










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de biodiversidad
versión On-line ISSN 2007-8706versión impresa ISSN 1870-3453
Rev. Mex. Biodiv. vol.82 no.1 México mar. 2011
Nota científica
Novedades sobre el polen de Acacia mayana, especie poco conocida (Leguminosae: Mimosoideae: Acacieae)
Novelties on the pollen of Acacia mayana, a little known species (Leguminosae: Mimosoideae: Acacieae)
M. de Lourdes Rico–Arce
HLAA, Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3AB, UK.
Correspondencia:
l.rico@kew.org
Recibido: 16 octubre 2009
Aceptado: 09 abril 2010
Resumen
Se presenta la descripción del polen de Acacia mayana Lundell, se discuten las similitudes de éste con el de otras especies de la subfamilia Mimosoideae, incluyendo aquellas del paleotrópico. Asimismo, se proporciona una descripción de la especie y un mapa de distribución de la misma en el neotrópico.
Palabras clave: Acacia, polen, políada, Leguminosae, Mimosoideae.
Abstract
A description of Acacia mayana Lundell polyad is presented. It is compared with the morphology of other polyads of species within the subfamily Mimosoideae, including some from the paleotropics. A morphological description of the species and a distribution map are provided.
Key words: Acacia, pollen, polyad, Leguminosae, Mimosoideae.
Acacia mayana Lundell es una de las 15 especies de Acacia mirmecófilas neotropicales (Rico Arce, 2007). El grupo es fácil de distinguir dentro de Acacia subg. Acacia: las plantas se caracterizan por tener espinas grandes modificadas en domacios, comúnmente habitados por una o varias especies de hormigas del género Pseudomyrmex. Con respecto a su morfología, las especies mirmecófilas poseen una sinapomorfia, que consiste en que la forma de la bráctea floral es peltada, a diferencia de otras especies en las que tiende a ser espatulada o claviforme.En la mayoría de las especies mirmecófilas, el fruto es indehiscente, las semillas se encuentran rodeadas por un arilo generalmente amarillo y las inflorescencias son espigas o capítulos. En A. mayana son espigas organizadas en un eje central, lo que difiere del resto de las especies del grupo (Fig. 1). Los colores y morfología de esta inflorescencia dan la impresión de una posible polinización por aves, aunque hasta la fecha no se tiene evidencia de esto.

Fuera del neotrópico se distribuyen 4 especies de Acacia mirmecófilas: A. bussei Hams ex Sjöstedt A. drepanolobium Hams ex Sjöstedt, A. formicarum Hams y A. seyal Del., pero en éstas la mirmecofilia es facultativa, es decir, las plantas viven exitosamente con o sin la presencia de hormigas; así mismo, las espinas pueden formar domacios o no, y el fruto y brácteas florales de estas plantas mirmecófilas son como en las especies no mirmecófilas.
Otra especialización única (autoapomorfia) de las especies mirmecófilas neotropicales son los cuerpos de Belt, ubicados en los ápices de los folíolos, como en A. gentlei Standl. (Fig. 2). Estas estructuras, que son fuente de carbohidratos para las hormigas, no las tienen en ninguna etapa de su vida las especies africanas analizadas.

Estudios moleculares recientes (Gómez–Acevedo et al., 2010) demuestran que el grupo mirmecófilo americano es monofilético, teniendo como grupo hermano especies de Acacia subg. Acacia, p. ej., Acacia macracantha Humb. et Bonpl. ex Willd.; a diferencia de las 2 especies mirmecófilas africanas, no se resuelven como monofiléticas per se, pero sí se encuentran dentro del clado que contiene otras especies del género Acacia subg. Acacia en África, que no son mirmecófilas.
La palinología de numerosas especies del género ha sido estudiada por Van Campo y Guinet (1961), Guinet (1969, 1986), Guinet y Lugardon (1976), Kordofani e Ingrouille (1992), Caccavari y Dome (2000a, 2000b), y Rico y Banks (2001); aunque para las especies mirmecófilas en particular, sólo 3 han sido documentadas por Guinet (1969).
