










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de biodiversidad
versión On-line ISSN 2007-8706versión impresa ISSN 1870-3453
Rev. Mex. Biodiv. vol.82 no.3 México sep. 2011
Research note
Sampling necrophagous and predatory insects using different lures in a Mexican pine forest
Muestreo de insectos necrófagos y depredadores utilizando diferentes cebos en un bosque de pino en México
Gerardo Sánchez–Rojas, Ignacio Castellanos* and Amelia Márquez–Vázquez
Centro de Investigaciones Biológicas, Universidad Autónoma del Estado de Hidalgo. Apartado postal 69–1, 42001 Pachuca, Hidalgo, México. * Correspondent: ignacioe@uaeh.edu.mx
Recibido: 30 agosto 2010;
aceptado: 17 diciembre 2010
Abstract
Sampling with a single bait as a lure is a commonly used practice in studies of necrophilous insect communities. Here, we determined if there were differences in necrophilous insect species richness and abundance when attracted to 4 types of carrion: squid, bird, lizard, and mouse. A comparison between necrophagous and predatory insects was also made to detect whether they differ in their abundance, depending on the bait used. Thirteen species of Silphidae, Staphylinidae (Coleoptera), Calliphoridae, and Sarcophagidae (Diptera) were collected in the baited traps; no single bait was able to attract all species. The abundances of 5 species varied significantly depending on the type of bait used. Predatory insects were less abundant than necrophagous insects in lizard and mouse baits. These results show that an evaluation of the structure of a necrophilous insect community can depend on the bait used for sampling.
Keywords: Necrophilous insects, guilds, species accumulation curves, carrion, Mexico.
Resumen
La utilización de un solo tipo de cebo es una práctica común en los estudios de comunidades de insectos necrófilos. En este trabajo determinamos si existen diferencias en la riqueza y abundancia de especies de insectos necrófilos atraídos a 4 tipos de carroña: calamar, ave, lagartija y ratón. También se determinó si la abundancia de insectos necrófagos y depredadores difiere dependiendo del tipo de cebo que se utilice. Se recolectaron 13 especies de Silphidae, Staphylinidae (Coleoptera), Calliphoridae y Sarcophagidae (Diptera); ningún tipo de cebo logró atraer a todas las especies. Se encontró que la abundancia de 5 de las especies difirió significativamente dependiendo del cebo utilizado. La abundancia de los insectos depredadores fue menor que la de los carroñeros en los cebos de lagartija y ratón. Estos resultados muestran que la evaluación de la estructura de una comunidad de insectos necrófilos puede depender del tipo de cebo que se utilice.
Palabras clave: insectos necrófilos, gremios, curvas de acumulación de especies, carroña, México.
Necrophagous insects and their associated predators have been used frequently to investigate various aspects of insect diversity, including studies focused on understanding community structure, and the effects of disturbance on their diversity (Gibbs and Stanton, 2001; Ulrich et al., 2008). The influence of necrophilous insects on certain ecosystem processes, such as the decomposition and relocation of animal organic matter has also received attention (Carter et al., 2007). However, a commonly used practice in many studies of necrophilous insect communities has been the use of a single bait as a lure. Since necrophilous insect species may differ in carcass preferences as well as in their habits, the employment of different baits may result in different number of species and individuals being recorded. Thus, an evaluation of the structure of a necrophilous insect community may depend on the bait used for sampling.
The aim of this study was to investigate whether the abundance and species richness of necrophilous insects (Coleoptera and Diptera) differ when employing different baits. Specifically, a comparison of insects’ richness, abundances, and diversity attracted to 4 types of bait, and an evaluation between the baits in terms of their attractiveness for necrophagous and predatory insects were made.
The study was conducted in a pine–oak forest located within Rancho Santa Elena, Huasca de Ocampo, Hidalgo, Mexico, between 20°06´07’’and 20°09’50’’N, and 98°30’04’’ and 98°32’06’’W, 2 300–2 700 m alt. The climate is temperate to semi humid. The average monthly temperature is 15.1°C, with January and May registering the coldest and warmest months, respectively. The total annual rainfall is 787.6 mm with December and September corresponding to the driest and wettest month respectively (CNA, 2006).
