










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de biodiversidad
versión On-line ISSN 2007-8706versión impresa ISSN 1870-3453
Rev. Mex. Biodiv. vol.84 no.4 México dic. 2013
https://doi.org/10.7550/rmb.36248
Ecología
Diversidad de las comunidades de escamados en la Reserva Natural del Iberá, Corrientes, Argentina
Squamate diversity in the Ibera Natural Reserve, Corrientes, Argentina
Eduardo G. Etchepare1,3*, María del Rosario Ingaramo1,3, Eduardo Porcel2 y Blanca B. Álvarez1
1 Laboratorio de Herpetología. Facultad de Ciencias Exactas y Naturales y Agrimensura. Universidad Nacional del Nordeste. Av. Libertad 5470, 3400 Corrientes, Argentina. *eduardoetchepare@hotmail.com
2 Departamento de Matemáticas, Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste. Av. Libertad 5470, 3400 Corrientes, Argentina.
3 Consejo Nacional de Investigaciones Científicas y Tecnológicas. Av. Rivadavia 1917 C1033AAJ Ciudad Autónoma de Buenos Aires, Argentina.
Recibido: 08 marzo 2013
Aceptado: 27 junio 2013
Resumen
La Reserva Natural del Iberá alberga uno de los humedales más importantes de Sudamérica y actualmente enfrenta distintos problemas de conservación debido a las actividades humanas. En el presente trabajo se compara la riqueza, abundancia y complementariedad —diversidad beta— de escamados en las comunidades vegetales más representativas de 2 localidades: paraje Galarza y colonia Pellegrini del margen oriental de la Reserva Natural del Iberá, Corrientes, Argentina. Entre 2007-2008 y mediante el método de muestreos por cuadrantes, se registraron 33 especies de escamados, lo que representa el 53% y 38% de las especies conocidas para la reserva y la provincia de Corrientes, respectivamente. La riqueza específica y la diversidad fueron mayores en colonia Pellegrini (S= 29 y 21.26 especies efectivas) que en paraje Galarza (S= 22 y 15.76 especies efectivas). La complementariedad de especies entre localidades fue del 45%. Los pastizales se destacaron por su alta riqueza, diversidad y número de especies exclusivas. Los esteros y bañados, así como los bosques fueron los que presentaron valores altos de complementariedad, reflejando la particularidad de las especies que lo componen. Los resultados demuestran la importancia que revisten los distintos ambientes nativos para la fauna de escamados y aportan información para el manejo y conservación de la Reserva Natural del Iberá.
Palabras clave: biodiversidad, conservación, reptiles, pastizales, esteros del Iberá, humedales.
Abstract
The Iberá Nature Reserve has one of the most important wetlands in South America. This system is currently facing various threats to its preservation as a result of human activities. In this work we compare the diversity and complementarity —beta diversity— of the squamates fauna in relation to plant communities representative of 2 localities: Paraje Galarza and Colonia Pellegrini on the eastern edge of the Nature Reserve of Iberá, Corrientes province, Argentina. Using the method of sampling by quadrants, we recorded 33 species of squamates between 2007 and 2008, representing 53% and 38% of known species for the reserve and Corrientes province, respectively. Species richness and species diversity were higher in Colonia Pellegrini (S= 29 and 21.26 effective species) than paraje Galarza (S= 22 and 15.76 effective species). The complementarity of species among sites was about 45%. The grasslands were notable for their high species richness, diversity and high number of unique species. The wetlands, marshes and forests, had the highest values of complementarity, reflecting the specificity of their species. Our results demonstrated the importance of the different native environments for conservation of squamates and provide useful information for the management and conservation of the Iberá Natural Reserve.
Key words: biodiversity, conservation, reptiles, grassland, estuaries of Iberá, wetlands.
Introducción
Con una superficie aproximada de 1 300 000 ha, la Reserva Natural del Iberá, constituye uno de los humedales más importantes de Sudamérica. Se ubica en el centro norte de la provincia de Corrientes, Argentina y ocupa un quinto de su superficie total. De ésta, solamente 24 550 ha que comprenden lagunas y esteros fueron declarados sitio Ramsar, por lo que sólo esas hectáreas cuentan con una protección efectiva (Burkart, 2006).
Los esteros del Iberá se caracterizan por tener una alta biodiversidad y por albergar numerosas especies bajo alguna categoría de amenaza (Álvarez et al., 2003; Abdala et al., 2012; Giraudo et al., 2012). Sin embargo, a pesar de poseer estos atributos, en las últimas décadas enfrenta distintos problemas de conservación como son la sustitución de ambientes nativos por monocultivos, práctica de ganadería tradicional con quemas no controladas, construcción de canales y mejoramiento de redes viales (Álvarez et al., 2003; Neiff, 2004; Neiff y Poi de Neiff, 2006), siendo las áreas adyacentes a los esteros las que manifiestan el mayor impacto en lo que concierne a la utilización de los recursos, y por este motivo es una zona donde urge mejorar los planes de manejo (Giraudo et al., 2006).
Los escamados —lagartijas, anfisbénidos y serpientes— son representantes típicos y conspicuos de este ecosistema y su preservación está en íntima relación con la conservación de la biodiversidad. Estos poseen una gran abundancia, diversidad de funciones, requerimientos ecofisiológiocos muy específicos y capacidad de dispersión baja, por lo que han sido considerados excelentes indicadores de la salud ambiental de los ecosistemas (Tuberville et al., 2005; Rey-Benayas et al., 2006; Carvajal-Cogollo y Urbina-Cardona, 2008). A pesar de ésto, sólo algunos aspectos de la composición de especies, la diversidad y patrones de distribución son conocidos para este grupo taxonómico en la región y se remiten a citas puntuales (Bergna et al., 1992; Montanelli y Álvarez, 1997; Tedesco, 1998; Etchepare e Ingaramo, 2008; Etchepare y Zaracho, 2009), a tratados generales de divulgación o listas de especies (Álvarez et al., 2002, 2003; Giraudo et al., 2006).
