










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de biodiversidad
versión On-line ISSN 2007-8706versión impresa ISSN 1870-3453
Rev. Mex. Biodiv. vol.84 no.3 México sep. 2013
https://doi.org/10.7550/rmb.33819
Taxonomía y sistemática
An enigmatic and large-sized specimen of Panochthus (Glyptodontidae, "Panochthini") from the Ensenadan (Early-Middle Pleistocene) of the Pampean region, Argentina
Un enigmático gran ejemplar de Panochthus (Glyptodontidae, "Panochthini") del Ensenadense (Pleistoceno Temprano-Medio) de la región pampeana de Argentina
Martín Zamorano1*, Gustavo Juan Scillato-Yané1 and Alfredo Eduardo Zurita2
1 Departamento Científico de Paleontología de Vertebrados, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. Paseo del Bosque s/n, 1900 La Plata. Argentina. *marzamorano@fcnym.unlp.edu.ar
2 Centro de Ecología Aplicada del Litoral-Consejo Nacional de Investigaciones Científicas y Técnicas and Universidad Nacional del Nordeste, Ruta 5, km 2.5 CC, 128 (3400), Corrientes, Argentina.
Recibido: 05 octubre 2012
Aceptado: 22 marzo 2013
Abstract
Panochthus Burmeister (Mammalia, Xenarthra, Glyptodontidae) is one of the best characterized Pleistocene genera. It is possible to recognize 2 Ensenadan (Early-Middle Pleistocene) species from the Pampean region of Argentina and Bolivia, Panochthus intermedius Lydekker and P. subintermedius Castellanos, whereas 2 species are considered as valid for the age Bonaerian-Lujanian (Middle Pleistocene-Late Pleistocene), namely P. tuberculatus Owen and P. frenzelianus Ameghino. This paper provides the first record of a skull belonging to Panochthus from Ensenadan deposits in the Pampean Region of Argentina, being the second record for the genus in this lapse. The synapomorphies that support this taxonomic interpretation are: a frontal-ventrally inclined nasal zone forming a 45° angle between the palate plane and the plane of the upper rostral zone; a cranial length/height ratio that less than 154 of the height, without considering the descending processes of the zygomatic arch; external nasal openings with a fronto-ventral orientation. A comparative study with P. intermedius and other species from the Bonaerian-Lujanian refutes its placement among any of these; on the other hand, no cranial materials are known for the other species from the Ensenadan (P. subintermedius). Considering that the former is a large species, and that this skull corresponds undoubtedly to a species of Panochthus of large dimensions, the most parsimonious taxonomic hypothesis is to tentatively assign the material MLP 84-IX-2-11 to Panochthus cf. subintermedius. Future findings of more complete materials would provide support or refute this hypothesis.
Key words: taxonomy, South America, Pleistocene, Glyptodontidae, diversity.
Resumen
Panochthus Burmeister (Mammalia, Xenarthra, Glyptodontidae) es uno de los gliptodontes mejor caracterizados para el Pleistoceno. Se han reconocido 2 especies para el Ensenadense (Pleistoceno temprano-medio), P. intermedius Lydekker y P. subintermedius Castellanos, mientras que para los pisos Bonaerense-Lujanense (Pleistoceno medio-Pleistoceno tardío), se hallan P. tuberculatus Owen y P. frenzelianus Ameghino. En esta contribución damos a conocer el primer registro craneano de Panochthus para el Ensenadense de la región Pampeana y el segundo conocido para el género durante este lapso. Las sinapomorfías que sustentan su asignación son: región nasal con inclinación ventral, formando un ángulo de 45° entre el plano palatal y el rostral; índice craneal diámetro antero-posterior/diámetro transverso menor a 14 la altura craneana (sin considerar los procesos descendentes del maxilar); aberturas nasales inclinadas fronto-ventralmente. Un estudio comparado con P. intermedius y las otras especies de Panochthus del Bonaerense-Lujanense indica que este material no corresponde a ninguna de ellas; por otra parte, no se conoce aún el cráneo de P. subintermedius, la otra especie del Ensenadense. Considerando que esta última especie es de gran tamaño y que el cráneo aquí presentado corresponde también a un ejemplar de gran tamaño, la hipótesis taxonómica más parsimoniosa consiste en asignar el material MLP 84-IX-2-11 a Panochthus cf. subintermedius. Nuevos hallazgos permitirán confirmar o refutar esta postura.
