










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de biodiversidad
versão On-line ISSN 2007-8706versão impressa ISSN 1870-3453
Rev. Mex. Biodiv. vol.82 no.1 México Mar. 2011
Ecología
Abundancia de dos poblaciones de toninas (Tursiops truncatus) en el norte de Veracruz, México
Abundance of two populations of bottlenose dolphins (Tursiops truncatus) in northern Veracruz, Mexico
Michelle P. Valdes–Arellanes1, Arturo Serrano1*, Gisela Heckel2, Yolanda Schramm3 e Ibiza Martínez–Serrano1
1 Laboratorio de Mamíferos Marinos, Facultad de Ciencias Biológicas y Agropecuarias, Universidad Veracruzana. Km. 7.5 Carretera Tuxpan–Tampico Núm. 70 , 92850 Tuxpan, Veracruz, México.
2 Departamento de Biología de la Conservación, Centro de Investigación Científica y de Educación Superior de Ensenada, 22860 Ensenada, Baja California, México.
3 Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, 22800 Ensenada, Baja California, México.
*Correspondencia:
arserrano@uv.mx yschramm@uabc.mx
Recibido: 05 enero 2010
Aceptado: 28 mayo 2010
Resumen
En México no se conoce el tamaño de las poblaciones de la tonina (Tursiops truncatus) debido a su gran movilidad y al ocasional intercambio de individuos entre poblaciones. El objetivo de este estudio fue determinar la abundancia de toninas en la zona norte de Veracruz. De marzo de 2005 a diciembre de 2007 se realizaron 50 navegaciones con un esfuerzo total de 236.7 horas. Los resultados obtenidos de la foto–identificación de toninas, se emplearon en el modelo Jolly–Seber y se estimó con este modelo un total de N=302 ±113 organismos para toda el área de estudio. Para Tamiahua, se estimó una población de N=177 ± 10 delfines y para Tuxpan N=161 ± 68 delfines. Estos resultados sugieren que existe un mínimo intercambio de delfines entre zonas y por tanto, las poblaciones no están aisladas; sin embargo, los organismos tienen preferencia por ciertas áreas. La foto–recaptura de algunos individuos en la zona de Tamiahua confirma que la fidelidad al sitio se ha mantenido por 13 años. Es probable que esto se deba a que cada una de las zonas presenta estuarios con alta productividad que son ideales para refugio y crianza de las toninas.
Palabras clave: foto–identificación, golfo de México, Jolly–Seber, esfuerzo, fidelidad al sitio, intercambio de individuos.
Abstract
In Mexico, the population size of the bottlenose dolphin (Tursiops truncatus) is unknown, due to the fact that they are highly mobile, and occasionally there are exchanges of individuals between populations. The objective of this study was to determine the abundance of dolphins in the north of Veracruz. From March 2005 to December 2007 were 50 surveys with a total effort of 236.7 hours. The results of the photo–identification of dolphins were used in the Jolly–Seber model, and with this model we estimated a total of N = 302 ± 113 dolphins throughout the study area. For Tamiahua a population estimate of N = 177 ± 10 dolphins, and for Tuxpan N = 161 ± 68 dolphins. These results suggest that there is minimal exchange of dolphins between areas; therefore it is assumed that populations are not isolated but dolphins have a preference for certain areas, and confirms that there is fidelity to the site of 13 years from the photo–recapture of some individuals in the area of Tamiahua. The fidelity may be a consequence of the high primary productivity present in the area, therefore is an ideal place for breeding and shelter.
Key words: photo identification, Gulf of Mexico, Jolly–Seber, effort, site fidelity, exchange.
Introducción
En el golfo de México, especialmente en la zona norte del estado de Veracruz, la tonina (Tursiops truncatus) ha comenzado a emplearse como bioindicador para el monitoreo del estado de salud del mar (Serrano et al., 2007), ya que en esta zona se realizan actividades pesqueras y petroleras de gran impacto económico para el estado. Por lo tanto, es importante conocer el tamaño de las poblaciones de toninas en la zona.
