










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de biodiversidad
On-line version ISSN 2007-8706Print version ISSN 1870-3453
Rev. Mex. Biodiv. vol.84 n.1 México Mar. 2013
https://doi.org/10.7550/rmb26091
Ecología
The biological and reproductive parameters of the invasive armored catfish Pterygoplichthys disjunctivus from Adolfo López Mateos El Infiernillo Reservoir, Michoacán-Guerrero, Mexico
Parámetros biológicos y reproductivos del bagre armado invasivo Pterygoplichthys disjunctivus del embalse Adolfo López Mateos El Infiernillo, Michoacán-Guerrero, México
Rebeca Aneli Rueda-Jasso1*, Antonio Campos-Mendoza1, Francisco Arreguín-Sánchez2, Edmundo Díaz-Pardo3 and Carlos Antonio Martínez-Palacios4
2 Centro Interdisciplinario de Ciencias Marinas, Instituto Politécnico Nacional, Unidad-La Paz. Ave. Instituto Politécnico Nacional S/N, Playa Palo de Santa Rita, 23096 La Paz, Baja California Sur, México.
3 Facultad de Ciencias Naturales, Universidad Autónoma de Querétaro. Querétaro, Querétaro, Ave. de la Ciencias S/N, Juriquilla, Del. Sta. Rosa de Jauregui, 76230 Querétaro, Querétaro, México.
4 Instituto de Investigaciones Agropecuarias y Forestales, Universidad Michoacana de San Nicolás de Hidalgo. Avenida San Juanito Itzícuaro S/N, San Juanito Itzícuaro, 58330 Morelia, Michoacán, México.
Recibido: 30 junio 2011
Aceptado: 18 julio 2012
Abstract
Invasive Pterygoplichthys disjunctivus (Weber, 1991) (synonymy: Liposarcus multiradiatus) armored catfish were first reported in Mexico in 1995 and have spread successfully in several aquatic ecosystems. In Adolfo López Mateos El Infiernillo Reservoir, Michoacán - Guerrero, the population of this armored catfish is growing substantially because natural predators are not present and exploitation by humans does not occur. Over a complete annual cycle, we studied the following population parameters: weight, total and standard length, the relationship of weight to total length, the hepatosomatic (HSI) and gonadosomatic (GSI) indices, total fecundity (TF) and relative fecundity (RF). The average total (TL) and standard lengths (SL) were 248 ± 37.5 and 188 ± 30.7 mm, respectively, and the average weight (W) was 135.3± 66.8 g. The largest fish reported was 520 mm in TL and weighed 1 280 g. The species showed an extended reproductive season from May to November, as indicated by the GSI, TF and RF. The strongest reproductive peak occurred from July through October. During the annual cycle, the highest monthly average fecundity was 2 447 eggs, and the average relative fecundity was 12.6 eggs per g of fish (August and September). This information will be crucial for calculating the current biomass and future growth of the population. The calculation of these results could provide a basis for the exploitation of this resource for human consumption and animal feeds.
Key words: Loricariidae, armored catfish, invasive species, biological and reproductive parameters, control strategy.
Resumen
La especie invasiva "bagre armado" P. disjunctivus (Weber 1991) (sinonimia de Liposarcus multuradiatus) fue registrada por primera vez en México en 1995; en diversos sistemas acuáticos se ha distribuido exitosamente. En la presa Adolfo López Mateos El Infiernillo, Michoacán - Guerrero, la población de peces armados se ha incrementado considerablemente debido a la ausencia de depredadores y la falta de aprovechamiento. Por ello, a lo largo de un ciclo anual evaluamos los siguientes parámetros: longitud total (LT) y estándar (LS), peso (W), relación peso-longitud total, índices hepatosomático (HSI) y gonadosomático (GSI), fecundidad total (FT) y relativa (FR). La longitud total y estándar promedio fueron 248 ± 37.5 y 188 ± 30.7 mm respectivamente y el peso promedio de 135.3 ± 66.8 gr. El pez de mayor talla presentó 520 mm TL y peso 1 280 gr. La especie presenta una amplia temporada reproductiva, que de acuerdo con GSI, TF y RF se extiende desde mayo hasta noviembre. El mayor pico reproductivo ocurre de julio a octubre. Durante el ciclo anual, la mayor fecundidad promedio fue de 2 447 huevos y la fecundidad relativa fue de 12.6 huevos por gr. de pez (agosto y septiembre). Esta información es fundamental para estimar la biomasa y el crecimiento de la población, con lo cual se pueden plantear estrategias de manejo y explotación del recurso para consumo humano y animal.
Palabras clave: Loricaridae, plecos, especies invasivas, parámetros biológicos y reproductivos, estrategias de control.