Palabras clave: taxonomía, América del Sur, Pleistoceno, Glyptodontidae, diversidad.
Introduction
The record for the Glyptodontidae "Panochthini" extends from the late Miocene to the late Pleistocene of South America, and includes the genera Nopachtus Ameghino, 1888, Propanochthus Castellanos, 1925, and Panochthus Burmeister, 1866 (see Castellanos, 1942; Scillato-Yané et al., 1995; Cione et al., 1999; Zamorano, 2012; Zamorano and Brandoni, 2012). However, recent cladistic analyses suggest that this tribe might not be a natural group, because Tertiary taxa seem to be more related to some species of Hoplophorinae "Plohophorini" (see Zamorano, 2012; Zamorano and Brandoni, 2012).
Panochthus is perhaps one of the best-characterized Pleistocene genera (see Burmeister, 1874), with records ranging from Santa Cruz Province in Argentina (Tauber and Palacios, 2007) to northeastern Brazil (Porpino and Bergqvist, 2002). This taxon was described by Burmeister (1866), using as type a species originally recognized by Owen (1845), Glyptodon tuberculatus.
The monophyly of this genus has been recently tested, and the results suggest that it is supported by several synapomorphies. These include: a ventrally inclined nasal region forming a 45° angle between the palate plane and the plane of the upper rostral zone; the cranial length/ height ratio being less than 1½ of the height, without considering the descending processes of the zygomatic arch; the external nasal openings presenting a fronto-ventral orientation; and the central figures of the osteoderms in the dorsal region of the caudal tube being mostly rugged (Zamorano, 2012).
The only valid species of Panochthus recognized for the Ensenadan Age/Stage (Early-Middle Pleistocene) of the Pampean Region are P. intermedius Lydekker, 1895 and P. subintermedius Castellanos, 1937 (Zamorano, 2005; Cruz et al., 2011; Zamorano, 2012). P. intermedius was recognized on the basis of an almost complete dorsal carapace, found in Buenos Aires Harbor. According to Ameghino (1895, 1909) the material came from "basal Ensenadan" levels (sensu Ameghino, 1909). On the other hand, Cruz et al. (2011) revalidated the species P. subintermedius. According to Castellanos (1942) the holotype came from "top Ensenadan" levels (sensu Ameghino, 1909) from Buenos Aires city, although the excavation depth was not stated.
In this scenario, a partial skull was exhumed in 1998 from a quarry located in Hernández (surroundings of La Plata, Buenos Aires Province, Argentina; Fig. 1). It was catalogued as MLP 84-IX-2-11 and referred to the genus Panochthus, given that the specimen shares all the synapomorphies of the genus.
Previous works (Scillato-Yané and Carlini, 1998; Tonni et al., 1999; Zamorano and Scillato-Yané, 2006) assigned this specimen to Panochthus cf. intermedius. However, Zurita et al. (2011) presented new material from the Early Pleistocene of Bolivia, which was referred to P. intermedius. In this sense, the noticeable morphological differences between both skulls preclude the assignment of the material MLP 84-IX-2-11 to P. cf. intermedius. In turn, there is no known skull for P. subintermedius. This situation poses a number of questions about the specific identity of the material MLP 84-IX-2-11.
This work presents a detailed anatomic description of the skull MLP 84-IX-2-11, and compares this material with other species of Panochthus and other glyptodonts in order to discuss some taxonomic aspects of this Ensenadan Panochthini.