Se elaboró una micrografía electrónica del polen no acetolizado de Acacia mayana. Las muestras de polen se obtuvieron de los botones de flores en ejemplares de herbario depositados en K y MEXU. Se colocaron en una solución humectante, libsorb al 1 %; posteriormente se transfirieron a etanol 95 % y se montaron en el portamuestras. Una vez que el material estuvo seco se bañó con platino. La observación del polen y toma de micrografías fueron hechas en un microscopio electrónico de barrido Hitachi S4700 a 2kv. Para la descripción del polen se usó la terminología de Punt et al. (1994) y Hesse et al. (2009).
Se revisó la colección de laminillas palinológicas para microscopio de luz, de las tribus Acacieae e Ingeae depositadas en la colección Kew–Jodrell de los Royal Botanic Gardens, Kew. Asimismo, se revisó la literatura correspondiente a la palinología de estas tribus.
Con el objeto de elaborar la descripción morfológica y el mapa de distribución de la especie, se revisaron los ejemplares de herbario depositados en BM, K y MEXU (Index herbariorum <http://sweetgum.nybg.org/ih/>).
Material examinado. GUATEMALA: Dept. Petén, M. Aguilar 495 (MICH). MEXICO: Chiapas; D.E. Breedlove 34245, 34505 (MEXU); G. Domínguez Vázquez 629 (MEXU); S. Levy 2 (MEXU); E.M. Martínez S. 6086, 15075 (MEXU) 26360 (K, MEXU); F. Miranda 6634 (MEXU); J. Quintanilla 1014 (MEXU); G. Shapiro 445 (K, MEXU); S. Sinaca 1111 (MEXU); M. Sousa 12386 (BM, MEXU). Oaxaca; M. Sousa 7286, 9213 (MEXU).Tabasco; E. Matuda, 3410 (K, MEXU); Rovirosa 807 (K); M. Sousa 12789 (MEXU); O. Téllez 7505 (MEXU). Veracruz; S. Flores 84 (MEXU); G. Ibarra M. 1423, 1449, 1546, 1624, 2007, 2639, 2641 (MEXU); M. Sousa 3088 (MEXU); T. Wendt 4049, 5276 (MEXU).
Descripciones
Polen de Acacia mayana Lundell. Políadas de 16–granos, acalimadas. De simetría bilateral y circular en un plano, formada por 2 capas, cada una con 4 granos centrales, éstos rodeados por 8 granos, con engrosamientos, de tal manera que forman una políada discoide convexa. Las políadas miden 47–62 µm de ancho por 22–28 µm de grosor. Las aperturas están en las posiciones ecuatoriales, en la esquina de unión de cada 3 o 4 granos de polen. Pseudocolpos circulares o algo cuadrangulares en la porción distal. La ornamentación en las porciones distales es rugular–perforada y en las superficies de los pseudocolpos granular (Fig. 3A, B). Material tomado de E.M. Martínez 26360 (K, MEXU).
Acacia mayana Lundell
Carnegie Inst. Washington 478: 210. 1937. Holotipo: Guatemala, M. Aguilar 495 (MICH), isotipos (GH!, NY!, US!).Sinonimia: Vachellia mayana (Lundell) Seigler et Ebinger, Phytologia 87(3):163. 2005 [2006].
Nombre común: Acamte, acante, crucetillo.