Tursiops truncatus es una especie con distribución geográfica amplia, por lo que sus patrones de movimiento pueden variar a lo largo de un año con migraciones estacionales (Shane et al., 1986) que influyen de manera temporal en el tamaño de sus poblaciones. Las técnicas de marcaje–recaptura que comúnmente se usan para estudios de poblaciones permiten determinar su tamaño y estimar otros parámetros, como supervivencia y reclutamiento, que pueden relacionarse con aspectos de presión ambiental, competencia de recursos y depredación (Hammond et al., 1990).
Uno de los parámetros para medir las poblaciones es la abundancia. La estimación de la abundancia se realiza principalmente mediante 2 métodos: muestreo de distancias por transecto lineal y captura–recaptura (Buckland et al., 2001). Con el primero sólo se estima la abundancia y densidad en un tiempo específico, en tanto que el segundo permite la estimación de otros parámetros poblacionales de manera no invasiva (Seber, 1982; Hammond, 1990; Hammond et al ., 1990; Pollock et al., 1990). Algunos de estos parámetros son la densidad poblacional, la madurez sexual, la fidelidad al sitio, la estructura de grupo e incluso la organización social (Wells et al., 1990, Lusseau, 2005, Merriman, 2007).Una herramienta útil en la estimación de la abundancia de una población para ambos métodos y fundamental para el método captura–recaptura, es la foto–identificación (Weller, 1991), la cual se utiliza para individualizar organismos de acuerdo con marcas permanentes en sus aletas dorsales (Defran et al., 1990) y provee un registro permanente que puede ser documentado y verificado. El estado de conservación de la mayor parte de las poblaciones de esta especie es desconocida, por lo que en la Lista roja de la IUCN se ubica en la categoría "estado de riesgo desconocido"; en CITES se encuentra dentro del Apéndice II, y en la Norma Oficial Mexicana NOM–ECOL–059–2001 se considera como "sujeta a protección especial". Lo anterior se debe a la ausencia de datos y estudios estandarizados de las poblaciones existentes.
En la zona del golfo de México se han realizado muy pocos estudios de mamíferos marinos (Ortega–Ortiz et al., 2004), en especial de las toninas, que son un depredador potencial de gran importancia como último eslabón de las cadenas tróficas (Kelly, 1983; Leatherwood y Reeves,1990), por lo que en la zona norte del estado de Veracruz ha comenzado a utilizarse T. truncatus como bioindicador del estado de salud del ecosistema marino, ya que en esta zona se realizan actividades pesqueras y petroleras de gran impacto económico para la región (Serrano et al., 2007); por lo tanto, entre otros parámetros poblacionales de esta especie, es importante conocer su abundancia.
Materiales y métodos
El área de estudio está situada en el golfo de México, en la zona norte del estado de Veracruz, entre las coordenadas 21° 29' N, 97° 08' O y 20° 09' N, 96° 39' O, y abarca un área aproximada de 9 000 km2 (Fig. 1), se caracteriza por la presencia de una laguna y 2 ríos y se ha observado que estas desembocaduras de agua dulce proveen oportunidades de alimentación a T. truncatus; en la zona se hallan arrecifes coralinos que pueden brindar refugio a algunas especies, tanto de peces como de cetáceos (Barros y Wells, 1998; Connor et al., 2000, Baumgartner et al., 2001) y existen registros previos de toninas (Heckel, 1992; Schramm, 1993); por tanto, el área fue dividida en 2 zonas de estudio:
1. Tamiahua. Desde punta Cabo Rojo al norte (21°35'N, 97°19'O; 21°34'N, 97°06'O) hasta la comunidad de San Antonio al sur, donde limita con la zona de Tuxpan (21°03'N, 97°20'O; 21°13'N, 96°56'O). El área de estudio está caracterizada por la presencia de arrecifes coralinos y la desembocadura de la laguna de Tamiahua.
2. Tuxpan. Limita al norte con la zona de Tamiahua (21°03'N, 97°20'O; 21°13'N, 96°56'O), y al sur con las coordenadas 20°34'N, 97°06'O; 20°43'N, 96°40' O. En esta zona se encuentran las desembocaduras de los ríos Tuxpan y Cazones.

