restauración de áreas con sucesión temprana.
Literatura citada
Bautista, F., D. Palacio-López y W. Huchin-Malta. 2005. Actualización de la clasificación de suelos del estadio de Yucatán. In Caracterización y manejo de suelos de la península de Yucatán, F. Bautista y A. Palacios (eds.). Ed. Instituto Nacional de Ecología. p. 105-202. [ Links ]
Benjamín, T., P. Montañez, J. Jiménez y A. Gillespie. 2001. Carbon, water and nutrients flux in Maya homegardens in the Yucatán peninsula of Mexico. Agroforestry Systems 53:103-111. [ Links ]
Bush, J. K. y O. W. Van. 1995. Interactions between seedlings of an early and late successional woody species. The Southwestern Naturalist 40:379-387. [ Links ]
Ceccon, E., P. Huante y E. Rincon. 2006. Abiotic factors influencing tropical dry forest regeneration. Brazilian Archives of Biology and Technology 49:305-312. [ Links ]
Chapin, F. S., J. Bioom, B. Field y R. Waring. 1987. Plant responses to multiples factors, physiological ecology provides tools studying how interacting environmental resources control plant growth. BioScience 37:49-59. [ Links ]
Close, D., K. Ruthrof, S. Turner, D. Rokich y K. Dixon. 2009. Ecophysiology of species with distinct leaf morphologies: effects of plastic and shadecloth tree guards. Restoration Ecology 17:33-41. [ Links ]
Correa, J. C., R. Iral y L. Rojas. 2006. Estudio de potencia de pruebas de homogeneidad de varianza. Revista Colombiana de Estadística 29:57-76. [ Links ]
Cruz-Rodríguez, J. A. y L. López-Mata. 2010. Cambios ontogénicos en la morfología de plántulas de Manikara zapota: análisis de sus implicaciones ecológicas. Revista Mexicana de Biodiversidad 81:81-86. [ Links ]
Dahdouh-Guebas, F., J. G. Kairo, L. P. Jayatissa, S. Cannicci y N. Koed. 2002. An ordination study to view vegetation structure dynamics in disturbed and undisturbed mangrove forest in Kenya and Sri Lanka. Plant Ecology 162:123-135. [ Links ]
Dañobeytia, F., S. Levy, H. Perales, N. Ramírez, D. Douterlungne y S. López. 2007. Establecimiento de seis especies nativas en un pastizal degradado en la selva lacandona, Chiapas, México. Ecología Aplicada 6:1-8. [ Links ]
Davies-Colley, R., G. W. Payne y M. Van Elswijk. 2000. Microclimate gradients across a forest edge. New Zealand Ecological Society 24:111-121. [ Links ]
Duch, J. 1991. Fisiografía del estado de Yucatán, su relación con la agricultura. Universidad Autónoma de Chapingo, Chapingo. p. 121-122. [ Links ]
Ekinci, H. 2006. Effect of forest fire on some physical, chemical and biological properties soil in Canakkale, Turkey. International Journal of Agriculture and Biology 1:102-106. [ Links ]
Ekta, K. y J. S. Singh. 2001. Ecology of tree seed and seedlings: Implications for tropical forest conservation and restoration. Current Science 80:748-757. [ Links ]
Fotelli, M., A. Gebler, A. Peuke y H. Rennenberg. 2001. Drought affects the competitive interactions between Fagus sylvatica seedlings and an early successional species, Rubus fruticosus: responses of growth, water status and δ13 composition. New Phytologist 151:427- 435. [ Links ]
Forster, R., H. Albrecht, M. Belisle, A. Caballero, H. Galletti, O. Lacayo, S. Ortiz y D. Robinson. 2002. Comunidades forestales y mercadeo de maderas tropicales poco utilizadas de Mesoamérica. Ducere, S. A. México, D. F. p. 65. [ Links ]