Árbol o arbusto de 3–10 m de alto, 15 cm de diámetro, tallos ligeramente hirsutos con numerosas lenticelas. Estípulas espiniformes, 5.5×10(–16) mm. Hojas de 15–38 cm de largo, pecíolo de 2–2.8 cm de largo, glabro, con 1 glándula navicular cercana al primer par de pinnas; raquis de 12–30 cm de largo, glabro, con 1 glándula entre cada par de pinnas; pinnas 7–9(–13) pares, de 6.5–10(–14) cm de largo; folíolos de 28–44 pares por pinna, linear–oblongos, de 11–14(–17)×2–3.5 mm, base truncada, inserción subcentral, ápice redondeado, glabros, venación palmado reticulada, conspicua en ambas superficies. Espigas solitarias en un eje central de hasta 11.5 cm de largo, glabro, espigas de 3–4.5 cm de largo, pedúnculos de 1–1.5 cm de largo, glabros, con un involucro en la base formado de brácteas de 1.5–2 mm de largo, membranáceas, estrigulosas, cada pedúnculo con 1 bráctea linear de 8 mm de largo, portando 1 glándula apical estipitada hasta de 2 mm; bráctea floral de 0.6–1 mm de largo, peltada, persistente, estrigulosa, flores blanquecino–café a rojizas; cáliz de 1.2 mm de largo, infundibuliforme, 5(3+2)–lobado, casi truncado, los lóbulos se unen para formar una estructura algo gibosa, glabros; corola de 1.4 mm de largo, infundibuliforme, ligeramente 5–lobada en menos de ¼ de su longitud, ápice glabro; estambres eglandulares; ovario de 0.6–1 mm de largo, glabro, sésil, sin nectario intraestaminal. Fruto 8–12 cm de largo, 1.8–2.2 cm de diámetro, cilíndrico, indehiscente, cartáceo, glabro, ligeramente curvo, rojizo, con base aguda, con estípite de hasta 1.5 cm de largo, con ápice agudo, con 1 apículo de hasta 1.5 cm. de largo. Semillas castaño claro, de forma obovoide, de 10–11×6–7.5×3.5–4.5 mm, con pleurograma de un 80 %, abierto hacia el área del micrópilo, rodeadas por un arilo amarillo, esponjoso.
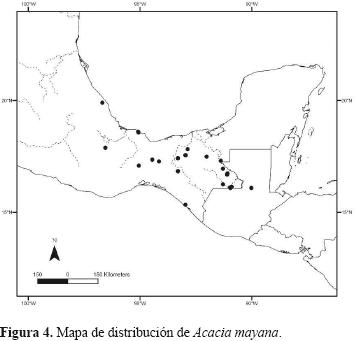
Crece en los bosques tropicales perennifolios. La especie se ha encontrado en floración de marzo a mayo y en noviembre; en fructificación, de marzo a abril; se distribuye en México en los estados de Chiapas, Oaxaca, Tabasco y Veracruz y en Centroamérica en Guatemala (Fig. 4), confirmando aquí que la especie no existe en Belice como se habia mencionado en Rico Arce (2007).
La preparación no acetolizada de Acacia mayana mostró políadas similares a las del género Zapoteca en la tribu Ingeae (Guinet y Hernández, 1989), Zapoteca fue segregado de una sección del género Calliandra (Hernández, 1986). Las características palinológicas que presenta Zapoteca son engrosamientos circulares en los granos centrales de la políada, como se observan en Guinet y Hernández (1989: 9). Este tipo de engrosamiento fue reportado previamente por Niezgoda et al. (1983), quienes propusieron que este grupo podría ser reconocido como un nuevo género; sin embargo, no fue publicado como tal. El género Zapoteca fue oficialmente formalizado por Hernández (1986), quien tomó en cuenta otros caracteres adicionales a su palinología (Guinet y Hernández, 1989). Los engrosamientos circulares característicos de las políadas de Zapoteca (Fig. 3c) se encuentran presentes en Acacia mayana. Por otro lado, Caccavari (2002, y com. pers.) sugiere que las especies de Acacia son más comparables con el género Piptadenia (tribu Mimoseae) compartiendo algunas características, como las políadas homomorfas en la estructura de la exina. Para confirmar esto en A. mayana, es necesario tener cortes e imágenes al microscopio de luz y de transmisión y así poder describir la estructura de la misma, especialmente la presencia o no de columelas. Por otro lado, la forma discoide convexa de la políada de A. mayana la hace más comparable con el género Zapoteca, aunque cabe mencionar que éste es un carácter muy variable entre los géneros de la subfamilia Mimosoideae.
Caccavari (2002) también señala que en las Ingeae es común que las políadas superen los 40 µm, siendo que en general en Acacia y Piptadenia son pequeñas. Las políadas de Acacia mayana miden 47–62 ×22–28 µm, un carácter más que la hace comparable con el género Zapoteca, cuyas políadas miden 60–78 ×28–32 µm.