Registro de datos durante navegaciones. Se llevaron a cabo navegaciones semanales durante el periodo comprendido entre marzo del 2005 y diciembre del 2007. Se utilizó una embarcación para pesca deportiva de 9 m de eslora con 2 motores integrados de gasolina, marca Chevrolet, de 275hp. Se efectuaron transectos al azar, no sistemáticos, tratando de cubrir el área de estudio en su totalidad. Las navegaciones se hicieron a una velocidad aproximada de 18 km/hr, con una duración promedio de 5 horas, dependiendo de las condiciones meteorológicas, procurando navegar con el estado del mar ≤3 en escala de Beaufort. Para cada avistamiento así como para las navegaciones se llenaron fichas previamente diseñadas donde se registraron fecha, hora de salida, hora de llegada, nombre del observador, posición geográfica (usando un GPS Garmin Etrex, precisión ± 3 metros), hora de inicio y final del avistamiento, número de organismos observados, y composición grupal de acuerdo con John et al. (2000).
Foto–identificación. Una vez avistado un grupo de toninas, se les tomaron fotografías de ambos costados del cuerpo, utilizando una cámara digital profesional Cannon EOS Rebel y un lente zoom de 100–400 mm. La lancha permaneció con el grupo hasta que se consideró haber fotografiado la totalidad de los individuos. La técnica de foto–identificación consiste en diferenciar los organismos por medio de características presentes en las aletas dorsales, tales como su coloración, así como la presencia de muescas, marcas,y cicatrices (Mazzoil et al., 2004, Würsing y Jefferson, 1990). El proceso de foto–identificación comienza con la revisión de las carpetas de cada avistamiento usando el programa Adobe Photoshop 7.0. Estas imágenes se revisan de manera seriada para identificar las fotografías de mejor calidad, las cuales son seleccionadas para su posterior comparación con el catálogo existente. Todas las imágenes en la carpeta se guardan, anotando fecha y zona de avistamiento y un número de serie de 3 dígitos. Con el fin de reducir los errores por criterios unilaterales y/o ambiguos, el proceso de comparación lo realizan 3 personas capacitadas.
Para conocer si se ha fotografiado la totalidad de organismos en la población se adaptó el modelo de Darling–Morowitz (1986) que consiste en la construcción de una gráfica que representa la tasa de aparición de nuevos individuos identificados, en donde "x" es igual al número acumulativo de toninas individualizadas, y "y" es igual al número acumulativo de delfines foto identificados. Cuando el número de animales diferenciados se incrementa de la misma manera que el número de identificaciones, la gráfica presentará una pendiente de 1, lo que indica que siguen apareciendo animales nuevos en la población; cuando la pendiente de la curva comienza acercarse a la asíntota en el valor de "y" es porque ya no se están registrando animales nuevos en la población; por lo tanto, el valor de "y" será igual al tamaño de la población.
Eficiencia fotográfica. Una vez fotografiados los organismos se obtuvo el índice de éxito de la foto–identificación, este indicador es útil para determinar la existencia de posibles sesgos en el uso de la técnica de foto–identificación a lo largo del muestreo. Se obtiene dividiendo el número de fotos exitosas (A) entre el número de fotos tomadas (B) y se multiplica por 100.
Posteriormente y con el fin de evaluar la residencia de los organismos, las fotografías del catálogo generado, fueron comparadas con uno elaborado anteriormente por Heckel (1992) y Schramm (1993) para delfines observados en la laguna de Tamiahua.
Abundancia absoluta. Para la estimación del número de organismos en cada población, se utilizó el modelo Jolly Seber para poblaciones abiertas (Ec.1) (Seber, 1982; Pollock et al., 1990), el cual tiene los siguientes supuestos: a) cada animal tiene la misma probabilidad de ser foto–identificado al tiempo de cada muestra; b) las maracas de identificación no cambian ni se pierden durante el periodo de estudio, y c) se presenta emigración entre los individuos. Los individuos son marcados en este caso por medio de la fotografía durante el tiempo de muestreo.
Las fotografías tomadas a cada uno de los organismos sirven como marca durante el periodo de muestreo; así, la primera vez que fue fotografiado un individuo se consideró como la captura, y las siguientes fotografías del mismo organismo tomadas en diferente año se consideraron como recaptura; en este caso, foto–recapturas. Se consideró por años ya que para poblaciones abiertas se da por hecho el que haya adiciones o pérdidas de individuos (nacimientos y/o muertes) y también fue debido a que el índice de foto–recapturas fue bajo.