Sampling was carried out during 4 consecutive weeks in June and July of 2004. Traps were constructed using one liter plastic jars with 3 openings (1.5 cm2) cut into the upper walls to allow access to necrophilous insects; the jars were closed with a lid under which was attached a perforated 150 ml plastic container that contained the lure. Traps were buried with their upper section above the soil surface. At each sampling period, each jar was exchanged with another jar filled one–third with 70% ethyl alcohol.
Three different types of bait that represented locally available resources for necrophilous insects were used, lizards (Sceloporus grammicus Wiegmann), mice (Peromyscus maniculatus Wagner), and commercial chicken. Squid, an exogenous but commonly–used luring element in studies of necrophilous insects in Mexico, was also included. Each trap was baited with 40 g of fresh bait which decomposed throughout the sampling period. In the case of squid and chicken, pieces of fresh muscle were used as bait, whereas in the cases of mouse and lizard, pieces of the entire body were employed. A control group of traps without bait was also used, and these contained only 70% ethyl alcohol.
Six replicate traps with each bait as well as 6 controls for a total of 30 traps were randomly placed within 6 transects. Traps were separated by at least 100 m in order to allow for independence between traps (Larsen and Forsyth, 2005). Samples were collected every 7 days over a 4 week period. The study of the necrophagous community was restricted to all trapped species of the Calliphoridae, Sarcophagidae and Silphidae families, and only those species of Staphylinidae of known predatory habits were included in the analyses.
Diptera were identified by M. A. Marcos–García (Universidad de Alicante, Spain), and Coleoptera were identified by J. Márquez (Universidad Autónoma del Estado de Hidalgo, Mexico). Voucher specimens were deposited in the Coleoptera Collection of the Universidad Autónoma del Estado de Hidalgo, Mexico.
The completeness of the species inventories for each bait was assessed as the proportion of observed species richness relative to maximum expected richness. Expected richness was calculated using a non–parametric richness estimator, Chao2, which is based on incidence data (presence–absence) using EstimateS version 8.0.0 (Colwell, 2006). To determine whether the abundance of the different species of necrophilous insects attracted varied among baits, a Kruskal–Wallis test with a chi–square approximation was performed (SAS Institute, 1989).
The diversity of the necrophilous communities collected with each type of bait was compared using Shannon’s index (Magurran, 2004) and its 95% confidence interval computed by bootstrap methods for a direct statistical comparisons among bait types using the StatsDirect v.2.5.7 software (StatsDirect Ltd, 2005). Abundance patterns and species evenness among baits were compared graphically with rank–abundance plots (Feinsinger, 2001).
In order to determine whether differences exist between the baits in terms of their attractiveness for necrophagous and predatory insects, a step–by–step discriminant function analysis was used (Manley, 1994). The abundance of necrophagous and predatory species was analyzed in order to determine whether differences between guilds were manifested between baits.
Five species of Coleoptera and 8 of Diptera were recorded (Table 1). None of these species were recorded in the control traps. The percentage of completeness of species inventories for each bait, according to the expected richness predicted with Chao2 estimator, was 98% for squid, 100% for chicken, 97% for mouse, and 87% for lizard. Squid attracted 43% of the total specimens from 12 species, chicken attracted 36% from 12 species, mouse attracted 13% from 9 species, and lizard attracted 8% from 10 species (Table 1).
The abundance of the predator Philonthus sp. (Staphylinidae), and the necrophagous Calliphora sp. 1, Calliphora sp. 2, Phaenicia sp., and Lucilia sp. (Calliphoridae) differed significantly among baits (p < 0.05) (Table 1). The reason for these differences are currently not known; however, other studies have shown that necrophagous species such as the calliphorids make differential use of carcasses, due to preference or attraction of adult flies for carcass type, size, grade of decomposition, and the relative abundance of a particular type of carcass (Ives, 1991).
The high abundance of Philonthus sp. in the squid and chicken baited traps stands out because predators do not necessarily need certain substrates when searching for prey. As we lack information concerning other substrates where this species may forage, for example excrement or decomposing fungi, it can only be suggested that this species is attracted to the odors that are more easily detected.