El estudio de las preparaciones palinológicas depositadas en la colección Kew–Jodrell, mostró que el polen perteneciente a otros géneros neotropicales también presenta estos engrosamientos circulares, muy evidentes en Marmaroxylon y menos en Cojoba, mencionados para ambos por Guinet y Rico (1988). Por otro lado, durante la revisión de las preparaciones en el Laboratorio Jodrell también se observaron en el género Archidendropsis del paleotrópico. Nielsen et al. (1983:336) informan que Archidendropsis presenta fósulas leves en los granos de polen centrales: "faint fossules in central cells"; Palinológicamente, estas fósulas corresponden a los engrosamientos que aquí se comparan con los de Acacia mayana y que, en este caso, parecen presentar una capa más gruesa de polenkite.
Guinet (1969) agrupó dentro del género Acacia subg. Acacia, tanto especies mirmecófilas (A. cornigera [L.] Willd.) como no mirmecófilas (A. constricta Benth., A. pennatula [Schltdl. et Cham.] Benth., A. curvifructa Burkart y A. macracantha), porque éstas presentan políadas acalimadas de 16 granos de polen sincolpados, téctum perforado, poro pequeño de 1.3–2.3 µm de diámetro situado en los ángulos de las caras distales de los granos; columela pequeña de 0.3–0.7 µm; granos distales de 20–44 µm y granos centrales de 9–13 µm. Otras preparaciones hechas por Guinet (com. pers.) corresponden a las especies mirmecófilas de A. chiapensis Saff., A. globulifera Saff. y A. gentlei, las cuales presentan políadas estructuralmente similares a las de A. cornigera.
Es importante señalar que los engrosamientos circulares en los granos de polen en Acacia, subgéneros Acacia y Aculeiferum no han sido documentados previamente. Esta característica palinológica es la primera vez que se registra para las políadas de A. mayana y para Acacia subg. Acacia; mostrando, consecuentemente, más similitud con aquellos géneros de la tribu Ingeae que con los de Mimoseae. Es interesante mencionar que Guinet (1969) describió e ilustró varias políadas de especies de Acacia subg. Phyllodineae, en las que A. farinosa Lindl. muestra también los tipos de engrosamientos circulares como los presentes en A. mayana.
Cowan y Maslin (1995) mencionan que la morfología de la inflorescencia de A. farinosa es notable y no común, ya que presenta en el pedúnculo un involucro formado de brácteas unidas en sus márgenes; este involucro es generalmente una característica única en Acacia subg. Acacia, pero no para el subgénero Phyllodineae, predominantemente nativo de Australia, que posee 970 especies, con sólo otras 9 en islas del Pacífico. Cowan y Maslin (1995) aclaran que, aunque A. farinosa presenta este involucro, no se puede sugerir la hipótesis de que ésta sea una estructura homóloga a la que se encuentra en las especies de Acacia subg. Acacia.
La autora agradece al profesor Philippe Guinet, a la Dra. Caccavari (†) y al Dr. G. Lewis, por sus valiosos comentarios al manuscrito inicial de este artículo; a la Dra. H. Banks, por las magníficas preparaciones de polen; a Christine Niezgoda, por ceder el permiso de usar la imagen de Zapoteca formosa (Calliandra capillata); a Steve Bachman, por la elaboración del mapa de distribución; al Dr. Fernando Chiang, por sus atinadas correcciones y sugerencias, y a los revisores anónimos que con sus sugerencias y cuidadoso trabajo mejoraron esta contribución.