Como el conocimiento y la protección de la biodiversidad en sus distintos niveles se ha tornado uno de los objetivos centrales de la biología de la conservación, en los últimos años se han propuesto diferentes aproximaciones tendientes a medirla y cuantificarla (Gaston, 2000). Con base en ello, el objetivo del presente trabajo fue comparar la composición, diversidad y grado de reemplazo —diversidad alfa y beta— de los escamados en las comunidades vegetales más representativas de 2 localidades ubicadas en la margen oriental de la Reserva Natural del Iberá, provincia de Corrientes, Argentina. Dado que en la región existen distintos tipos de comunidades vegetales con condiciones ambientales contrastantes; incluyendo pastizales naturales, esteros, bosques y ambientes antropizados; se espera encontrar: 1) diferencias claras en la riqueza y diversidad de especies de escamados entre comunidades y entre localidades, y 2) una alta complementariedad y diferenciación en la composición de especies entre comunidades y entre localidades. De ser así, la conservación de los escamados en la región podría lograrse preservando la heterogeneidad ambiental, y no centrándose sólo en la protección de algún tipo de comunidad particular.
Materiales y métodos
El presente trabajo se llevó a cabo en la Reserva Natural del Iberá, donde confluyen 3 regiones fitogeográficas: Chaco Oriental, Paranaense y Espinal (Carnevali, 2003). Los muestreos se realizaron en 2 localidades: paraje Galarza (29°17' S, 58°43' O, departamento Santo Tomé) y colonia Pellegrini (28°33' S, 57°10' O, departamento San Martín), ambas ubicadas en la margen oriental de la reserva. Para su elección se tuvo en cuenta su ubicación fitogeográfica particular: la primera localidad en el distrito de los Campos de la provincia Paranaense y la segunda, por poseer características ecotonales con el Espinal. En cada una de estas localidades se identificaron las siguientes comunidades vegetales nativas, utilizando la descripción de Carnevali (2003):
Paraje Galarza: ') pastizal de Andropogon lateralis y Sorghastrum setosum (PA, pastizal de paja colorada y paja amarilla, en adelante): esta comunidad de 1 a 1.20 m de altura, se encuentra ampliamente difundida en toda el área. Es discontinua con otras cespitosas erectas reunidas en grupos, dejando espacios donde se desarrolla un tapiz graminoso; 2) malezal de A. lateralis (M, malezal, en adelante): comunidad que se establece en suelos con hidromorfismo acentuado, sujetos a anegamientos periódicos y prolongados por las lluvias estacionales, exhibiendo un microrelieve columnar y red de canalículos. Su fisonomía es mixta con un pastizal alto y una vegetación praderiforme hidrófila a palustre en la red de los pequeños canales; 3) bosques meso-higrófilos (IM, islotes de monte, en adelante): formaciones leñosas de 8 a 12 m de altura, dispersas como islas en los pastizales. De superficie pequeña, de 2 a 3 ha. Poseen una masa leñosa moderadamente alta y densa con escasa cantidad de lianas y epífitas, donde se destacan Handroanthus heptaphyllus (lapacho), Enterolobium contortisiliquum (timbó) y Nectandra angustifolia (laurel); 4) esteros y bañados (EB1): comunidad que reúne cuerpos de agua permanentes —los esteros— y otros de agua semipermanentes —los bañados— muy ligados ecológicamente. Los bañados se ubican en la periferia de los esteros y abarcan el área de expansión de sus aguas. Las especies más conspicuas del estero son Cyperus giganteus (pirí) y Schoenoplectus californicus (junco), mientras que las especies más abundantes del bañado son la flotantes libres y arraigadas Cephalanthus glabratus (sarandí) y Paspalum durifolium (paja azul).
Colonia Pellegrini: 5) sabanas leñosas (SL): pequeñas superficies de V a 1 ha con sabanas arbóreas de Prosopis affinis (ñandubay) y Acacia caven (aromito) que alternan con mosaico de prados de Elionurus muticus (espartillo), Axonopus sp. (pasto jesuita) Paspalum sp. (pasto horqueta) y pastizales de Andropogon lateralis (paja colorada) poco densos; 6) esteros y bañados (EB2): de características similares a las descritas para paraje Galarza; 7) espartillar de Elionurus muticus (E, espartillar, en adelante): pastizales de 0.60 a 1 m de altura. Especie que también se asocia a las sabanas leñosas descritas anteriormente y se disponen sobre suelos con distintos tipos de drenaje; 8) bosquecillos riparios (BR): bosques en galería de un centenar de metros de largo. Muestran un solo nivel arbóreo, de 6 a 8 m de altura, ocupado por 2 o 3 especies leñosas. Se destacan aquí especies hidrófilas tales como Erythrina crista-galli (ceibo), Inga uruguensis (ingá), Cecropia pachystachys (ambaí) y Croton urucurana (sangre de drago).
En cada localidad se incluyó un ambiente antropizado con el fin de compararlo con los nativos. En paraje Galarza se muestreó 9), un monocultivo de Eucalyptus sp. (ME, monocultivo, en adelante). Estas plantaciones exóticas han experimentado un crecimiento continuo en los últimos años en todo el sistema Iberá remplazando los ambientes nativos. En colonia Pellegrini se incluyó '0), un exobrador (EO), sitio ubicado 5 km al norte de dicha localidad y donde se encuentran restos de construcciones y carreteles utilizados para transportar cables de alta tensión. Esta obra, remplazó las sabanas leñosas que se encontraban originalmente en ese sitio.
Entre los años 2007 y 2008, se realizaron 8 viajes de campo —2 por estación— de 8 días de duración a cada una de las localidades seleccionadas. En cada comunidad vegetal descrita anteriormente, se aplicó el muestreo por cuadrantes, seleccionando 3 parcelas de 200 m2, que se examinaron exhaustivamente en busca de escamados. Esta técnica es las más adecuada para incluir la heterogeneidad del hábitat cuando se trabaja con varias especies que presentan diferentes preferencias de microhábitat (Heyer et al., 2001). Posteriormente, se elaboró una matriz de abundancia para las distintas comunidades vegetales de cada localidad y los taxones presentes en ellos.