Introduction
The family Loricariidae, originally from South America, is considered the largest catfish family, with 680 recognized species (Armbruster and Page, 2006). The sailfin catfish genus Pterygoplichthys originally inhabited the entire Amazon basin (Peru, Bolivia and Brazil) (Weber, 1992; Page and Robins, 2006). As a result of the inadequate control of the pet-fish trade, members of this family have been accidentally or intentionally introduced into several countries (Hoover et al., 2004), where they have adapted successfully. Records have been reported from Texas, Nevada and Florida, USA (Fuller et al., 1999; Capps, 2008, Gibbs et al., 2008; Pound et al., 2011), Puerto Rico (Bunkled-Williams et al., 1994), Taiwan (Liang et al., 2005, 2006), the Philippines (Chávez et al., 2006), Vietnam (Levin et al., 2008), Java and Australia (Capps, 2008), Bangladesh (Hoosain et al., 2008), Turkey (Ozdilek, 2007), Poland (Keszka et al., 2008) and Italy (Piazzini et al., 2010); a recent Danube invasion has also been reported (Simonovic et al., 2011). Several studies report the invasion of loricariids in Mexico (Guzman and Barragan, 1997; Mendoza et al., 2007; Okolodkov et al., 2007; Wakida-Kusonoki et al., 2007; Wakida-Kusunoki and Amador-del Angel, 2008; Capps et al., 2011).
Loricariids have successfully invaded new habitats because these fish are protected by modified scales and by strong spines on the fins (Armbruster and Page, 2006) and because they show a high tolerance to low oxygen concentrations or desiccation (up to 20 hours). The latter ability can be attributed to an enlarged and vascularized stomach, which functions as an accessory respiratory organ (Armbruster, 1998). The fish have multiple-spawning reproduction, nest construction and parental care (Tello et al., 1992). Additionally, Pterygoplichthys has proven to have a high tolerance to salinity (up to 10 parts per thousand) (Capps et al., 2011).
In Mexico, the sailfin catfish was detected for the first time in the Mezcala River, Guerrero (Balsas Basin), in 1995 (Guzmán and Barragán, 1997) and later in Tecpatan, Chiapas, the surroundings of Villahermosa, Tabasco (Mendoza et al., 2007), and in the Grijalva-Usumacinta Basin (Tabasco and Chiapas) (Wakida-Kusunoki et al., 2007; Wakida-Kusunoki and Amador del-Angel, 2008). The first specimens were identified as Liposarcus multiradiatus, a synonym of Pterygoplichthys multiradiatus (Armbruster, 2004); however, the species is currently identified as P disjunctivus (Armbruster pers. comm.).
Invasive sailfin catfish have impacted fish communities and fisheries in several countries. Several previous studies have suggested that competition between this invasive fish and the native species can produce negative impacts on the local fisheries (Keszka et al., 2008). Pterygoplichthys pardalis showed overlapping isotope signatures with native species, including some high-value fishery species (Mendoza-Carranza et al., 2010). In Florida, interactions between Pterygoplichthys and native manatees have been recorded. The catfish attached to the manatees and grazed on the epibiota of their skins. Although it is unclear whether this behavior is damaging to the manatees, some manatees were observed in acts to remove the fish (Nico et al., 2009).
Armored catfish (P disjunctivus) have been found in El Infiernillo Reservoir, located in the states of Michoacán and Guerrero. In view of the proliferation of the species in the study area, we investigated selected biological and reproductive parameters of the population. This information may contribute to the understanding of the reproductive success of this invasive species and may indicate the reasons for the proliferation of the population. This information could be used to plan possible control alternatives, such as fishing for consumption.
Materials and methods
Study area. El Infiernillo Reservoir is located in the Balsas Basin, between the states of Michoacán and Guerrero (18°47" and 17°55' N, 101°47" and 102°11' W), at an elevation of 150 m above sea level (INEGI, 1985) (Fig. 1). El Infiernillo is 120 km in length and contains an average of 11 860 000 m3 of water. It was built in 1963 to serve a hydropower station. The inundated surface can reach 40 000 ha, and the average depth of the reservoir is 35 m (Juárez-Palacios, 1995).