Gamon, J. A., C. B. Filed, M. L. Guoulden, K. L. Griffin, G. Hartley, A. E. Joel, J. Peñuelas y R. Valentini. 1995. Relationships between NDVI, canopy structure, and photosynthesis in three Californian vegetation types. Ecological Applications 5:28-41. [ Links ]
García, E. 1988. Modificaciones al sistema de clasificación climática de Köpepen. Cuarta edición. Instituto de Geografía. Universidad Nacional Autónoma de México, México, D. F. p. 9-20. [ Links ]
Gehlhausen, S., M. W. Schwartz y C. K. Augspurger. 2000. Vegetation and microclimatic edge effects in two mixed-mesophytic forest fragments. Plant Ecology 147:21-35. [ Links ]
Griscom, H., P. Ashton y G. Berlin. 2005. Seedlings survival and growth of native tree species in pastures: implications for dry tropical forest rehabilitation in central Panama. Forest Ecology and Management 218:396-318. [ Links ]
Guarnaschelli, A., A. Mantese y A. Battaglia. 2009. Cambios anatómicos y morfológicos en plántulas de Eucalyptus sp. en respuesta a diferentes niveles de luz. Boletín de la Sociedad Argentina de Botánica 44:273-285. [ Links ]
Guevara, S., J. Laborde, y G. Sánchez-Ríos. 2005. Los árboles que la selva dejó atrás. INCI 30:595-601. http://www.scielo.org.ve/scielo.php?script=sci_arttext&pid=S03781844200501000004&lng=es&nrm=iso ISSN 0378-1844; última consulta: 25.VI.2012. [ Links ]
Hau, B. y R. Corlett. 2003. Factors affecting the early survival and growth of native tree seedlings planted on a degraded Hillside Grassland in Hong Kong, China. Restoration Ecology 11:483-488. [ Links ]
Harold, W. y J. R. Hocker. 1984. Introducción a la biología forestal. AGT. Editor. Mexico, D. F. p. 179-226. [ Links ]
Jaramillo-Robledo, A. 2005. La redistribución de la radiación solar y la lluvia dentro de las plantaciones de café (Coffea arabica L.). Revista Académica Colombiana de Ciencia 29:371-382. [ Links ]
Jiménez, J., R. Durán, J. Dupuy y J. González-Iturbe. 2010. Uso del suelo y vegetación secundaria. In Biodiversidad y desarrollo humano en Yucatán, R. Durán y M. Méndez (eds.). CICY, PPD-FMAM, Conabio, Seduma, Mérida. p. 460-464. [ Links ]
Joffre, R. y S. Rambal. 1993. How tree cover influences the water balance of mediterranean rangelands. Ecological Society of America 74:570-582. [ Links ]
Khurana, E. y J. S. Sing. 2001. Ecology of tree seed and seedlings: implications for tropical forest conservation and restoration. Current Science. 80:748-757. [ Links ]
Khumbongmayum, A. D., M. L. Khany y R. S. Tripathi. 2005. Survival and growth of seedlings of a few tree species in the four sacred groves of Manipur, Northeast India. Current Sciencie 88:1781-1788. [ Links ]
Lambers, H., F. S. Chapin III y T. L. Pons. 2008. Plant physiological ecology. Springer, New York. 610 p. [ Links ]
Lavender, D. P. 1984. Plant physiology and nursery environment: interactions affecting seedling growth. In Duryea, L. Mary y Thomas D. Landis (eds.). Forest nursery manual: production of bare root seedlings. Research Laboratory, Oregon State University. Corvallis. 133-141 p. [ Links ]
López-Torres, J. L. y J. C Tamarit-Urias. 2005. Crecimiento e incremento en diámetro de Lysiloma latisiliquum (L.) Benth. En bosques secundarios en Escárcega, Campeche, México. Revista Chapingo. Serie Ciencias Forestales y del Ambiente 9:117-123. [ Links ]