Los primeros ejemplares de cada especie fueron recolectados y sacrificados según el método estándar establecido en la Guía para la Eutanasia Animal propuesto por la IACUC (The Institutional Animal Care and Use Committee). Luego fueron fijados con formol al 10% y conservados en alcohol al 70% para ser depositados en la Colección Herpetológica de la Universidad Nacional del Nordeste, Corrientes (UNNEC), como ejemplares de referencia. Los demás especímenes fueron identificados, marcados y liberados en el mismo ambiente donde fueron registrados.
La diversidad alfa se calculó mediante la riqueza (S) de especies por localidad y en cada comunidad vegetal estudiada. Esta es una de las estrategias más sencillas, sólidas y comúnmente usadas para evaluar la diversidad alfa (Myers et al., 2000; Moreno, 2001). Para determinar la existencia de diferencias significativas entre la riqueza de ambas localidades, se construyeron curvas de acumulación de especies, a partir de la riqueza específica y 40 muestras obtenidas de las 5 comunidades vegetales durante los 8 muestreos. Estas curvas "suavizadas", se realizaron mediante reordenamiento aleatorio repetido (999 repeticiones) de las muestras con intervalos de confianza del 95%.
Se determinó la diversidad por medio de la fórmula propuesta por Jost (2006), quien incorporó el término de diversidad verdadera:
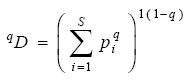
donde qD es la diversidad verdadera, pi es la abundancia relativa —abundancia proporcional— de la especie i, S es el número de especies y q es el orden de la diversidad y define la sensibilidad del índice a las abundancias relativas de las especies (Jost, 2006, 2007; Tuomisto, 2010a, 2010b; García-Morales et al., 2011; Moreno et al., 2011). El valor del parámetro q determina qué tanto influyen las especies comunes o las especies raras en la medida de la diversidad (Hill, 1973). Para este trabajo, la diversidad verdadera utilizada fue de orden 1 (1D), en la cual todas las especies son consideradas con un valor de diversidad proporcional a su abundancia en la comunidad (Jost, 2006, 2007; Tuomisto, 2010a, 2010b; Moreno et al., 2011). El resultado se expresa como el "número de especies efectivas" (Jost, 2006). Esta forma de expresar la diversidad de una comunidad, cumple con la propiedad de duplicación expuesta por Hill (1973). Esto posibilita unificar el análisis haciendo comparable los resultados de diversidad y también obtener una interpretación directa y más sencilla sobre cuál es la diferencia en la diversidad de 2 o más comunidades (Jost, 2007, 2010; Moreno et al., 2011). La diferencia en la diversidad verdadera entre 2 comunidades, también se expresó como el cociente, entre la de mayor diversidad respecto a la de menor diversidad. De esta forma, se puede ver cuánto más diversa es una comunidad comparada con otra. Otra forma de presentarla fue como el porcentaje de diversidad que representa la comunidad de menor respecto a la de mayor diversidad (Moreno et al., 2011).
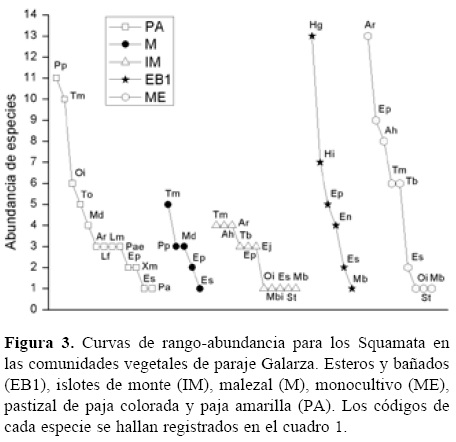
Para comparar la composición, abundancia y uniformidad de especies entre localidades y comunidades vegetales, se usaron curvas de rango-abundancia (Feinsinger, 2001). Para esto, se graficó la abundancia relativa de las especies contra el rango ocupado por cada especie desde la de mayor a la de menor abundancia (Urbina-Cardona et al., 2008).
Asumiendo que los inventarios pueden ser incompletos, es decir, que la muestra observada suele ser menor que la esperada (Moreno et al., 2011), se calculó el estimador de riqueza no-paramétrico Ace (Abundance-based coverage estimator, Chao y Lee, 1992), para medir la completitud del muestreo por localidad y comunidad vegetal (Colwell y Coddington, 1994). Este análisis se realizó con el software de acceso libre EstimateS versión v8.2.0 (http://purl.oclc.org/ estimates; Colwell, 2009).
La diversidad beta, entre localidades y comunidades vegetales, se calculó mediante el índice de complementariedad (CAB ). Este método mide el grado de recambio en la composición de especies entre pares de biotas (Colwell y Coddington, 1994). Cuando ambos sitios son idénticos en composición, el resultado es 0 y es 1 cuando las especies de ambos sitios son completamente distintas. El valor se puede expresar como el porcentaje de especies que son complementarias entre ambientes (Moreno, 2001).
Resultados
La fauna de escamados para toda el área de estudio estuvo representada por 33 especies, pertenecientes a 9 familias y 24 géneros, con un registro total de 386 individuos. Dentro de los saurios, las familias Teiidae y Anguidae presentaron 2 especies cada una, mientras que Gymnophtalmidae y Scincidae tuvieron sólo 1 especie. En el caso de las serpientes, la familia con más especies fue Colubridae con 23 y el resto de las familias —Boidae, Typhlopidae y Viperidae— estuvieron constituidas por 1 especie. Finalmente, sólo se registró 1 especie de anfisbénidos (Cuadro 1).
Según el estimador de riqueza calculado, la representatividad del muestreo en colonia Pellegrini fue de 96%; mientras que en paraje Galarza fue de 94%, mostrando un número de especies muy similar al encontrado en el campo.
La riqueza específica y abundancia fueron mayores en colonia Pellegrini (S= 29, 213 individuos) que en paraje Galarza (S= 22, 173 individuos). Los valores de riqueza fueron significativamente diferentes entre localidades, dado que los intervalos de confianza del 95% para las curvas de acumulación de especies no se superponen (Fig. 1). Lo mismo se observó en cuanto a la diversidad de especies, dado que el número efectivo de especies fue mayor en colonia Pellegrini (1D= 21.26) que en paraje Galarza (1D= 15.76), siendo 1.35 veces más diversa la primera localidad. Dicho de otra forma, en paraje Galarza hay una disminución del 26% de la diversidad de escamados.