Materials and methods
The anatomical terminology follows Gillette and Ray (1981), Zurita (2007), and Zamorano (2012). Comparative materials correspond to skulls of known species of Panochthus (P. intermedius, P. tuberculatus, and P. frenzelianus), other Glyptodontidae (e.g., Glyptodon reticulatus Owen, 1845), and Pampatheriidae (Appendix). All measurements were taken with a digital caliper rounding up to 0.5 mm. The following comparative measurements are listed in table 1: maximum length (ML), from the anterior end of the nasal apertures to the upper margin of foramen magnum, measured on the sagittal plane; maximum bi-zygomatic width (BW); maximum transverse diameter of the orbital notch (MTDO); maximum dorso-ventral diameter of the orbital notch (MDVO); maximum transverse diameter of temporal fenestra (MTDT); maximum anterior-posterior diameter of temporal fenestra (MAPDT); dorso-ventral diameter of the zygomatic arch below the orbit (DZABO); dorso-ventral diameter of the zygomatic arch below the temporal fenestra (DZABTF); transverse diameter of the palate (TDP), following a transverse axis between the molariforms M7 and M8; maximum dorso-ventral diameter of choanae apertures (MDVCA); maximum transverse diameter of choanae apertures (MTCA); minimum bicondylar transverse diameter (mBTD), i.e., the minimal distances between the occipital condyles; total length of dental series (LDS) from the anterior end of M1 alveolus to the posterior border of M8 alveolus.
Institutional abbreviations. AMNH: American Museum of Natural History (New York, New York, USA); MCA: Museo Carlos Ameghino, Mercedes, Buenos Aires; MERO: Museu do Estado de Rondônia, Porto Velho, Rondônia, Brazil. MLP: División Paleontología Vertebrados, Facultad de Ciencias Naturales y Museo de Ciencias Naturales (La Plata, Buenos Aires, Argentina); MHGN: Départament de Géologie et de Paléontologie, Muséum d'Histoire Naturelle (Genève, Schweiz); MHNC: Museo de Historia Natural de Cochabamba "Alcide d'Orbigny" (Cochabamba, Bolivia).
Geographic and stratigraphic setting. The quarry at Hernández is located 5.3 km NW of La Plata (34°54'35" S, 58°00'15" W), Buenos Aires Province (Fig. 1). Excavations at the quarry reached 13.9 m deep, exposing deposits of the Ensenada, Buenos Aires, and Luján formations (Pleistocene-Holocene). The specimen MLP 84-IX-2-11 was recovered from the Ensenada Formation, being in stratigraphic association with Pseudoseisura sp., Neosclerocalyptus Paula Couto, 1954 sp., and Mesotherium cristatum Serres, 1867 (Tonni et al., 1999). Bed sediments were clay-sandy muds, of a brown-yellowish color (Fig. 2). Previous magnetostratigraphic studies dated these sediments at 0.78 My BP (Tonni et al., 1999).

Description
Magnorder Xenarthra Cope, 1889
Order Cingulata Illiger, 1811
Suborder Glyptodontia Ameghino, 1889
Superfamily Glyptodontoidea Gray, 1869
Family Glyptodontidae Gray, 1869
Panochthus Burmeister, 1866
Panochthus cf. subintermedius Castellanos, 1937 Figs. 3, 4
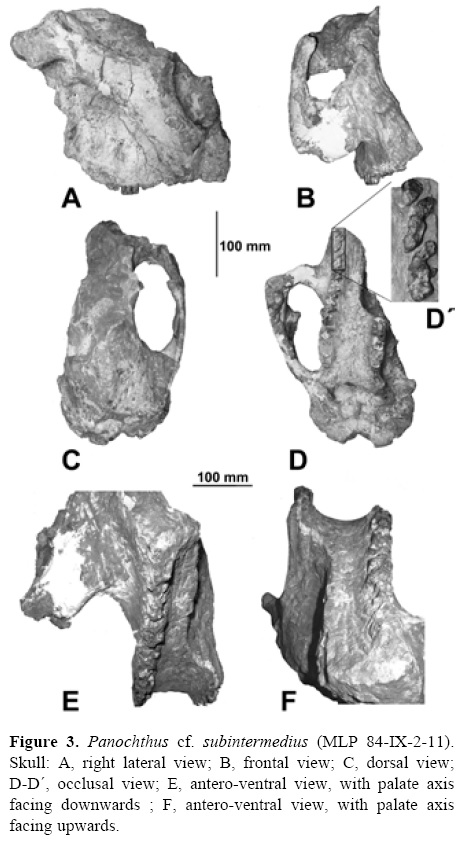
Stratigraphic and geographic provenance: upper section of Ensenada Formation (Early-Middle Pleistocene), quarry at Hernández, surroundings of La Plata, Buenos Aires Province.