Literatura citada
Caccavari, M. A. 2002. Pollen morphology and structure of tropical and subtropical American genera of the Piptadeniagroup (Leguminosae: Mimosoideae). Grana 41:130–141. [ Links ]
Caccavari, M. A. y E. A. Dome. 2000a. Subpseudocolpi in polyads of Acacia, subgenus Aculeiferum. Grana 39:32–38. [ Links ]
Caccavari, M. A. y E. A. Dome. 2000b. An account of morphological and structural characterization of American Mimosoideae pollen. Part I. Tribe Acacieae. Palynology 24:231–248. [ Links ]
Cowan, R. S. y B. R. Maslin. 1995. Acacia Miscellany 11. Miscellaneous taxa of northern and eastern Australia of Acacia section Plurinerves (Leguminosae: Mimosoideae). Nuytsia 10:63–84. [ Links ]
Gómez Acevedo, S. L., M. de L. Rico Arce, A. O. Delgado Salinas, S. Magallón y L. E. Eguiarte. 2010. Neotropical mutualism between Acacia and P s e u d o m y r m e×: phylogeny and divergence times. Molecular Phylogenetics and Evolution 56:393–408. [ Links ]
Guinet, Ph. 1969. Les Mimosacées. Etude de palynologie fundamentale, corrélations, évolution. Travaux de la Section Scientifique et Technique, Institut Français Pondichéry 9:1–293. [ Links ]
Guinet, Ph. 1986. Geographic patterns of the main pollen characters in the genus Acacia (Leguminosae), with particular reference to subgenus Phyllodineae. In Pollen et spores: form and function, Linnean Society Symposium series, 12, S. Blackmore y I. K. Ferguson (eds.) Academic, London. p. 297–311. [ Links ]
Guinet, Ph. y B. Lugardon. 1976. Diversité des structures de l'exine dans le genre Acacia (Mimosaceae). Pollen et Spores 18:483–511. [ Links ]
Guinet, Ph. y L. Rico Arce. 1988. Pollen characters in the genera Zygia, Marmaroxylon and Cojoba (Leguminosae, Mimosoideae, Ingeae): a comparison with related genera. Pollen et Spores 30:313–328. [ Links ]
Hernández, H. M. 1986. Zapoteca: a new genus of neotropical Mimosoideae. Annals of the Missouri Botanical Garden 73:755–763. [ Links ]
Hesse, M., H. Halbritter, R. Zetter, M. Weber, R. Buchner, A. Frosch–Radivo y S. Ulrich. 2009. Pollen terminology: an illustrated handbook. Berlin: Springer Verlag. 261 p. [ Links ]
Index Herbarioum. New York Botanical Garden. Holmgren, P. K. y N. H. Holmgren (eds.). http://sweetgum.nybg.org/ih/; última consulta: 01.X.2009. [ Links ]
Kordofani, M. y M. I. Ingrouille. 1992. Geographical variation in the pollen of Acacia (Mimosaceae) in Sudan. Grana 31:113–118. [ Links ]
Nielsen, I., Ph. Guinet y T. Baretta–Kuipers. 1983. Studies in the Malesian, Australian and Pacific Ingeae (Leguminosae–Mimosoideae): the genera Archidendropsis, Wallaceodendron, Paraserianthes, Pararchidendron and Serianthes. Bulletin du Muséum National d'Histoire Naturelle, 4e série, section B. Adansonia 5:335–360. [ Links ]
Niezgoda, C. J., S. M. Feuer y L. I. Nevling. 1983. Pollen ultrastructure of the tribe Ingeae (Mimosoideae: Leguminosae). American Journal of Botany 70:650–667. [ Links ]
Punt, W., S. Blackmore, S. Nilsson y A. Le Thomas. 1994. Glossary of pollen and spore terminology. LPP Contributions series No. 1.Laboratory of Palaeobotany and Palynology, University of Utrecht. 71 p. [ Links ]
Rico Arce, L. y H. Banks. 2001. A preliminary survey of pollen and other morphological characters in neotropical Acacia subgenus Aculeiferum (Leguminosae: Mimosoideae). Botanical Journal of the Linnean Society 135:263–270. [ Links ]
Rico Arce, M. de L. 2007. A checklist and synopsis of American species of Acacia (Leguminosae: Mimosoideae). Comisión Nacional para el Conocimiemnto y Uso de la Biodiversidad/ Redacta, México, D.F. 207. p. [ Links ]
van Campo, M. y Ph. Guinet. 1961. Les pollen composés, l'example des Mimosacées. Pollen et Spores 3:201–218. [ Links ]