Significant differences in diversity (Shannon index) between baits were found. The estimated diversity index for the squid baited traps (H= 1.56) differed significantly from those estimated for lizard (H= 2.07), chicken (H= 1.94), and all baits pooled (H= 1.86), but did not differ from mouse baited traps (H= 1.59). The indexes estimated for lizard, chicken, mouse, and all baits pooled did not differed significantly. The low diversity index for squid was due to the dominance of Philonthus sp., since a decrease in the magnitude of Shannon diversity index occurs when dominant single species are considered (Magurran, 2004). The relative abundances of the different species also varied between baits, and Philonthus sp. was the dominant species in all baits employed, except in lizard–baited traps, where P. hoegei was dominant (Fig. 1).
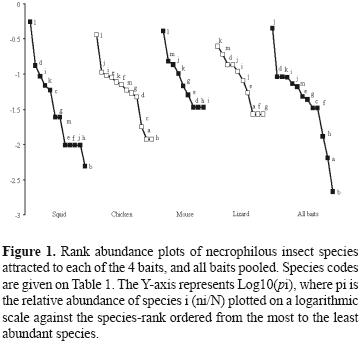
The results of the discriminant function analysis revealed that the abundance of predatory insects was significantly lower than that of necrophagous insects among baits (λ= 0.093, F= 19.57; d.f.= 4, 8, p < 0.0003). The analysis required only a single axis in order to detect differences between the 2 guilds, where the relevant variables were mouse and lizard baits. These results suggest that necrophagous insects differ from predators in their sensitivity or preference to odors produced by different carcasses. This aspect deserves further study since there appears to be no study comparing sensitivity and preference of necrophagous insects and their associated predators.
The fact that none of the individual baits was able to attract all species and that their abundances varied among baits, suggests that the employment of different baits is important when studies are addressing questions about community structure, and to obtain a more uniform unit for measuring collection effort (Longino et al., 2002).
We sincerely thank Roger Guevara, Juan Márquez, Fernando Vaz de Mello, and Iriana Zuria for reviewing and offering suggestions to improve the manuscript and Claudia E. Moreno for help with the diversity analyses. We would like to thank the economic support provided by the project FOMIX Hidalgo 43761 and 95828. I. Castellanos thanks PROMEP for supporting this work.
Literature cited
Carter, D. O., D. Yellowlees and M. Tibbett. 2007. Cadaver decomposition in terrestrial ecosystems. Naturvissenschaften 94:12–24. [ Links ]
CNA. 2006. Oficina de Cálculo Climático. Red Hidroclimatológica Estatal de Hidalgo, Hidalgo, México. 68 p. [ Links ]
Colwell, R. K. 2006. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 8.0. Department of Ecology and Evolutionary Biology, University of Connecticut, Storrs, Connecticut. http://viceroy.eeb.uconn.edu/estimates; last consulted: 23.VII.2010. [ Links ]
Feinsinger, P. 2001. Designing field studies for biodiversity conservation. The Nature Conservancy, Island Press, Washington, D.C. 212 p. [ Links ]
Gibbs, J. P. and E. J. Stanton. 2001. Habitat fragmentation and arthropod community change: carrion beetles, phoretic mites, and flies. Ecological Applications 11:79–85. [ Links ]
Ives, A. R. 1991. Aggregation and coexistence in a carrion fly community. Ecological Monographs 61:75–94. [ Links ]
Larsen, T. H. and A. Forsyth. 2005. Trap spacing and transect design for dung beetle biodiversity studies. Biotropica 37:322–325. [ Links ]
Longino, J. T., J. Coddington and R. K. Colwell. 2002. The ant fauna of tropical rain forest: estimating species richness three different ways. Ecology 83:689–702. [ Links ]
Manley, B. F. J. 1994. Multivariate Statistical Methods: a Primer, 2nd edn. Chapman and Hall/CRC, Boca Raton, Florida. 232 p. [ Links ]
SAS Institute. 1989. SAS/STAT User´s Guide, version 6, 4th edn. SAS Institute, Cary, North Carolina. 1674 p. [ Links ]
StatsDirect Ltd. 2005. StatsDirect statistical software. England, StatsDirect Ltd. http//www.statsdirect.com; last consulted: 23.VII.2010. [ Links ]
Ulrich, W., K. Komoznski and M. Zalevski. 2008. Body size and biomass distributions of carrion visiting beetles: do cities host smaller species? Ecological Research 23:241–248. [ Links ]