Referred material: MLP 84-IX-2-11, incomplete skull, with its right half best preserved. Anterior end of rostral region and frontal sinuses missing; zygomatic arch partially complete with broken descending apophysis; postorbital apophysis missing. Right upper dental serie complete, albeit with molariforms not well preserved. Left half with only 3 posterior molariforms (M6-M8), and partly fragmented.
Remarks. In anterior view, the nasofrontal region is ventrally inclined forming a >45° angle with the palate plane (Figs. 3A, B), as observed in all species of the genus Panochthus. Even when the nasal and frontal regions are partially preserved in the skull MLP 84-IX-2-11, it can be inferred that the degree of pneumatization does not reach the degree observed in P. tuberculatus, P. frenzelianus, and P. intermedius.
In lateral view (Fig. 3A), the orbital notch is dorso-ventrally elongated, as in P. frenzelianus and P. intermedius, and differs from that of P. tuberculatus, which has a subcircular contour. The orbital fossa is larger than in other species of the genus (Table 1). The postorbital process, not united to the zygomatic arch, is placed between the orbital fossa and the temporal fossa; therefore both fossae are connected, as in P. frenzelianus, P. intermedius, and all species of the genus Glyptodon. On the other hand, P. tuberculatus, Neosclerocalyptus, and Doedicurus Burmeister, 1874 present a complete postorbital process, which obliterates the connection between both fossae in external view.
The contour of the orbital fossa is broken in the lacrimal bone area; however, it can be inferred that the lacrimal tubercle protrudes, as in P. tuberculatus, and unlike in P. frenzelianus. The descending apophysis of the maxilla is not preserved, thus its length and morphology cannot be observed. The zygomatic arch is more robust than that of P. intermedius, P. tuberculatus, and P. frenzelianus, whereas its dorso-ventral diameter diminishes in an antero-posterior direction, as in P. frenzelianus. The dorso-ventral diameter of the arch is more constant than in P. tuberculatus, a character shared with P. frenzelianus. Posteriorly to the orbital fossa, the zygomatic arch does not protrude laterally, but rather reaches gradually the area of its temporal insertion, as is the case in P. frenzelianus, and unlike in P. tuberculatus. The zygomatic arch of MLP 84-IX-2-11 describes a concave curve in the posterior-dorsal direction, running from below the orbit to its insertion in the temporal zone, as is the case in P. frenzelianus and P. intermedius; conversely, in P. tuberculatus the zygomatic arch runs straight from behind the orbit to immediately before its insertion.
The alveolar edge of the maxillae is irregular, being elevated in the middle. This morphology differs from the other species of this genus, in which the alveolar edge of the maxillae is straight.
A sagittal crest is observed in dorsal view (Fig. 3A), despite the poor preservation of parietal and frontal bones, a character that is absent in P. tuberculatus. Besides, the lambdoid or parieto-occipital crest is well developed, more than in P. intermedius and much more than in P. tuberculatus. The temporal fossa is larger than that of P. tuberculatus and P. intermedius. The temporal muscle, which acts together with the masseter and the pterigoid to elevate the mandible, inserts into this fossa (Sisson and Grossman, 1959: 245).
In posterior view, the foramen magnum has an elliptic section, with the major axis being transverse. The occipital condyles are large, with a sub-elliptic section and the major axis is transverse; conversely, in P. tuberculatus and P. intermedius, the occipital condyles are proportionally much smaller, and have a sub-quadrangular section. The contour of the aperture of the choanae is rectangular, with the transverse diameter being slightly larger than the dorso-ventral, a character that differs from P. tuberculatus and P. intermedius, in which the transverse diameter is the smaller one.