Los patrones de distribución de la abundancia y el orden jerárquico de las especies entre ambas localidades, presentaron diferencias menores, observándose pendientes similares. Tupinambis merianae, Erythrolamprus poecilogyrus y Philodryas patagoniensis se ubicaron en las posiciones más altas de la curva. En paraje Galarza Atractus reticulatus se destacó entre las especies dominantes, mientras que para colonia Pellegrini se encontró dentro de las especies raras. Aunque la proporción de especies raras fue similar, ninguna se repite en este nivel de la curva (Fig. 2).

Según el estimador de riqueza calculado para cada comunidad vegetal en paraje Galarza, la representatividad del muestreo se consideró completo en el pastizal de paja colorada y paja amarilla (95%), malezal, y esteros y bañados (ambos con 93%), mientras que con una representatividad menor se encontraron islotes de monte y el monocultivo (ambos 76%).
La mayor riqueza específica en esta localidad se registró en el pastizal de paja colorada y paja amarilla (S= 13), seguido por los islotes de monte (S= 11), el monocultivo (S= 9), y los esteros y bañados (S= 6). Mientras que el malezal fue la comunidad vegetal donde se encontró el menor número de especies (S= 5; Cuadro 1). La mayor diversidad se registró en el pastizal de paja colorada y paja amarilla (1D= 10.3 especies efectivas), siendo 1.1 veces más diversa en especies que los islotes de monte (1D= 9.4 especies efectivas) y 1.6 veces más diversa que el monocultivo (1D= 6.5 especies efectivas). Por último, quedaron los esteros y bañados (1D= 4.6 especies efectivas) y el malezal (1D= 4.5 especies efectivas) como las unidades de paisajes menos diversas, mostrando una disminución en la diversidad del 55% y 56% en proporción al pastizal, respectivamente.
En paraje Galarza, 3 comunidades vegetales presentaron una marcada dominancia respecto a la abundancia de algunas de sus especies, Hydrodynastes gigas en esteros y bañados,A. reticulatusen el monocultivo y P.patagoniensis en el pastizal de paja colorada y paja amarilla. Tanto el malezal como los islotes de monte, presentaron pendientes poco pronunciadas, siendo T. merianae la especie que ocupó el nivel jerárquico más alto. Esta especie, a su vez, se encontró en la mayoría de las comunidades vegetales (Fig. 3).
De acuerdo con el estimador de riqueza calculado para cada comunidad vegetal en colonia Pellegrini, la representatividad del muestreo fue alta en el espartillar (90%), sabana leñosa (97%) y esteros y bañados (94%), mientras que fue baja en los bosquecillos riparios (76%) y en el exobrador (60%).
En esta localidad, las comunidades vegetales de mayor riqueza fueron el espartillar (S= 18) y el exobrador (S= 16). Luego, continuaron las sabanas leñosas (S= 11) y los bosquecillos riparios (S= 10). Por último, el menor número de especies se registró en los esteros y bañados, con sólo 9 especies (Cuadro 1). La mayor diversidad se observó en el espartillar (1D= 14 especies efectivas), siendo éste 29% más diverso que el exobrador (1D= 9.9 especies efectivas). Las sabanas leñosas, a pesar de presentar una riqueza específica menor que el ambiente antrópico, exhibió prácticamente la misma diversidad (1D= 9.8 especies efectivas), mientras que la menor diversidad se registró en los bosquecillos riparios (1D= 8.2 especies efectivas) y en los esteros y bañados (1D= 5.4 especies efectivas), mostrando una disminución en la diversidad, respecto al espartillar del 41% y 61%, respectivamente.
En colonia Pellegrini, las especies que ocuparon las posiciones jerárquicas más altas con dominancias marcadas fueron H. gigas en el estero y bañado y E. poecilogyrus en el exobrador. Asimismo, P. patagoniensis en el espartillar presentó una elevada abundancia, aunque la pendiente de la curva en esta comunidad no fue tan acentuada como la de las comunidades anteriores. Tanto la sabana leñosa como los bosquecillos riparios, exhibieron curvas con pendientes similares y poco pronunciadas (Fig. 4).

De acuerdo con el análisis de complementariedad, existió un 45% de especies complementarias entre ambas localidades.
El grado de disimilitud más alto en paraje Galarza se presentó entre esteros y bañados con el resto de las comunidades, con una complementariedad de especies, igual o mayor al 75% (Cuadro 2). Lo mismo se observó en los islotes de monte, con una complementariedad de especies igual o mayor al 74% con el resto de las comunidades vegetales, excepto con el monocultivo donde la complementariedad fue baja (18%). El pastizal de paja colorada y paja amarilla fue el sitio con mayor número de especies únicas.

En colonia Pellegrini, nuevamente esteros y bañados fue la comunidad vegetal más complementaria con el resto de las comunidades superando en todos los casos el 80%; además, fue el sitio con mayor número de especies únicas. Los bosquecillos riparios, mostraron un resultado similar, superando el 75% de especies complementarias, excepto con el exobrador, donde la complementariedad fue de 63%. El espartillar, junto con las sabanas y el exobrador, presentaron baja complementariedad, es decir, que comparten un gran número de especies (Cuadro 3).

Discusión
En este estudio se registró un total de 33 especies de escamados, lo que constituye el 53% de la fauna que se conoce para toda la Reserva Natural del Iberá (Álvarez et al., 2003; Giraudo et al., 2006; Etchepare e Ingaramo, 2008; Etchepare y Zaracho, 2009) y representa más de un tercio de la diversidad de escamados conocidos para la provincia de Corrientes, Argentina (Giraudo, 2001; Álvarez et al., 2002). Esto muestra, que a pesar de las modificaciones que sufre actualmente la reserva (Giraudo et al., 2006; Neiff y Poi de Neiff, 2006), aún sostiene condiciones que favorecen una gran diversidad de especies en este grupo taxonómico.