Para obtener estimaciones del tamaño de poblaciones abiertas es necesario contar con 3 o más recapturas Por lo tanto, la abundancia se estimó mediante la siguiente ecuación (Begon et al., 1996):
Ec.1.

donde:
i = Año (periodo de muestreo)
Ni = Abundancia en el periodo de muestreo i (i = 2,..., k–1)
mi = Número de animales marcados capturados en el periodo de muestreo i
ni = Número de individuos fotografiados en la muestra i (i = 1,...., k);
Ri = Número de individuos liberados de los ni marcados (i = 1,..., k–1) (igual que ni debido a que sólo se fotografía).
Zi = Suma del número de individuos foto–recapturados antes y después de la muestra i, pero no en la muestra i (i = 2,..., k–1);
ri = Suma del número de individuos fotografiados de los liberados en Ri que son foto–recapturados (i=2,..., k–1)
Dentro del modelo Jolly–Seber se efectúa el cálculo de la varianza como a continuación se expresa (Seber, 1982):
Ec.2.
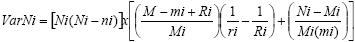
donde:
ai = Porción de individuos capturados de la población en el muestreo i:
ai = mi/ni
Mi = Número de individuos de la población capturados inmediatamente antes del muestreo i:
La siguiente ecuación es la forma general del modelo (Seber, 1982; Pollock et al., 1990):
Ec. 3.

La estimación se aplicó para toda el área de estudio y para cada una de las zonas, considerando cada año como un periodo de muestreo.
Resultados
Se llevaron a cabo 50 navegaciones con un esfuerzo total de 250.8 horas de navegación. El esfuerzo realizado en Tamiahua fue de 92.4 hr y de 158.4 hr en la zona de Tuxpan. Se tomaron un total de 3 190 fotografías, de las cuales se individualizaron 202 organismos en toda el área de estudio. De éstos, 118 pertenecieron a la zona de Tamiahua y 84 a Tuxpan (Fig. 2). La eficiencia fotográfica fue del 56% para toda el área de estudio; 53.4% para la zona de Tamiahua y 58.8% para la de Tuxpan. No presentaron diferencias específicas en la eficiencia fotográfica a pesar de la diferencia de esfuerzo en las zonas.

Abundancia absoluta. De acuerdo con los organismos individualizados, se estimó la abundancia absoluta de las poblaciones con el estimador de Jolly–Seber (Cuadro 1), en el periodo comprendido entre marzo de 2005 a diciembre de 2007, la cual fue de N=302 ±113 en toda el área de estudio. La estimación de la población para Tamiahua fue de N= 177 ± 10 y para Tuxpan fue de N= 161 ± 68.
La mayoría de los avistamientos se registraron muy cerca de la costa, a menos de 10 km (Fig. 3) en las 2 zonas de estudio. Las foto–recapturas fueron ubicadas geográficamente en cada una de las zonas de estudio (Tamiahua y Tuxpan), con la finalidad de corroborar si existe algún intercambio entre las poblaciones. El mes en el que más foto–recapturas se obtuvieron fue diciembre de 2007 para la zona de Tamiahua, en la cual se obtuvieron 25 foto–recapturas de diferentes individuos, y en el mes de junio de 2007 en la zona de Tuxpan se obtuvieron 18. Es importante resaltar que para la estimación de la abundancia se tomaron en cuenta sólo las foto–recapturas de cada zona, sin embargo se registraron 11 casos en los que los individuos se foto–recapturaron tanto en la zona de Tamiahua como de Tuxpan y viceversa, equivalente al 5.4% de la población total (202 toninas).
Fidelidad al sitio. Los resultados de foto–identificación obtenidos fueron comparados con los de Heckel (1992) y Schramm (1993). Se encontraron dos foto–recapturas en la zona de Tamiahua lo cual demuestra que la fidelidad al sitio de estos organismos es por lo menos de 13 años (Fig. 4).