In occlusal view (Figs. 3D-F), the zygomatic arch is more markedly separated from the neurocranium than in dorsal view (Fig. 3A), similar to that observed in P. intermedius and P. tuberculatus, although in these taxa this character is less evident. The palate is concave (Fig. 3E, F), unlike in P. tuberculatus, in which it is flat. This structure has a particular morphology, gradually descending until it joins the alveoli forming an obtuse angle. In contrast, this union is more abrupt, i.e., almost at a right angle, in P. tuberculatus; whereas P. intermedius presents an intermediate morphology between the above-mentioned species.
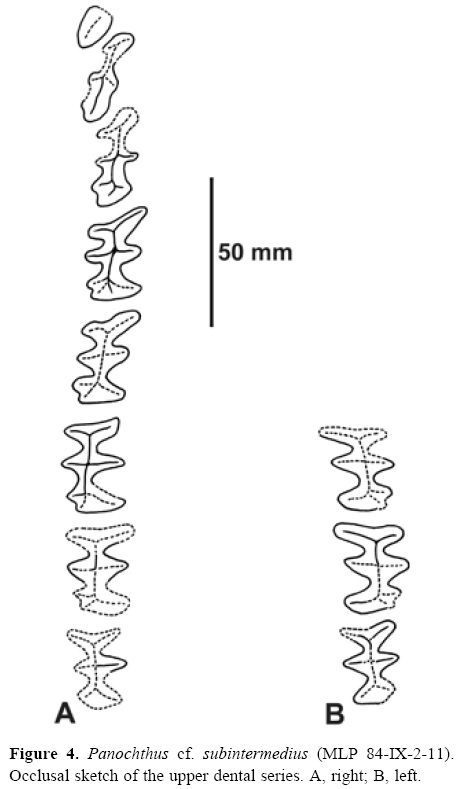
Dentition. Three left molariforms (M6-M8) are preserved (Fig. 4B), but the anterior lobe of M6 is missing. The right dental series is complete (Fig. 4A), albeit the M7 and M8 are poorly preserved (i.e., the occlusal surface is not visible). The complete dental serie is proportionally longer than that of P. tuberculatus (Table 1). The imbrication of M1-M3 (Figs. 3D', 4A) is a singular character, because this is not present in any other Glyptodontidae "Hoplophorinae", whereas it is shared with several Cingulata Pampatheridae (e.g., all species of Holmesina Simpson; see Góis et al., 2012). The M1 presents a sub-elliptic section, making it very different from the M1 of P. tuberculatus and P. intermedius, which is trilobed with isodiametric lobes. M2 is narrow and elongated, more than M4. M3 has a particular morphology, being more elongated than all other molariforms; it is narrow too, but not as much as M2. It presents 3 transverse lobes, with an additional, posterior lobe. The M4-M8 are trilobated and similar to those of P. tuberculatus.
Discussion
In previous works (e.g., Scillato-Yané and Carlini, 1998; Tonni et al., 1999; Zamorano and Scillato-Yané, 2006), the specimen MLP 84-IX-2-11 was tentatively assigned to Panochthus cf. intermedius, which is one of the 2 species of Panochthus corresponding to the Ensenadan lapse. However, Zurita et al. (2011) recently described remarkably complete material from Sacaba Formation in Bolivia (Early Pleistocene), belonging to P. intermedius. This taxonomic classification was mainly based on the characters observed in the dorsal carapace, which is very similar to that of the holotype of Panochthus intermedius (MLP 16-36), consisting in an almost complete dorsal carapace coming from the Ensenadan levels of Buenos Aires Harbor.
A comparative study between the Bolivian material and MLP 84-IX-2-11 clearly indicates that both differ morphologically. For example, the zygomatic arch in MLP 84-IX-2-11 is more robust; the lambdoid or parieto-occipital crest is more developed; the alveolar ridge of the maxillae is irregular and elevated in the middle. Therefore, we discard the assignment of the material described herein to Panochthus cf. intermedius. As it has been mentioned, the other species of the genus Panochthus that comes from Ensenadan levels of the Pampean Region is P. subintermedius, whose skull has not been preserved, thus precluding comparisons between homologous materials.
However, taking into account that the other species of the Ensenadan Age/Stage, P. subintermedius, was a large-sized animal (Cruz et al., 2011), and that the skull MLP 84-IX-2-11 undoubtedly corresponds to a large species of Panochthus, the most parsimonious taxonomic hypothesis is to assign MLP 84-IX-2-11 to Panochthus cf. subintermedius.