Fish sampling. A total of 7 731 P. disjunctivus were obtained in monthly samples collected between April 2007 and April 2008. The fish were captured as bycatch in the tilapia fishery; 4 sites were selected: a) Churumuco (CHU), b) La Obra (LO), c) Nuevo Centro (NC) and d) Pinzandaran (PIN) (Fig. 1). The sampling sites were selected because they are the most important landing areas for fishermen. The captures were performed at random points located within a 3 to 6 km radius in the reservoir. At each sampling site, 4 fishermen supplied their total sailfin fish catch. The fish were captured using 2.5 to 3.25 inch (6.5 - 8.3 cm) mesh gill nets. The fishing began 2 hours before sunrise and ended 3 to 4 hours after sunrise. All of the fish were measured with an ichthyometer to determine the total (TL) and standard length (SL) and wet-weighed (W) on a digital balance (Ohaus Corporation, Scout Pro SP2001, NJ, USA; precision of 0.01 g). Ten fish from each sampling point were dissected. The gonads and liver were extracted to calculate the gonadosomatic and hepatosomatic indices. Both organs were weighed on a digital balance (Ohaus Corporation, Scout Pro SP202, NJ, USA; precision of 0.001 g) and fixed with 10% formalin. The hepatosomatic (HSI) and gonadosomatic (GSI) indices were calculated as follows:
HSI= (liver weight [g] /fish weight [g]) * 100
[g] /fish weight [g]) * 100
All of the measurements of TL and W were used to calculate the average and standard deviation of TL and W and the weight-total length relationship. To perform comparative analyses of the TL, W, HSI and GSI by locality and season, we selected (using random numbers) and grouped 20 values of TL and W by season for each sampling site, whereas only 8 values were used for the HSI and GSI. The comparisons were performed using a two-way ANOVA (Factor A was the season, Factor B the locality) with JMP Ver 6.0. If differences were observed, a Tukey post hoc analysis was performed.
To evaluate the total fecundity (TF), the left gonad (heavier and larger) of each fish was dissected, and the oocytes were passed through a 500 u.m sieve; only the vitellogenic oocytes (usually present in preparation for ovulation during the current reproductive season) were individually counted. A tally counter (JQ-14A Fenghua Guangya Counter Manufacturing Co, China) was used to count these oocytes. The relative fecundity (RF) was calculated as the total number of eggs estimated for both gonads as twice the number of vitellogenic oocytes found in the left gonad divided by the weight of the female (the data are reported as the number of eggs per g body weight). For the TF and RF, the data for the 4 localities were grouped, and the average values ± standard deviations were graphed using Microsoft Excel. The duration of the reproductive period was determined using the highest values of the monthly GSI, TF and RF as the criteria.
The length-weight relationships were estimated for each locality using the linear form of the formula W= aLb (Le Cren, 1951), log W= log a + b log L. W denotes the weight of the fish, L denotes the total length of the fish and a and b are constants. The values of the constants a and b were estimated using the method of least squares. A t test was employed to evaluate the significance of the differences in the weight-length relationship among the localities. JMP Ver. 6 was used for the statistical analysis.
Results
On average, the largest and heaviest fish were reported at PIN (270 ± 37.3 mm TL, and 173.0 ± 69.1 g W), whereas the smallest fish were recorded at NC (199.5 ± 35.7 mm and 75.5 ± 20.2 g total length and weight, respectively) (Table 1). The largest armored catfish collected at CHU was 520 mm in TL and 1 280 g in W. Although the two-way ANOVA for the TL and W showed no differences if the data were compared seasonally, the comparisons by locality showed that the smallest fish (TL and W) were recorded at NC and that LO, CHU and PIN grouped together with similar results (Fig. 2). The total length and weight were strongly correlated (R2= 0.86 - 0.90). This relationship did not differ significantly among the localities (t test) (Table 1). The HSI values indicated that the hepatic condition of the fish from the different localities was similar (Fig. 3A). The lowest average HSI occurred during the summer, whereas the spring, autumn and winter HSI values were similar (Fig. 3B). The GSI values were highest at PIN, followed by LO and NC, which grouped together, and CHU, with the lowest value (Fig. 3C). The summer and autumn GSI values were significantly higher than the spring and winter values (Fig. 3D).
Fish with vitellogenic eggs were recorded throughout the year but were scarce during the spring and winter. If all of the GSI data were grouped by month, the highest average GSI was observed in September (9.51), followed by August (9.02), October (4.77), July (4.74), June (1.79), and May (1.45). The GSI values were lower than 0.8 for the other months. Calculating all GSI percentages using the highest GSI (September) as reference value, a gradual decrease was observed (August 94.8, October 50.1, July 49.8, June 18.8, May 15.1). The lowest percentage occurred in January (3.76%), and the percentage increased again in April 2008 (29%).
The highest individual value of TF was 6 686 eggs. This value was observed in a specimen (TL 278 mm, body weight 196 g) collected in June. The highest average TF was recorded during August (2 447 ± 252 eggs), followed by July, June, September, May, and October; the average TF for this period (May to October) was 1 856 ± 499.6 eggs. The lowest individual TF value was 40 eggs, found in a specimen (TL 221 mm, W 62.7 g) collected in January. The lowest average TF was observed in January (345 ± 94 eggs), and the average was 655 ± 326.6 for the period from November to March (Fig. 4A).