Lozada, J., J. Moreno y R. Suescun. 2003. Plantaciones en fajas de enriquecimiento. Experiencias en 4 unidades de manejo forestal de la Guayana Venezolana. Interciencia 28:568-575. [ Links ]
Martínez-Ramos, M. y X. García-Orth. 2007. Sucesión Ecológica y restauración de las selvas húmedas. Boletín de la Sociedad Botánica de México 80:69-84. [ Links ]
Maxwell, K., J. Marrison, R. Leech, H. Griffiths y P. Horton.1999. Chloroplast acclimation in leaves of Guzmania monostachi in response to high light. Plant Physiology 121:89-95. [ Links ]
Mendoza, P., E. García, J. Aguirre y E. Hernández. 1995. Regeneración natural de especies arbóreas en una selva mediana subperennifolia perturbada por extracción forestal. Acta Botánica Mexicana 32:11-23. [ Links ]
Mendoza-Arroyo, G., A. Navarro-Martínez, S. Ochoa-Ganoa y H. Wal. 2011. Arquitectura arbórea y desarrollo del bosque de Guaiacum sanctum L. en Balam-Kin, Campeche, México. Revista Chapingo. Serie Ciencias Forestales y del Ambiente 17:215-224. [ Links ]
Milla, R., P. B. Reich, Ü. Niinemets y P. Castro-Diez. 2007. Environmental and developmental controls on specific leaf area are little modified by leaf allometry. Functional Ecology 22:265-576. [ Links ]
Montgomery, D. C. 1991. Design and analysis of experiments. 3a. ed. John Wiley and Sons (WIE). USA. 668 p. [ Links ]
Montgomery, R. A. y R. L. Chazdon. 2002. Light gradient partitioning by tropical tree seedlings in the absence of canopy gaps. Oecologia 131:165-174. [ Links ]
Montero, F. M. 2009. Crecimiento inicial de especies arbóreas multipropósito en un terreno ganadero del norte de Veracruz. Tesis de maestría, Colegio de Posgraduados. Montecillo-Texcoco. 61 p. [ Links ]
Moreno, F. 1998. Crecimiento en plántulas de Sajo (Campnosperma panamensis) y Cuangare (Otoba gracilipes) bajo diferentes ambientes lumínicos. Crónica forestal y del medio ambiente 13:71-88. [ Links ]
Moreno-Casasola, P. y K. Paradowska. 2009. Especies útiles de la selva baja caducifolia en las dunas costeras del centro de Veracruz. Madera y Bosques 15:21-44. [ Links ]
Palacio, A. G., R. Noriega y P. Zamora. 2002. Caracterización físico-geográfico del paisaje conocido como bajos inundables. El caso del Área Natural Protegida Balamkín, Campeche. Investigaciones Geográficas 49:57-73. [ Links ]
Pausas, J. y M. Austin. 2001. Patters of plants species richness in relation to different environments: An appraisal. Journal of Vegetation Science 12:153-166. [ Links ]
Pennington, T. y J. Sarukhán. 2005. Árboles tropicales de México, Manual para la identificación de las principales especies. Tercera edición. Universidad Nacional Autónoma de México, Fondo de Cultura Económica, México, D. F. 523 p. [ Links ]
Puntieri, J. 2005. Variaciones intra-específicas en el crecimiento primario de Nothofagus dombeyi (Nothofagaceae). Boletín de la Sociedad Argentina de Botanica 40:181-192. [ Links ]
Rao, P. B., A. Kaur y A. Tewary. 2008. Drought resistance in seedlings of five important tree species in Tari region of Uttarakhand. Tropical Ecology 49:43-52. [ Links ]
Reuter, M., H. Tiessen, J. J. Jiménez-Osornio, J. Pohlan y P. L. G. Vlek. 2008. Establishment of Cordia dodecandra A.DC. with Bixa orellana L. on calcareous soils in Yucatán, Mexico. Advances in Agroforestry 4:195. [ Links ]