La alta riqueza y abundancia de especies observada en colonia Pellegrini podría deberse a que dicha localidad posee una gran proporción de terreno, de dominio fiscal y privado, dedicado a la conservación (Chebez, 2005; Burkart, 2006), a diferencia de paraje Galarza, donde la mayoría de los terrenos son de dominio privado y, por ende, los planes de manejo son de difícil aplicación, lo que ocasionaría una disminución en los parámetros mencionados. Rubio y Moreno (2010) obtuvieron resultados similares al comparar la riqueza y diversidad de especies de arañas con tejidos orbiculares en la misma zona de estudio. Sin embargo, Ingaramo et al. (2012) no encontraron diferencia significativa al comparar la riqueza de anuros entre colonia Pellegrini y paraje Galarza.
De acuerdo con los resultados obtenidos mediante el estimador de riqueza, calculado para las 2 localidades, se observó una buena aproximación al estado del conocimiento de la fauna local, debido a que los inventarios de especies estuvieron relativamente completos, superando al 90% de representatividad. Sin embargo, se puede esperar que la riqueza específica presente un incremento ligero en colonia Pellegrini si se continúan los muestreos.
El esfuerzo de muestreo por comunidad vegetal alcanzó una representatividad alta comparada con todo el estudio, excepto en los islotes de monte, bosquecillos riparios, exobrador y monocultivo. Por sus hábitos crípticos y su comportamiento evasivo, la probabilidad de detección de algunas de las especies de escamados, puede verse limitada, sobre todo, mediante la metodología de búsqueda por encuentros visuales (Vitt y Vangilder, 1983; Giraudo et al., 2004). Cabría esperar entonces, que la riqueza de especies sea mayor si aumenta el esfuerzo de muestreo, sobre todo, en los islotes de monte y bosquecillos riparios, donde la visibilidad es más limitada en comparación con el exobrador y el monocultivo.
Los pastizales de ambas localidades se destacaron por su alta riqueza, diversidad y por el gran número de especies registradas solamente en esta comunidad vegetal (Ophiodes aff. striatus, Lygophis flavifrenatus, Lygophis meridionalis, Philodryas agazissi, Xenodon merremi, Bothrops alternatus) demostrando la importancia que reviste para este grupo. Lo mismo se observó en estudios realizados con mamíferos y aves donde diversas especies tienen como ambientes exclusivos los pastizales de esta región (Stattersfield et al., 1998; Giraudo, 2001; Demaría et al., 2003; Giraudo et al., 2003; Bilenca y Miñarro, 2004; Giraudo y Di Giacomo, 2007; Di Giacomo et al., 2007). Por lo tanto, la sustitución de los mismos, produciría la extinción de la fauna que los habita y la supresión de los servicios ecosistémicos que ofrecen a la sociedad (Costanza et al., 1997; Balvanera et al., 2006; Baldi et al., 2008; León-Gamboa et al., 2010).
El monocultivo en paraje Galarza mostró una disminución de la diversidad del 37% y 31% comparado con el pastizal y los islotes de monte, respectivamente. El efecto de disminución de la riqueza y abundancia en monocultivos y en ambientes intervenidos se ha probado en varios organismos tales como, artrópodos (Jocqué, 1977; Bierregaard et al., 1992; León-Gamboa et al., 2010 ), anfibios (Lips et al., 2001), reptiles (Smith y Ballinger, 2001; Urbina-Cardona y Londoño-Murcia, 2003; Urbina-Cardona y Reynoso 2005; Calderón-Mandujano et al., 2008), aves (Woinarski, 1979; Di Giacomo y Krapovickas, 2001; Giraudo et al., 2003) y mamíferos (Dietz et al., 1975; Gorresen y Willig, 2004), siendo sugeridas como las principales causas, la fragmentación, simplificación y sustitución de los ambientes nativos.
A pesar de que el ambiente antrópico tomado en colonia Pellegrini presentó una diversidad alta, la abundancia de la mayoría de las especies fue baja, inclusive muchas de ellas presentaron sólo un registro. Esto podría indicar que utilizan este sitio como lugar de refugio alternativo, ya que existe una continuidad fisonómica entre el espartillar, las sabanas leñosas y el exobrador —sabana leñosa modificada—. Lo cual, explica también la baja complementariedad de especies observadas entre estas 3 comunidades.
Con respecto a la distribución de las abundancias y al orden jerárquico de las especies en ambas localidades, las diferencias fueron mínimas, esto demuestra que las comunidades de escamados se estructuran de forma similar. Sin embargo, se observaron diferencias al analizar las abundancias, el tipo de especies dominantes y la proporción de especies raras a nivel de las comunidades vegetales, lo que demuestra que en esta región las diferencias en las estructuras de las comunidades de escamados se presentan a esta escala. Los ambientes abiertos presentan condiciones fluctuantes, sobre todo, en las variaciones de temperatura, lo que beneficia a especies heliotérmicas generalistas (Medina-Rangel, 2011) tales como P. patagoniensis y T. merianae. Otras se asocian con ambientes que ofrecen condiciones de recursos más estables, como Amphisbaena heterozonata y A. reticulatus, que son especies adaptadas a vivir enterradas o debajo de troncos y hojarascas y Eunectes notaeus o H. gigas, serpientes adaptadas para vivir en diferentes cuerpos de agua como esteros y lagunas (Cei, 1993; Giraudo, 2001).
Al evaluar la diversidad beta entre localidades, se observó que la complementariedad de especies fue intermedia, mostrando una disimilitud del 45%. Ingaramo et al. (2012) encontraron valores similares para anuros (46%) en el porcentaje de especies compartidas, mientras que Rubio y Moreno (2010) observaron una complementariedad de especies relativamente alta en arañas (60%). Aunque ambas localidades pertenecen a regiones fitogeográficas distintas, las variaciones bioclimáticas y de las comunidades vegetales no inciden en la diferenciación de la fauna de anuros y escamados, como sí ocurre en arañas. Esto podría deberse a que la mayoría de los anuros y reptiles registrados en esta región son de distribución geográfica amplia (Cei, 1993; Giraudo, 2001; Álvarez et al., 2003; Ingaramo et al., 2012).