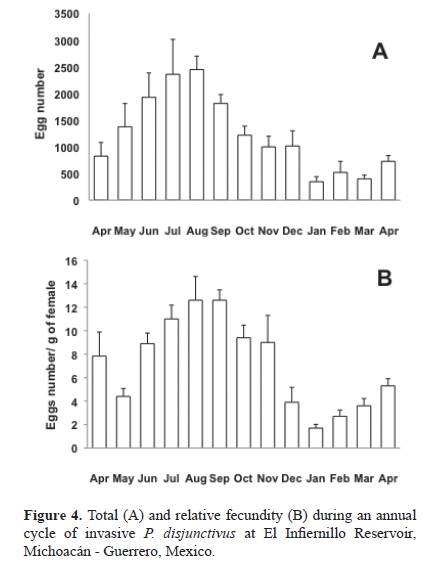
The highest relative fecundity was 12.6 ± 2.02 eggs, in August and September. The average RF was 9.81 ± 3.07 for the months of high reproductive activity (July to September) (Fig. 4B). The months during which reproduction was low (November to April) showed an average RF of 2.9 ± 0.9.
Discussion
Members of the invasive genus Pterygoplichthys show expanding distributions. The populations of these invasive species grow exponentially because they lack natural predators and because they are not used as food. The environmental impacts of these expansions have not yet been completely identified, although several studies have reported evidence of such impacts (Mendoza et al., 2007; Nico et al., 2009; Rueda-Jasso et al., 2009; Mendoza-Alfaro et al., 2011). However, certain basic aspects of the structure of these populations must be understood before a control or utilization strategy can be planned.
A substantial sample (7 731 fish) of P. disjunctivus was obtained during the annual cycle. The high abundance of this species may reflect the characteristics of the aquatic environment at the dam. The water has an average temperature of 29° C, a pH of 9.4, 8.6 mg/L of oxygen, a transparency of 1 m, and total dissolved solids (TDS) of 276 mg/L. The rainy season extends from June to September (Campos-Mendoza et al., 2011, http://smn.cna.gob.mx). These parameters are similar to those of the original habitat of the armored catfish. The Orinoco River exemplifies this original habitat: the rainy season extends from May to October, the water temperature reaches 23 to 28° C and the water transparency is 1-3 m (Montaña et al., 2010). The number of fish collected by the present study was significantly higher than the number of P. pardalis captured in the Palizada River, in southeastern Mexico (118 during an annual cycle) (Wakida-Kusunoki and Amador-del Angel, 2011). These authors found no differences in the abundance of this species during a daytime period beginning at 14 h and ending at 21 h. Nico (2010) found that adult P. disjunctivus were active throughout the day, whereas juveniles were passive during the daylight hours, perhaps to avoid predators. In our experience (preliminary sampling), the number of armored catfish was low during the daylight hours. The fishermen at El Infiernillo, stated that increases in the time spent fishing during the hours before sunrise result in an increase in the catch of armored catfish and a decrease in the catch of tilapia.
Heavier and longer (weight, total and standard length) fish were collected at PIN, CHU and LO, whereas the fish captured at NC were significantly lower in size and weight. This finding could result from the selectivity of the mesh size of the gill nets for tilapia. Although a regulation NOM-027-PESC-2000 (SEMARNAT, 2000) has existed since 2000 to limit the net sizes, the net number per fisherman and the hours of fishing, our results suggest that the net sizes are smaller than those recommended in such locations as NC. The armored catfish examined in this study were captured in the gill nets used to catch tilapia. To obtain a representative sample of the P. disjunctivus population at El Infiernillo Reservoir, complementary types of fishing equipment (electrofishing, seines and gill nets of different mesh sizes) must be used.
The size range of the armored catfish found in our study was similar to that of Liposarcus multiradiatus in the Kaoping River in Taiwan (Liang et al., 2005) and P. disjunctivus in Florida (Gibbs et al., 2008). However, the average size of P. pardalis captured with gill nets of 76 mm (3 inches) in the Palizada River (Wakida-Kusunoki and Amador-del Angel, 2011) was larger than that of the P. disjunctivus captured at El Infiernillo.
The b values for the relationship between the total length and the weight ranged between 2.3 - 2.7. These values indicate negative allometric growth. This finding is consistent with the results of Liang et al. (2005), Samat et al. (2008), and Wakida-Kusunoki and Amador-del Angel (2011). It has been suggested that the b values show ongoing variation in response to several intrinsic and extrinsic factors, even for the same species (Samat et al., 2008). For this reason, it is recommended that future studies examine this issue further.
In the present study, we found that highest reproductive activity occurred from July to September (summer). However, calculating all GSI, TF and RF using the highest reproductive months as reference values, the peak of the reproductive period was found to include 5 months, from June to October, based on those values that were > 50%. Nevertheless, the total and relative fecundity values for May and November were still high. Note that we recorded fish with ripe gonads throughout the year. Our results are consistent with the extension of the reproductive period for the same species in Florida, although the reproductive period in Florida began and ended a month earlier, extending from May to September (Gibbs et al., 2008). Similarly, the reproductive period of an invasive population of L. multiradiatus in Taiwan extended from March to September (Liang et al., 2005), and P. pardalis reached reproductive peaks from June to September (Wakida-Kusunoki and Amador-del Angel, 2011).