Until 50 years ago, the Ensenadan (Early-Middle Pleistocene) used to be characterized by the presence of taxa larger than the ones commonly found in any other lapse (Scillato-Yané and Carlini, 1998). Later revisions could not confirm this hypothesis for all cases (Zamorano et al., 2010). However, in the case of the genus Panochthus, the premise of the "Ensenadan gigantism" seems to be valid. As mentioned, the 2 known species of Panochthus that were found exclusively in Ensenadan times, P. intermedius and P. subintermedius, are markedly larger than those of the Bonaerian (Middle-Pleistocene) and Lujanian (only registered in the late Pleistocene). In agreement with this hypothesis, the material presented here (MLP 84-IX-2-11) undoubtedly corresponds to one of the larger Glyptodontidae ever known (see Scillato-Yané and Carlini, 1998; Tonni et al., 1999; Zamorano and Scillato-Yané, 2006; Zamorano, 2009). However, the material from the Early Pleistocene of Bolivia, which was assigned to P. intermedius by Zurita et al. (2011), is comparable in size to the above-mentioned species of Panochthus from the Bonaerian-Lujanian lapse (see Table 1). Therefore, the specimen of Panochthus found in the Bolivian altiplano, does not necessarily conform to the premise of "Ensenadan gigantism", which applies to the Panochthus specimens found in the Pampean Region of Argentina. It can be hypothesized that the reason for that lies in an adaptation of P. intermedius to the conditions of the Bolivian altiplano. Accordingly, Pujos (2008) states that some high-altitude members of the Megatheriinae Subfamily are of smaller size than those from low altitude regions, with the exception of Megatherium (Pseudomegatherium) celendinense Pujos (a particularly large species of Megatherium, from the northern Peruvian Andes; see Pujos, 2006; additionally see Pujos and Salas, 2004). Finally, Zurita et al. (2012) indicate that some specimens of Glyptodon restricted to Andean areas are of smaller size than their counterparts of the Pampean Region (see also Rodriguez et al., 2008).
We can conclude: a) the material presented in this paper corresponds to the first known skull of Panochthus from the Ensenadan Age/Stage of the Pampean Region (Argentina), and is, in turn, the second for this genus in the corresponding lapse; b) comparative studies with other species of the genus preclude its assignment to any of the current species; albeit the cranium of P. subintermedius is unknown; c) in this scenario, and taking into account that P. subintermedius is a large-sized species, as is the material studied here, the most parsimonious hypothesis is to classify this specimen as Panochthus cf. subintermedius, in the hope that future findings will confirm it or refute it, and d) the material presented in this paper exhibits some characters not previously observed in any other Glyptodontidae, in particular the imbrications of the upper molariforms.
Acknowledgments
We acknowledge A. A. Carlini for his critical comments on the anatomical description; F. Góis, for providing information on molariform teeth imbrication in the Pampatheriidae; E. Soibelzon, for photographing the material; J. C. González for collaboration on editing the photos; M. Sosa, for sketching the dentition; S. C. Ferris (associate editor) and 2 anonymous reviewers for their thorough reviews and helpful suggestions; J. Pan translated the manuscript to English.