En ambas localidades, los esteros y bañados fue la comunidad vegetal más complementaria con el resto de los ambientes estudiados. A pesar de su riqueza y diversidad bajas, casi la totalidad de las especies fueron exclusivas para esta comunidad (E. notaeus, Helicops leopardinus, H. gigas, Thamnodynastes hypoconia y Thamnodynastes strigatus). Halffter y Moreno (2005) y Medina-Rangel (2011) mostraron que la complementariedad de especies entre hábitats depende de la disponibilidad de recursos en cada uno de ellos, por lo que las especies de escamados registradas en los esteros y bañados, realizarían un aprovechamiento diferencial de los recursos, conformando así un ensamble con características propias.
Se pudo observar en paraje Galarza que la complementariedad entre los islotes de monte y el monocultivo fue muy baja (18%). Frente a la fragmentación que actualmente sufre la primera comunidad vegetal debido a la práctica de tala selectiva (Carnevali 2003), las especies de escamados podrían estar utilizando el monocultivo de Eucalyotus sp. como refugio alternativo.
El reemplazo y modificación de los ambientes nativos en toda la Reserva Natural Iberá, se ha producido de forma acelerada en las últimas décadas (Neiff y Poi de Neiff, 2006), sobre todo, en los pastizales y esteros, los cuales se ven afectados por monocultivos de pino, eucalipto, arroz y soja, así como por la realización de canales para riego, quemas no controladas, tendido de redes de alta tensión, la caza con fines comerciales y el ecoturismo no planificado. Como consecuencia, se produce un cambio drástico del hábitat causando la fragmentación de los recursos y con esto la disminución de la biodiversidad. Por ello, es imprescindible intensificar los trabajos de inventarios biológicos en la zona debido a que la riqueza de vertebrados tetrápodos de los esteros del Iberá es sumamente elevada en el contexto de la Argentina (Giraudo et al., 2006), y es probable un incremento en la riqueza a medida que se intensifiquen este tipo de estudios. Este conocimiento, junto con la protección efectiva de los diferentes hábitats pueden dar respuestas concretas y eficaces para proteger y mantener la biodiversidad de la reserva.
Agradecimientos
A la Dirección de Recursos Naturales de la Provincia de Corrientes por los permisos de colecta. A las familias Falero y Barrios, y a Julieta Balparda por hospedarnos durante los viajes de campo. A los guardaparques de la Reserva Natural Iberá por facilitarnos el traslado dentro de la reserva. A los integrantes del Laboratorio de Herpetología, en especial a Roberto Aguirre, por su importante colaboración en los trabajos de campo. Se agradece a los 2 revisores anónimos por las valiosas sugerencias realizadas. A la Universidad Nacional del Nordeste por el financiamiento de este trabajo.
Literatura citada
Abdala, C. S., J. L. Acosta, J. C. Acosta, B. B. Alvarez, F. Arias, L. J. Avila, M. G. Blanco, M. Bonino, J. M. Boretto, G. Brancatelli, M. F. Breitman, M. R. Cabrera, S. Cairo, V. Corbalan, A. Hernando, N. R. Ibargüengoytía, F. Kakoliris, A. Laspiur, R. Montero, M. Morando, N. Pellegrini, C. H. F. Pérez, A. S. Quinteros, R. V. Semhan, M. E. Tedesco, L. Vega y S. M. Zalba. 2012. Categorization del estado de conservación de las lagartijas y anfisbenas de la República Argentina. Cuadernos de Herpetología 26:215-248. [ Links ]
Álvarez, B. B., R. H. Aguirre, J. A. Céspedez, A. B. Hernando y M. E. Tedesco. 2003. Herpetofauna del Iberá. In Fauna del Iberá, B. B. Alvarez (ed.). EUDENE, Corrientes. p. 99-178. [ Links ]
Álvarez, B. B., R. H Aguirre, J. A Céspedez, A. B. Hernando y M. E. Tedesco. 2002. Atlas de anfibios y reptiles de las provincias de Corrientes, Chaco y Formosa (Argentina). I. Anuros, Cecílidos, Saurios, Anfisbénidos y Serpientes. EUDENE, Corrientes. 156 p. [ Links ]
Baldi, G., M. D. Nosett y E. G. Jobbágy. 2008. El efecto de las plantaciones forestales sobre el funcionamiento de los ecosistemas sudamericanos. Ambiéncia - Revista do Setor de Ciéncias Agrárias e Ambientais 4:23-34. [ Links ]
Balvanera, P., A. B. Pfisterer, N. Buchmann, J. S. He, T. Nakashizuka, D. Raffaelli, B. Schmid. 2006. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecology Letters 9:1146-1156. [ Links ]
Bergna, S., L. Rey y B. Alvarez. 1992. Nuevas localidades para ofidios del nordeste argentino. Facultad de Ciencias Exactas y Naturales y Agrimensura 9:101-109. [ Links ]
Bierregaard Jr., R. O., T. E. Lovejoy, V. Kapos, A. A. Dos Santos y R. W. Hutchings. 1992. The biological dynamics of tropical rain forest fragments. A prospective comparison of fragments and continuos forest. BioScience 42:859-866. [ Links ]
Bilenca, D. y F. Miñarro. 2004. Identificación de áreas valiosas de pastizal (AVPs) en las Pampas y Campos de Argentina, Uruguay y sur de Brasil. Fundación Vida Silvestre Argentina. Buenos Aires. 351 p. [ Links ]
Burkart, R. 2006. Las áreas protegidas de Argentina. In La situación ambiental Argentina 2005, A. Brown, U. Martínez-Ortiz, M. Acerbi y J. Corcuera (eds.). Fundación Vida Silvestre Argentina. Buenos Aires. p. 399-404. [ Links ]
Calderón-Mandujano, R. R., C. Galindo-Leal, y J. R. Cedeño-Vázquez. 2008. Utilización de hábitat por reptiles en estados sucesionales de selvas tropicales de Campeche, México. Acta Zoológica Mexicana 24:95-114. [ Links ]