The hepatosomatic index did not show differences between the localities, whereas differences were observed for the GSI. Accordingly, the relationship between the hepatosomatic index and the gonadosomatic index remains unclear. However, the HSI values were higher during the spring and winter because the fish were storing energy for reproduction. This finding is consistent with the low GSI observed during the spring and winter. During the summer, an inverse effect was observed as energy was transferred from the liver to the gonads, resulting in a high GSI and a low HSI. An inverse relationship between the HSI and the GSI has been observed in several tropical and temperate fish, such as Lutjanus gutattus (Arellano-Martinez et al., 2001), Lutjanus peru (Gallardo-Cabello et al., 2010), Oreochromis niloticus (Gomez-Marquez et al., 2003) and Pingues chilensis (Gonzalez and Oyarzun, 2002).
The autumn GSI and HSI values of the armored catfish were high. A possible explanation would be that certain fish continued to reproduce after the reproductive maximum. At that time, energy was resupplied to the liver in the fish that had completed reproduction. In another species, Salmo salar, the occurrences of increases or decreases in the HSI depended on the feeding level over a period of 2 - 4 days (Arndt, 2000).
In addition, it has been hypothesized that in tropical regions, the rainy season influences the limnological parameters (the load of nutrients and phyto- and zooplankton production) in ways that ultimately yield an abundance of food and facilitate fish reproduction (Peterson et al., 2004). In our results for El Infiernillo, the rainy season coincided with the reproductive period of the armored catfish (http://smn.cna.gob.mx). For Hypostomus affinis (Loricariidae), the period of ovarian development extended from September to February and was related to the rainy season (Mazzoni and Caramaschi, 1997).
The size at first maturity of the armored catfish in Florida was reported to be 260 mm, whereas the size at first maturity was 192 mm in our study. This difference could be due to the higher temperature reported at the El Infiernillo Reservoir, whereas lower temperatures could have delayed the onset of puberty and reproduction in the Florida population of P. disjunctivus.
At the El Infiernillo Reservoir, the P. disjunctivus population has found an appropriate habitat in which a lack of predators and exploitation, a surplus of available food and the presence of parental care, as well as a long reproductive season have produced high recruitment and successful colonization. The region around the El Infiernillo reservoir is undeveloped, and many families depend on the tilapia fishery. Therefore, P. disjunctivus has been considered a pest, and fishermen commonly discard these fish directly into the water and at landing areas, generating contamination. In South America (i.e., Brazil, Colombia and Venezuela); however, this species is viewed as a high-quality item for human consumption (Martínez-Palacios et al., 2010a, b). To understand more about the population structure of this alien species and develop efficient strategies for its utilization, it is necessary to continue research on the population parameters. Studies performed in parallel to the current investigation and conducted by members of the same research group have concluded that P. disjunctivus has potential uses as a human food fish in the form of fresh fillets, processed meat "surimi" and caviar. All of these products performed well in palatability tests and showed good nutritional quality. Furthermore, the byproducts from the fish can be processed as silage for animal diets (Martínez-Palacios et al., 2010a, b).
Acknowledgments
This work was funded by CONACYT through the Fondos Mixtos CONACYT-Gobierno del Estado de Michoacán grant numbers, 37147 and 53437, and PIFI-UMSNH project 2010. We would like to thank the fishermen who helped with the fish samples. We also appreciate the help of Dr. Jonathan Armbruster, from Auburn University, USA, who helped us to identify the fish, and Dr. Javier Ponce, from Universidad Michacana, who advised us on the statistical analysis. Finally, we would like to thank the reviewers for their excellent recommendations, which helped to improve this report.