Literature cited
Ameghino, F. 1888. Rápidas diagnosis de mamíferos fósiles nuevos de la República Argentina. Obras Completas 5: 471-480. [ Links ]
Ameghino, F. 1895. Sur les édentés fossiles de l'Argentine. Examen critique, révision et correction de l'ouvrage de Mr. R. Lydekker: "The extinct edentates of Argentina". Revista del Jardín Zoológico de Buenos Ayres 3:97-192. [ Links ]
Ameghino, F. 1909. Le Diprothomo platensis, un précurseur de l'homme. Anales del Museo Nacional de Buenos Aires 19:107-209. [ Links ]
Burmeister, G. 1866. Lista de los mamíferos fósiles del terreno diluviano. Anales del Museo Público de Buenos Aires 1:121-232. [ Links ]
Burmeister, G. 1874. Monografía de los Glyptodontes en el Museo Público de Buenos Aires. Anales del Museo Público de Buenos Aires 2:1-412. [ Links ]
Castellanos, A. 1925. Breves notas sobre evolución de la coraza y del extremo caudal en los Loricata (= Hicanodonta). Revista de "El Círculo", número extraordinario. p. 93-96. [ Links ]
Castellanos, A. 1937. Los sedimentos del Pampeano inferior y del Araucano en el Valle de Los Reartes (Sierra de Córdoba). Publicaciones de la Facultad de Ciencias Matemáticas, Físico-Químicas y Naturales aplicadas a la Industria, (serie Técnico-Científica) 6:1-110. [ Links ]
Castellanos, A. 1942. A propósito de los géneros Plohophorus, Nopachthus y Panochthus (3a. parte). Publicaciones del Instituto de Fisiografía y Geología (Rosario) 11:413-592. [ Links ]
Cione, A. L., E. P. Tonni, M. Bond, A. A. Carlini, U. F. J. Pardiñas, G. J. Scillato-Yané, D. Verzi and M. G. Vucetich. 1999. Ocurrence charts of Pleistocene mammals in the Pampean area, eastern Argentina. In Quaternary of South America and Antarctic Peninsula, Special Volume 12, Quaternary Vertebrate Paleontology in South America, E. P. Tonni and A. L. Cione (eds.). A. A. Balkema, Rotterdam. p. 53-73 [ Links ]
Cruz, L. E., M. Zamorano and G. J. Scillato-Yané. 2011. Diagnosis and redescription of Panochthus subintermedius Castellanos (Xenarthra, Glyptodontia) from the Ensenadan (early-middle Pleistocene) of Buenos Aires (Argentina). Palaeontologische Zeitschrift 85:115-123. [ Links ]
Gillette, D. D. and C. E. Ray. 1981. Glyptodonts of North America. Smithsonian Contributions to Palaeobiology 40:1-251. [ Links ]
Góis, F., G. J. Scillato-Yané, A. A. Carlini and M. Ubilla. 2012. Una nueva especie de Holmesina Simpson (Xenarthra, Cingulata, Pampatheriidae) del Pleistoceno de Rondônia, sudoeste de la amazonia, Brasil. Revista Brasileira de Paleontologia 15:211-227. [ Links ]
Lydekker, R. 1895. Contributions to knowledge of the fossil vertebrates of Argentina. 2. The extinct edentates of Argentina. Anales del Museo de La Plata (Paleontología Argentina 3:1-118. [ Links ]
Owen, R. 1845. Descriptive and illustrated catalogue of the fossil organic remains of Mammalia and Aves. Museum of the Royal College of Surgeons of London. London. 391 p. [ Links ]
Porpino, K. O. and L. P. Bergqvist. Novos achados de Panochthus (Mammalia, Cingulata, Glyptodontoidea) no Nordeste do Brasil. Revista Brasileira de Paleontologia 4:1-62. [ Links ]
Pujos, F. 2006. Megatherium celendinense sp. nov. from the Pleistocene of the Peruvian Andes and the phylogenetic relationships of Megatheriines. Paleontology 49:285-306. [ Links ]
Pujos, F. 2008. Paleogeographic distribution and anatomical adaptations in Peruvian megatheriine ground sloths (Xenarthra: Megatherioidea). In The biology of the Xenarthra, S. F. Vizcaíno and W. J. Loughry (eds.). University Press of Florida, Gainesville. p. 56-63. [ Links ]
Pujos, F. and R. Salas. 2004. A systematic reassessment and paleogeographic review of fossil Xenarthra from Peru. Bulletin de Institut Français d'Études Andines 33:331-377. [ Links ]