Carnevali, R. 2003. El Iberá y su entorno fitogeográfíco. EUDENE, Corrientes. 112 p. [ Links ]
Carvajal-Cogollo, J. E. y J. N. Urbina-Cardona. 2008. Patrones de diversidad y composición de reptiles en fragmentos de bosque seco tropical en Córdoba, Colombia. Tropical Conservation Science 1:397-416. [ Links ]
Cei, J. M. 1993. Reptiles del noroeste, nordeste y este de la Argentina. Herpetofauna de las selvas subtropicales, Puna y Pampas. Monografie XIV, Museo Regionale di Scienze Naturali - Torino. 949 p. [ Links ]
Chao, A. y S. M. Lee. 1992. Estimating the number of classes via sample coverage. Journal of the American Statistical Association 87:210-217. [ Links ]
Chebez, J. C. 2005. Guía de las reservas naturales de la Argentina: nordeste. Albatros, Buenos Aires. 288 p. [ Links ]
Colwell, R. K. y J. A. Coddington. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society of London Series B 345:101-118. [ Links ]
Colwell, R. K. 2009. EstimateS: statistical estimation of species richness and shared species from samples, version 8.2.0. User's guide and application. http://purl.oclc.org/estimates; última consulta: 6.XII.2012. [ Links ]
Costanza, R., R. d'Arge, R. de Groot, S. Farber, M. Grasso, B. Hannon, K. Limburg, S. Naeem, R. V. O'Neill, J. Paruelo, R. G. Raskin, P. Sutton y M. Van den Belt. 1997. The value of the world's ecosystem services and natural capital. Nature 387:253-260. [ Links ]
Demaría, M. R., W. J. Mcshea, K. Koy y N. O. Maceira. 2003. Pampas deer conservation with respect to habitat loss and protected area considerations in San Luis, Argentina. Biological Conservation 115:121-130. [ Links ]
Di Giacomo, A. S. y S. Krapovickas. 2001. Afforestation threatens Argentina's grasslands. World Birdwatch 23:24-25. [ Links ]
Di Giacomo, A. S., M. V. De Francesco y E. G. Coconier. 2007. Areas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: Temas de Naturaleza y Conservación 5. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires. 163 p. [ Links ]
Dietz, J. M., E. A. Couto, C. A. Alfenas, A. Faccini y G. F. Da Silva. 1975. Efeitos de duas plantações de florestas homogeneas sobre populações de mamiferos pequenos. Brasil Florestal 23:54-57. [ Links ]
Etchepare, E. G. y M. R. Ingaramo. 2008. Pseudablabes agassizii (Jan, 1863) (Serpentes: Colubridae). Primer registro para la provincia de Corrientes (República Argentina). Cuadernos de Herpetología 22:51. [ Links ]
Etchepare, E. G. y V. H. Zaracho. 2009. Serpentes, Colubridae, Taeniophallus poecilopogon: Rediscovery in Corrientes, Argentina, and natural history. Check List 5:770-772. [ Links ]
Feinsinger, P. 2001. Designing field studies for diversity conservation. Island, Washington D. C. 212 p. [ Links ]
García-Morales, R., C. E. Moreno y J. Bello-Gutiérrez. 2011. Renovando las medidas para evaluar la diversidad en comunidades ecológicas: el número de especies efectivas de murciélagos en el sureste de Tabasco. Therya 2:205-215. [ Links ]
Gaston, K. J. 2000. Global patterns in biodiversity. Nature 405:220-227. [ Links ]
Giraudo, A. R. 2001. Serpientes de la selva Paranaense y del Chaco Húmedo. Taxonomía, biogeografía y conservación. LOLA, Buenos Aires. 381 p. [ Links ]
Giraudo, R. A., A. S. Di Giacomo, M. Ordano, E. Krauczuk, M. Chatelleneaz y C. Saibene. 2003. Aves amenazadas de los esteros del Iberá: un refugio que se desvanece. In Fauna del Iberá, B. B. Alvarez (ed.). EUDENE, Corrientes. p. 273-303. [ Links ]
Giraudo, A. R., V. Arzamendia y S. López. 2004. Ofidios del litoral fluvial de Argentina (Reptilia: Serpentes): biodiversidad y síntesis sobre el estado actual de conocimiento. INSUGEO, Miscelánea 12:323-330. [ Links ]
Giraudo, A. R., A. Bortoluzzi y V. Arzamendia. 2006. Vertebrados tetrápodos de la reserva y sitio Ramsar "Esteros del Iberá" (Corrientes, Argentina): análisis de su composición y nuevos registros para especies amenazadas. Natura Neotropicalis 37:1-20. [ Links ]
Giraudo, A. R. y A. S. Di Giacomo. 2007. Galarza. In Areas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad, A. S. Di Giacomo, M. V. De Francesco y E. G. Coconier (eds.). Temas de Naturaleza y Conservación 5. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires. p. 153-154. [ Links ]
Giraudo, A. R., V. Arzamendia, G. P. Bellini, C. A. Bessa, C. C. Calamante, G. Cardozo, M. Chiaraviglio, M. B. Costanzo, E. G. Etchepare, V. Di Cola, D. O. Di Pietro, S. Kretzschmar, S. Palomas, S. J. Nenda, P. C. Rivera, M. E. Rodríguez, G. J. Scrocchi y J. D. Williams. 2012. Categorización del estado de conservación de las serpientes de la República Argentina. Cuadernos de Herpetología 26:303-326. [ Links ]
Gorresen, P. y M. R. Willig. 2004. Landscape responses of bats to habitat fragmentation in Atlantic forest of Paraguay. Journal of Mammalogy 85:688-697. [ Links ]
Halffter, G. y C. E. Moreno. 2005. Significado biológico de las diversidades alfa, beta y gamma. In Sobre diversidad biológica: el significado de las diversidades alfa, beta y gamma, G. Halffter, J. Soberón, P. Koleff y A. Meliá (eds.). Monografías tercer milenio, Sociedad Entomológica Aragonesa, Zaragoza. p. 5-18. [ Links ]