Literature cited
Arndt, S. K. A. 2000. Influence of sexual maturity on feeding, growth and energy stores of wild Atlantic salmon parr. Journal of Fish Biology 57:589-596. [ Links ]
Arellano-Martinez, M., A. Rojas-Herrera, F. García-Domínguez, B. Ceballos-Vázquez, and M. Villalejo-Fuerte. 2001. Ciclo reproductivo del pargo lunarejo Lutjanus guttatus (Steindachner, 1869) en las costas de Guerrero, México. Revista de Biología Marina y Oceanografía 36:1-8. [ Links ]
Armbruster, J. W. 1998. Modifications of the digestive tract for holding air in Loricariid and Scoloplacid catfishes. Copeia 3:663-675. [ Links ]
Armbruster, J. W. 2004. Phylogenetic relationships of the suckedmouth armoured catfishes (Loricariidae) with emphasis on the Hypostominae and Ancistrinae. Zoological Journal of the Linnean Society 141:1-80. [ Links ]
Armbruster, J. W. and L. M. Page. 2006. Redescription of Pterygoplichthys punctatus and description of a new species of Pterygoplichthys (Siluriformes: Loricariidae). Neotropical Ichthyology 4:401-409. [ Links ]
Bunkled-Williams, L., H. E. Williams, J. R. Craig, G. Lilystrom, I. Corujo-Flores, J. A. Zerbi, C. Aliaume and T. N. Churchill. 1994. The South America sailfin catfish Liposarcus multiradiatus (Hancock), a new exotic established in Puerto Rico fresh waters. Caribbean Journal of Sciences 30:90-94. [ Links ]
Campos-Mendoza, A., E. Diaz-Pardo and A. Gutierrez-Hernández. 2011. Informe final del proyecto "Estudio prospectivo del estado ecológico y productivo del embalse Adolfo López Mateos: una propuesta para un adecuado manejo biológico, pesquero y de acuicultura" FOMIX CONACYT-Gobierno del Estado de Michoacán, 53437. [ Links ]
Capps, K. 2008. The impacts of sailfish catfish (Siluriformes: Loridariidae) on invaded freshwater ecosystems. Abstracts of Annual Meeting of American Ichthyologist and Herpetologist Society. 23 - 28 July, Montreal, Canada. [ Links ]
Capps, K. A., G. L. Nico, M. Mendoza-Carranza, W. Arévalo-Frías, A. J. Ropicki, S. A. Heilpern and R. Rodiles-Hernández. 2011. Salinity tolerance of normative suckermouth catfish (Loricariidae: Pterygoplichthys) in south eastern Mexico: implications for invasion and dispersal. Aquatic Conservation: Marine and Freshwater Ecosystems 21:528-540. [ Links ]
Chávez, J. M., R. M. De La Paz, S. K. Manohar, R. C. Pagulayan and J. R. Carandang. 2006. New Philippine record of South America sailfin catfishes (Pisces: Loricariidae). Zootaxa 1109:57-68. [ Links ]
Fuller, P. L., L. G. Nico and G. D. Williams. 1999. Nonindigenous fishes introduced into inland waters of the United States. American Fisheries Society, Special Publication 27, Bethseda, Maryland. p. 435-441. [ Links ]
Gallardo-Cabello, M., M. Sarabia-Mendez, E. Espino-Barr and V Anislado-Tolentino. 2010. Biological aspect of Lutjanus peru in Bufadero Bay, Michoacan, Mexico: growth, reproduction and condition factors. Revista de Biologia Marina y Oceanografia 45:205-215. [ Links ]
Gibbs, M. M., J. H. Shields, D. W. Lock, K. M. Talmadge and T. M. Farell. 2008. Reproduction in an invasive exotic catfish Pterygoplychtyys disjuctivus in Volusia Blue Spring, Florida, USA. Journal of Fish Biology 73:1562-1572. [ Links ]
Gómez-Márquez, J. L., B. Pena-Mendoza, I. H. Salgado-Ugarte and M. Guzman-Arroyo. 2003. Reproductive aspects of Oreochromis niloticus (Perciformes: Cichlidae) at Coatetelco lake, Morelos, Mexico. Revista de Biología Tropical 51:221-228. [Online]: http://www.scielo.sa.cr/scielo.php?script=sci_arttext&pid=S0034-77442003000100020&lng=es&nrm=iso; last access: 28.II.2013. [ Links ]
González, C. and C. Oyarzun. 2002. Variedad de índices biológicos en Pingues chilensis Valenciennes 1833 (Perciformes, Pinguipedidae) ¿Estan realmente correlacionados? Gayana 66:249-253. [ Links ]
Guzmán, A. F. and J. S. Barragán. 1997. Presencia de bagre Sudamericano (Osteichthyes: Loricariidae) en el río Mezcala, Guerrero, México. Vertebrata Mexicana 3:1-4. [ Links ]
Hoosain, M. Y., M. M. Rahman, Z. F. Ahmed, J. Ohtomi and A. B. M. S. Islam. 2008. First record of South America sailfin catfish Pterygoplichthys multiradiatus in Bangladesh. Journal of Applied Ichthyology 24:718-720. [ Links ]