Rodriguez, M. A., M. A. Olalla-Tárraga and B. A. Hawkins. 2008. Bergmann's Rule and the geography of mammal body size in the Western Hemisphere. Global Ecology and Biogeography 17:274-283. [ Links ]
Scillato-Yané, G. J. 1995. Xenarthra. Origen y evolución de la fauna de mamíferos en Sudamérica. X Jornadas Argentinas de Mastozoología, La Plata, 1995, resúmenes: 103-104. [ Links ]
Scillato-Yané, G. J. and A. A. Carlini. 1998. Un gigantesco gliptodonte en los alrededores de la ciudad de La Plata. Revista Museo, Fundación Museo de La Plata. 11:45-48. [ Links ]
Sisson, S. and J. D. Grossman. 1959. Anatomía de los animales domésticos. Salvat Editores, Barcelona. 952 p. [ Links ]
Tauber, A. A. and M. E. Palacios. 2007. Nuevos registros de mamíferos cuaternarios de gran porte en la provincia de Santa Cruz, República Argentina. Ameghiniana 44:41R. [ Links ]
Tonni, E. P., P. Nabel, A. L. Cione, M. Etchichury, R. Tófalo, G. J. Scillato-Yané, J. San Cristóbal, A. A. Carlini and D. Vargas. 1999. The Ensenada and Buenos Aires formations (Pleistocene) in a quarry near La Plata, Argentina. Journal of South American Earth Sciences 12:273-291. [ Links ]
Zamorano, M. 2005. Las especies de Xenarthros de la Región Pampeana comunes a las edades Ensenadense y Bonaerense. Bienal de Ciencia y Tecnología (Buenos Aires 2005). Área Biología y Ecología. (Ed. formato digital). [ Links ]
Zamorano, M. 2009. Estudio de un cráneo de Panochthus del Ensenadense (Pleistoceno temprano-medio) comparado con otros del mismo género del Bonaerense y Lujanense (Pleistoceno medio-Holoceno temprano), mediante análisis por componentes principales. Reunión anual de comunicaciones de la Asociación Paleontológica Argentina y Conferencias: "Darwin, Lamarck y la teoría de la evolución de las especies, resúmenes:76. [ Links ]
Zamorano, M.2012. Los Panochthini (Xenarthra, Glyptodontidae): sistemática y evolución. Tesis Doctoral, Universidad Nacional de La Plata, La Plata, Argentina, 269 p. [ Links ]
Zamorano, M. and G. J. Scillato-Yané. 2006. Descripción comparativa del cráneo e Panochthus cf. intermedius Lydekker (Glyptodontidae, Hoplophorinae, Panochthini), del Ensenadense de la región pampeana. 26 Jornadas Argentinas de Paleontología de Vertebrados, (San Juan), resúmenes:36. [ Links ]
Zamorano, M., E. Soilbenzon, G. J. Scillato-Yané and A. E. Zurita. 2010. Fenómenos de gigantismo en Panochthus Burmeister (Xenarthra, Glyptodontidae, Panochthini) del Ensenadense. XXV Jornadas Argentinas de Paleontología de Vertebrados, (San Luis, 2010), resúmenes:24 [ Links ]
Zamorano, M. and D. Brandoni. 2012. Análisis filogenético de los "Panochthini" (Glyptodontidae, "Holplophorinae"). 26 Jornadas Argentinas de Paleontología de Vertebrados, (Buenos Aires), Resúmenes: en formato digital. [ Links ]
Zurita, A. E. 2007. Sistemática y evolución de los Hoplophorini (Xenarthra, Glyptodontidae, Hoplophorinae. Mioceno tardío-Holoceno temprano). Importancia bioestratigráfica, paleobiogeográfica y paleoambiental. Tesis doctoral, Universidad Nacional de la Plata, 367 p. [ Links ]
Zurita, A. E., M. Zamorano, G. J. Scillato-Yané, L. R. González-Ruiz, B. Rivas Durán and R. Céspedez Paz. 2011. An exceptional Pleistocene specimen of Panochthus Burmeister (Xenarthra, Glyptodontoidea) from Bolivia: its contribution to the understanding of the early-middle Pleistocene Panochthini. Comptes rendus Palevol 10:655-664. [ Links ]
Zurita, A. E., A. R. Miño-Boilini, A. Francia, and J. E. Arenas-Mosquera. 2012. The Pleistocene Glyptodontidae Gray, 1869 (Xenarthra: Cingulata) of Colombia and some considerations about the South American Glyptodontinae. Revista Brasileira de Paleontologia 15:273-280. [ Links ]