Heyer, W. R., M. A. Donnelly, R. W. Mc Diarmid, L. C. Hayek y M. S. Foster. 2001. Medición y monitoreo de la diversidad biológica. Métodos estandarizados para anfibios. Smithsonian Institution. Universitaria de la Patagonia, Chubut. 350 p. [ Links ]
Hill, M. O. 1973. Diversity and evenness: a unifying notation and its consequences. Ecology 54:427-432. [ Links ]
Ingaramo, M. R., E. G. Etchepare, B. B. Alvarez y E. Porcel. 2012. Riqueza y composición de la fauna de anuros en la región oriental de la Reserva Natural Provincial Esteros del Iberá, Corrientes, Argentina. Revista de Biología Tropical 60:759-769. [ Links ]
Jocqué, C. A.1977. Malawi. A terrestrial baselines study of the Viphya Pulpmill Project Area. FAO, Internal Report, Rome. 36 p. [ Links ]
Jost, L. 2006. Entropy and diversity. Oikos 113:363-375. [ Links ]
Jost, L. 2007. Partitioning diversity into independent alpha and beta components. Ecology 88:2427-2439. [ Links ] Jost, L. 2010. The relation between evenness and diversity. Diversity 2:207-232. [ Links ]
León-Gamboa, A. L, C. Ramos y M. R. García. 2010. Efecto de plantaciones de pino en la artropofauna del suelo de un bosque Altoandino. Revista de Biología Tropical 58:1031-1048. [ Links ]
Lips, K. R., J. K. Reaser, B. E. Young y R. Ibáñez. 2001. Monitoreo de anfibios en América Latina: Manual de protocolos. Society for study of amphibians and reptiles. Herpetological Circulars 30. Ithaca. 114 p. [ Links ]
Medina-Rangel, G. F. 2011. Diversidad alfa y beta de la comunidad de reptiles en el complejo cenagoso de Zapatosa, Colombia. Revista de Biología Tropical 59:935-968. [ Links ]
Montanelli, S y B. B. Alvarez. 1997. Tantilla melanocephala. Geographic Distribution. Herpetolgical Reviow 29:179. [ Links ]
Moreno, C. E. 2001. Métodos para medir la biodiversidad. M&T-Manuales & Tesis SEA, Zaragoza. 84 p. [ Links ]
Moreno, C. E., F. Barragán, E. Pineda y N. P. Pavón. 2011. Reanalizando la diversidad alfa: alternativas para interpretar y comparar información sobre comunidades ecológicas. Revista Mexicana de Biodiversidad 82:1249-1261. [ Links ]
Myers, N., R. A. Mittermeier, C. G. Mittermeier, G. A. B. da Fonseca y J. Kent. 2000. Biodiversity hotspots for conservation priorities. Nature 403:853-858. [ Links ]
Neiff, J. J. 2004. El Iberá. ¿en peligro? Fundación Vida Silvestre de Argentina, Buenos Aires. 100 p. [ Links ]
Neiff. J. J. y A. Poi de Neiff. 2006. Situación ambiental en la ecorregión Iberá. In La Situación Ambiental Argentina 2005, A. Brown, U. Martínez-Ortiz, M. Acerbi y J. Corcuera (eds.). Fundación Vida Silvestre Argentina. Buenos Aires. p. 177-184. [ Links ]
Rey-Benayas, J. M., E. De La Montaña, J. Belliure y X. R. Eekhout. 2006. Identifying areas of high herpetofauna diversity that are threatened by planned infrastructure projects in Spain. Journal of Environmental Management 79:279-289. [ Links ]
Rubio, G. D y C. E. Moreno. 2010. Orb-weaving spider diversity in the Iberá Marshlands, Argentina. Neotropical Entomology 39:496-505. [ Links ]
Smith, G. R. y R. E. Ballinger. 2001. The ecological consequences of habitat and microhabitat use in lizards: a review. Contemporary Herpetology 3:1-32. [ Links ]
Stattersfield, A. J., M. J. Crosby, A. J. Long y D. C. Wege. 1998. Endemic bird areas of the world: priorities for biodiversity conservation. Bird Life International, Cambridge. 815 p. [ Links ]
Tedesco, M. E. 1998. Una nueva especie de Pantodactylus (Squamata, Gymnophthalmidae) de la provincia de Corrientes, República Argentina. FaCENA 14:53-62. [ Links ]
Tuberville, T. D., J. D. Willson, M. E. Dorcas y J. W. Gibbons. 2005. Herpetofaunal species richness of southeastern national parks. Southeastern Naturalist 4:537-569. [ Links ]
Tuomisto, H. 2010a. A consistent terminology for quantifying species diversity? Yes, it does exist. Oecologia 164:853-860. [ Links ]
Tuomisto, H. 2010b. A diversity of beta diversities: straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity. Ecography 33:2-22. [ Links ]
Urbina-Cardona, J. N. y M. C. Londoño-Murcia. 2003. Distribución de la comunidad de herpetofauna asociada a cuatro áreas con diferente grado de perturbación en la isla Gorgona, Pacífico Colombiano. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 27:105-113. [ Links ]
Urbina-Cardona, J. N. y V. H. Reynoso. 2005. Recambio de anfibios y reptiles en el gradiente potrero-borde-interior en Los Tuxtlas, Veracruz, México. In Sobre diversidad biológica: el significado de las diversidades alfa, beta y gamma, G. Halffter, J. Soberón, P. Koleff y A. Meliá (eds.). Monografías tercer milenio, Sociedad Entomológica Aragonesa, Zaragoza. p. 191-207. [ Links ]
Urbina-Cardona, J. N., M. C. Londoño-Murcia y D. G. García-Ávila. 2008. Dinámica espacio-temporal en la diversidad de especies de serpientes en cuatro hábitats con diferente grado de alteración antropogénica en el Parque Nacional Natural Isla Gorgona, Pacífico Colombiano. Caldasia 30:479-493. [ Links ]
Vitt, L. J. y L. D. Vangilder. 1983 Ecology of a snake community in Northeastern Brazil. Amphibia-Reptilia 4:273-296. [ Links ]
Woinarski, J. K. Z. 1979. Birds of Eucalyptus plantation and adjacent natural foresto. Australian Forestry 42:243-247. [ Links ]