Hoover, J., J. Killgore and A. Cofrancesco. 2004. Suckermouth catfishes: threats to aquatic ecosystems of the United States? In Aquatic Nuisance Species Research Bulletin 4-1. U.S. Army Corp of Engineers. [ Links ]
Instituto Nacional de Estadística Geografía e Informática (INEGI). 1985. Síntesis geográfica del rstado de Michoacán. INEGI. Mexico. 315 p. [ Links ]
Juárez- Palacios, J. R. 1995. Presa Infiernillo (Adolfo López Mateos). In Lagos y presas de México, G. De La Lanza and J. L. C. García (comps.). Centro de Ecología y Desarrollo, A.C. México D. F. p. 211-223. [ Links ]
Keszka, S., R. Panicz and A. Tanski. 2008. First record of the leopard pleco. Pterygoplichthys gibbiceps (Actinopterygii: Loricariidae) in Brda river in the centre of Bydgoszcz (Northern Poland). Acta Ichthyologica et Piscatoria 38:135138. [ Links ]
Le Cren, E. D. 1951. The length-weight relationship and seasonal cycle in gonad weight and condition in the perch (Perca fluviatilis). Journal of Animal Ecology 20:201-221. [ Links ]
Levin, A. B., O. H. Phoung and D. S. Pavlov. 2008. Discovery of the Amazon sailfin catfish Pterigoplichthys pardalis (Castelnau, 1855) (Teleostei: Loricariidae) in Vietnam. Journal of Applied Ichthyology 24:715-717. [ Links ]
Liang, S. H., Wu, H. P. L. and B. S. Shied. 2005. Size structure, reproductive phenology and sex ratio of an exotic sailfish catfish (Liposarcus multiradiatus) in the Kaoping river of southern Taiwan. Zoological Studies 44:252-259. [ Links ]
Liang, S. H., L. Ch. Chuang and M. H. Chuang. 2006. The pet trade as a source of invasive fish in Taiwan. Taiwania 51:93-98. [ Links ]
Martínez-Palacios, C. A., A. Campos-Mendoza, E. Díaz-Pardo, F. Arreguín-Sánchez, R. Rueda-Jasso, J. Fonseca-Madrigal, A. Gutiérrez-Hernández, R. Pacheco-Aguilar, J. C. Ramírez-Suárez, M. G. Ríos-Durán, E. M. Toledo-Cuevas, G. Salas-Razo, L. G. Ross, A. Shimada-Miyasake, M. T. Viana-Castrillón, A. Sánchez-Chinchillas, E. Ávila-González and E. Gasca-Leyva. 2010a. Bagres armados: ¿Erradicación o utilización? Ciencia y Desarrollo 236:28-33. [ Links ]
Martínez-Palacios, C. A., L. G. Ross, F. Arreguín-Sánchez, A. Campos-Mendoza, E. Díaz-Pardo, J. Fonseca-Madrigal, A. Gutiérrez-Hernández, R. Pacheco-Aguilar, J. C. Ramírez-Suárez, M. G. Ríos-Durán, R. Rueda-Jasso, E. M. Toledo-Cuevas, G. Salas-Razo, A. Shimada-Miyasake, A. Sánchez-Chinchillas, E. Ávila-González, M. T. Viana-Castrillón and E. Gasca-Leyva. 2010b. Peces amazónicos invaden aguas continentales de México y Guatemala en forma alarmante. El Salvador Ciencia y Tecnología 15:25-32. [ Links ]
Mazzoni, R. and E. P. Caramaschi. 1997. Spawning season, ovarian development and fecundity of Hypostomus affinis (Osteichthyes, Loricariidae). Revista Brasileira de Biologia 57:455-462. [ Links ]
Mendoza, R., S. Contreras, C. Ramírez, P. Koleff, P. Álvarez and V. Aguilar. 2007. Los peces diablo: especies invasoras de alto impacto. Biodiversitas 70:1-5. [ Links ]
Mendoza R., C. Escalera, S. Contreras, P. Koleff, C. Ramírez, P. Alvarez, M. Arroyo and A. Orbe-Mendoza. 2011. Chapter 5. Invasion of the armored catfish in the Infiernillo reservoir, Michoacan-Guerrero, Mexico. Socioeconomic impacts analysis: a tale of two invaders. Trinational risk assessment guidelines for aquatic alien invasive species. Test cases for the snakeheads (Channidae) and armored catfishes (Loricariidae) in North America inland waters. Commission for Environmental Cooperation. Montreal, Quebec. 51-59 p. [ Links ]
Mendoza-Carranza, M., D. J. Hoeinghause, A. L. García and A. Romero-Rodríguez. 2010. Aquatic food webs in mangrove and seagrass hábitats of Centla wetlands, a biosphere reserve in Southeaster Mexico. Neptropical Ichthyology 8:171-178. [ Links ]
Montaña, C. G., C. A. Layman and D. C. Taphorn. 2010. Inventario de la ictiofauna del Caño La Guardia, afluente del río Capanaparo (cuenca del Orinoco), estado Apure, Venezuela. Biota Colombiana 11:75-88. [ Links ]
Nico, L. G., W. F. Loftus and J. P. Reid. 2009. Interaction between non-native armored catfish (Loricariidae Pterygoplichthys) and native Florida manatee (Trichechus manatus latirostris) in artesian spring. Aquatic Invasions 4:511-519. [ Links ]