Rincon, E. y P. Huante. 1993. Growth responses of tropical deciduos tree seedlings to contrasting light conditions. Trees 7:202-207. [ Links ]
Rodríguez, G., J. Dorantes y E. Aquino. 2009. Ensayo de especies en la zona cálida del centro del estado de Veracruz, México. Foresta Veracruzana 1:19-24. [ Links ]
Romo, M. 2005. Efecto de la luz en el crecimiento de plántulas de Dipteryx micrantha Harms "Shihuahuaco" transplantadas a sotobosque, claros y plantaciones. Ecología Aplicada 4:1-8. [ Links ]
Sánchez, O., G. Islebe y M. Hernández. 2007. Flora arbórea y caracterización de gremios ecológicos en distintos estadios sucesionales de la selva mediana de Quintana Roo. Foresta Veracruzana 9:17-26. [ Links ]
Shankar, T. R., D. Mudappa y V. Kapoor. 2009. Restoring rainforest fragments: survival of mixed-native species seedlings under contrasting site conditions in the Western Ghats, India. Restoration Ecology 17:137-147. [ Links ]
Sherman, R., P. Martin y J. Timothy. 2005. Vegetation-environmental relationships in forest ecosystems of the Cordillera Central, Dominican Republic. Journal of the Torrey Botanical Society 132:293-310. [ Links ]
Souza, R. P. y I. F. Valio. 2003. Seedling growth of en Brazilian tropical tree species differing in succesional status. Revista Brasileña de Botánica 26:35-47. [ Links ]
Sun, G., G. Zuo, Liu S., M. Liu, S. McNulty y J. M. Vose. 2008. Watershed evapotranspiration increased due to changes in vegetation composition and structure under a subtropical climate. Journal of the American Water Resources Association 44:1164-1175. [ Links ]
Toledo, V., N. Barrera-Bassols, E. García-Frapolli y P. Alarcón-Chaires. 2008. Uso múltiple y biodiversidad entre los mayas yucatecos (México). INCI 33:345-352. <http://www.scielo.org.ve/scielo.php?script=sci_arttext&pid=S0378-18442008000500007&lng=es&nrm=iso>. ISSN 0378-1844; última consulta: 12.XII.2011. [ Links ]
Wacker, L., O. Baudois, S. Eichemberger-Glinz y B. Schmid. 2009. Effects of plant species richness on stand structure and productivity. Journal of Plant Ecology 2:95-106. [ Links ]
Wirth, R., B. Weber y R. J. Ryel. 2001. Spatial and temporal variability of canopy structure in a tropical moist forest. Acta Oecológica 22:1-10. [ Links ]
Wright, S. J. 2002. Plant diversity in tropical forest: a review of mechanisms of species coexistence. Oecologia 130:1-14. [ Links ]
Xu, F., G. Guo, W. H. Xu, Y. Wei y R. Wang. 2009. Leaf morphology correlates with water and light availability: what consequences for simple and compound leaves? Progress in Natural Science 19:1798-1798. [ Links ]
Zahawi, R. A. y C. K. Augspurger. 1999. Early plant succession in abandoned pastures in Ecuador. Biotropica 31:540-552. [ Links ]
Zamora, P., J. Flores y R. Ruenes. 2009. Flora útil y su manejo en el cono sur del estado de Yucatán, México. Polibotánica 28:227-250. [ Links ]
Zarco-Espinoza, V. M., J. L. Valdez-Hernández, G. Ángeles-Pérez y O. Castillo-Acosta. 2010. Estructura y diversidad de la vegetación arbórea del Parque Estatal Agua Blanca. Macuspuna, Tabasco. Universidad y Ciencia 26:1-17. [ Links ]
Zheng, D., J. Chen, B. Song, M. Xu, S. P. Phil y R. Jensen. 2000. Effects of silvicultural treatments on summer forest microclimate in southeastern Missouri Ozarks. Climate Research 15:45-59. [ Links ]