Discusión
Aunque el esfuerzo fue menor en la zona de Tamiahua, ésta fue la zona donde las toninas fueron observadas con mayor frecuencia, y con la estimación de la abundancia de las poblaciones de las 2 zonas de estudio se muestra que la población de Tamiahua es más grande a la de Tuxpan. Las poblaciones estudiadas se observaron principalmente cerca de los ríos presentes en cada zona así como en la desembocadura de la laguna de Tamiahua, lo cual coincide con lo publicado en cuanto a su distribución en lagunas costeras, desembocaduras de ríos, zona litoral nerítica y oceánica del golfo de México (Delgado–Estrella, 1991).
Se reconoce que las toninas prefieren estos lugares debido a que son áreas de crianza y refugio de fases larvales y juveniles de peces y crustáceos, que una vez en fases adultas son presas potenciales para las toninas (Baumgartner et al., 2001; Turner, 1977). Las toninas son organismos oportunistas y presentan una gran flexibilidad de comportamiento (plasticidad), esto es muy evidente y diverso en cuanto a sus técnicas y tipo de alimentación (Reynolds et al., 2000). Por consiguiente, se presume que las toninas estudiadas en este trabajo no necesitan trasladarse mucho en busca de alimento, ya que en ambas zonas pueden encontrarlo en abundancia. Existe poco movimiento de toninas entre las zonas Tamiahua–Tuxpan, y por esta razón son usualmente avistadas en la misma área.
Abundancia de las poblaciones. En el presente trabajo la curva de nuevos delfines foto–identificados (Fig. 2) mostró un incremento constante sin ninguna tendencia hacia la asíntota durante el periodo de estudio. De manera similar, en la costa de San Diego entre 2004 y 2005 se estimó un tamaño de población de 325 individuos (CV=0.13, 95%, CI=259–430). El número mínimo de delfines identificados fue de 164, sin embargo, la curva de descubrimiento de nuevos animales no había llegado todavía a una asíntota en ese estudio (Dudzik et al., 2006). La falta de identificación por marcas naturales en la aleta dorsal fue del 35%; entonces, considerando este punto, la estimación real sería de 400 a 500 individuos (Defran et al., 1999). Tomando en cuenta que algunos de los supuestos del modelo Jolly Sebe no se cumplen en su totalidad, como por ejemplo, que no todos los organismos presentaron marcas naturales distinguibles, lo que llevó a varios excluidos de la comparación. Por lo que los resultados indican, las poblaciones de toninas de la zona norte de Veracruz podrían estar subestimadas. Es probable que el área de estudio represente solamente una porción del ámbito hogareño de los toninas que habitan esta zona y que la densidad de individuos sea mayor a la supuesta en este estudio (Serrano et al., 2007).
La estimación mínima de delfines costeros en la parte estadounidense del golfo de México, desde el noreste del río Grande hasta el río Mississippi,fue de 93 , y en lo que respecta a bahías, lagunas y estuarios, la estimación fue de 3 933 individuos. Los resultados derivaron del uso de transecto lineal como método de estimación (Buckland et al., 2001, Waring et al., 2002). Otro ejemplo de estimación es la que se realizó en la plataforma continental de los Estados Unidos en el golfo de México, desde la frontera mexicana hasta la zona comprendida entre Key West y la Florida, donde se estimó una abundancia de 25 320 (C.V. 0.26) (Fulling et al., 2003). En contraste, los datos obtenidos dentro las 2 áreas de estudio del norte de Veracruz es mucho menor, es probable que la diferencia que se observa en el número esté relacionada con el área geográfica que se estudió en el presente trabajo, más pequeña que la de los trabajos citados anteriormente; sin embargo, es importante mencionar que las poblaciones pueden estar subestimadas, ya que la forma de muestreo empleada fue diferente, menor el número de datos y otro el modelo que se usó.La estimación de poblaciones de toninas en la parte mexicana del golfo de México fue de 58 toninas para el año de 1992 (Heckel, 1992) y 41 para el año de 1993. (Schramm, 1993). Ambas estimaciones fueron realizadas en Tamiahua Veracruz, usando foto–identificación como método. En este último caso se alcanzó la asíntota en N=42, lo que refleja que la población en ese momento dentro de la laguna era relativamente pequeña. Es importante mencionar que en el presente estudio no se registraron organismos dentro de la laguna debido a que presenta muchas charangas (arte de pesca para pequeños crustáceos y peces).en la boca de la laguna, lo que impide la entrada de las toninas; sin embargo, en el trabajo se menciona que la N puede estar , subestimada debido a que algunos supuestos del modelo no se cumplieron en su totalidad ; Tampoco, se cumplió en su totalidad otro supuesto en el presente trabajo: no todos los organismos fueron foto–capturados en el primer avistamiento porque no se abarcó el área completa ni el tiempo lo permitió.