Nico, L. G. 2010. Nocturnal and diurnal activity of suckermouth catfish (Loricariidae: Pterygoplichthys) associated with winter Florida manatees (Trichechus manatus latirostris). Neotropical Ichthyology 8:893-898. [ Links ]
Okolodkov, Y. V, R. Bastida-Zavala, A. L. Ibáñez, J. W. Chapman, E. Suárez-Morales, F. Pedroche and F. J. Gutierrez-Mendieta. 2007. Especies acuáticas no indígenas en México. Ciencia y Mar 11:29-67. [ Links ]
Ozdilek, S. Y. 2007. Possible threat for Middle East inland water: an exotic and invasive species, Pterygloplichthys disjuntivus (Weber 1991) in Asi river, Turkey (Pisces: Loriicaridae). Journal of Fisheries and Aquatic Science 24:303-306. [ Links ]
Page, L. M. and R. H. Robins. 2006. Identification of sailfin (Teleostei: Loricariidae) in Southeastern Asia. The Raffles Bulletin of Zoology 54:455-457. [ Links ]
Peterson, M. S., W. T. Slack, N. J. Brown-Peterson and J. L. McDonald. 2004. Reproduction in nonnative environments: establishment of Nile tilapia, Oreochromis niloticus in coastal Mississippi watersheds Copeia 4:842-849. [ Links ]
Piazzini S., E. Lori, L. Favilli, S. Cianfanelli, S. Vanni and G. Manganelli. 2010. A tropical fish community in thermal waters of southern Tuscany. Biological Invasions 12:2959-2965. [ Links ]
Pound, K. L., W. H. Nowlin, D. G. Huffman and T. H. Bonner. 2011. Trophic ecology of a nonnative population of suckermouth catfish (Hypostomus plecostomus) in central Texas spring-fed stream. Environmental Biology of Fishes 90:277-285. [ Links ]
Rueda-Jasso, R. A., A. Campos-Mendoza, C. Entzin-Díaz, E. Mata-Cortés, F. Arreguin-Sánchez y C. A. Martínez-Palacios. 2009. Population parameters of the invasive Pterygoplichthys disjunctivus (Webber 1991) in the reservoir Adolfo López Mateos (El Infiernillo), Michoacan Mexico. 13th European Congress of Ichthyology, Klaipeda, Lithuania. 6 - 12 September 2009. [ Links ]
Samat, A., M. N. Shukor, A. G. Mazlan, A. Arshad and M. Y Fatimah. 2008. Length-weigth relationship and condition factor of Pterygoplichthys pardalis (Pisces: Loricariidae) in Malaysia Peninsula. Research Journal of Fisheries and Hydrobiology 3:48-53. [ Links ]
SEMARNAT (Secretaría de Medio Ambiente y Recursos Naturales). 2000. Norma Oficial Mexicana NOM-027-PESCA-2000. Pesca responsable en la presa Adolfo López Mateos El Infiernillo, Michoacán y Guerrero. Especificaciones para el aprovechamiento de los recursos pesqueros. http://www.oeidrus-portal.gob.mx/oeidrusjalisco/jalisco/docs/serv/norm/NOM-027-PESC-2000.pdf; last access: 28.II.2013. [ Links ]
Simonovic, P., V. Nikolic and S. Grujic. 2011. Amazon sailfin catfish Pterygoplichthys pardalis (Castellnnau, 1855) (Loricariidae, Siluriformes), a new fish species recorded in the Serbian section of the Danube river. Biotechnology and Biotechnology E.Q. 24/2010/Special Edition On Line 655-660. http://www.diagnosisp.com/dp/journals/view_pdf.php?journal_id=1&archive=1&issue_id=27&article_id=1027; last access: 28.II.2013. [ Links ]
Tello, J. S., V. H. Montreuil, J. T. Maco, R. A. Ismiño and H. Sanchez. 1992. Bioecologia de peces de importancia económica de la parte inferior de los ríos Ucayali y Maranon, Peru. Folia Amazonica 4:87-107. [ Links ]
Wakida-Kusunoki, A. T., R. Ruíz-Carus and E. Amador-del Angel. 2007. Amazon sailfin catfish (Castelnau, 1985) (Loricariidae), another exotic species established in southeastern Mexico. Southwestern Naturalist 52:121-144. [ Links ]
Wakida-Kusunoki, A. T. and E. Amador-del Angel. 2008. Nuevos registros de los plecos Pterygoplichthys pardalis (Castelnau, 1855) y P. disjunctivus (Weber, 1991) (Siluriformes: Loricariidae) en el Sureste de México. Hidrobiologica 18:251-256. [ Links ]
Wakida-Kusunoki, A. T. and E. Amador-del Angel. 2011. Aspectos biológicos del pleco invasor Pterygoplichthys pardalis (Teleostei: Loricariidae) en el río Palizada, Campeche, México. Revista Mexicana de Biodiversidad 82:870-878. http://www.fishbase.org/Summary/SpeciesSummary.php?id=51938; last access: 20.VI.2011. [ Links ]
Weber, C. 1992. Revision du genre Pterygoplichthys sensu lato (Pisces, Siluriformes, Loricariidae). Reveu Francaise d Aquariologie et Herpetologie 19:1-36. [ Links ]