La estimación que se realizó en este trabajo para toda el área de estudio en el centro–norte de Veracruz fue de N=302 ± 113.1, lo que concuerda aproximadamente con lo registrado por otros autores en el golfo de México. Tal es el caso de la laguna de Términos, Campeche, donde se registraron 1 400 individuos (Delgado–Estrella, 2002), así como la laguna de Yalahau y la costa de Holbox, Quintana Roo, con 500 individuos (Delgado–Estrella, 1996). En la costa de Tabasco, la población fluctúa entre 300 y 573 individuos, dependiendo de la temporada (López Hernández, 1997). Actualmente, el catálogo de foto–identificación para la zona centro–norte de Veracruz cuenta con 250 individuos marcados, lo que es congruente con el número obtenido en la estimación realizada y sugiere que las poblaciones tienen un ámbito hogareño relativamente pequeño. Cabe mencionar que en comparación con los estudios de abundancia de las poblaciones, mencionados anteriormente existe una gran similitud en cuanto a las áreas de estudio, ya que cuentan con la presencia de ríos o lagunas costeras, lo que incrementa las posibilidades de observar a los organismos debido a la preferencia que tienen por permanecer en estos sitios.
En este estudio, la abundancia absoluta fue de 302 ± 113.1 organismos en toda el área de estudio, donde 177 ± 10 organismos pertenecen a la población de Tamiahua, y 161 ± 68 organismos a Tuxpan (zonas A y B, respectivamente, Fig. 1). De acuerdo con los trabajos realizados por Bearzi et al. (2008) e Irwin y Würsig (2004), se considera de tamaño medio la población de la zona norte de Veracruz. La estimación de la abundancia a través del modelo Jolly–Seber dio como resultado una varianza media debido a que las recapturas fueron apenas suficientes para cumplir con los supuestos del modelo.
Se pudo constatar que existe poca interacción entre las poblaciones de toninas de Tamiahua y Tuxpan, aunque se presentaron 11 casos de avistamientos de organismos entre las 2 zonas. Los esfuerzos en horas no presentaron diferencias específicas en la eficiencia de foto–identificación a pesar de que para la zona de Tuxpan el esfuerzo fue menor al de Tamiahua. Por lo tanto, se puede suponer que existe un intercambio mínimo de individuos entre sus poblaciones. Comparando los datos de este estudio con los de Scrhamm (1993) y Heckel (1992) se pudo comprobar con foto–recapturas que algunos organismos pertenecientes a la población de Tamiahua muestran una fidelidad al sitio de por lo menos 13 años.
Actualmente se desconoce el estado de las poblaciones de delfines en México, debido a la falta de estudios completos que aporten información demográfica de las poblaciones, a pesar de que esta especie se encuentra clasificada por la IUCN (2008) en estado de riesgo desconocido, en CITES (2008) dentro del Apéndice II, y en la NOM–ECOL–059–2001 (Secretaría de Medio Ambiente y Recursos Naturales, 2002) como sujeta a protección especial . En México, el aprovechamiento de esta especie sólo se permite con fines científicos y es fundamental para su aprovechamiento conocer el estado de las poblaciones de tursiones que garanticen las condiciones necesarias para su manejo y conservación.
Agradecimientos
Este trabajo se realizó gracias al apoyo otorgado por parte de la Cetacean Society International a Arturo Serrano. Agradecemos a la señora Parra, a Aurelio y a Esteban del Club Náutico Aquasports, por el apoyo en las navegaciones, y a Mariel Valdés, por sus enriquecedores comentarios.
Literatura citada
Barros, N. B. y R. S. Wells. 1998. Prey and feeding patterns of resident bottlenose dolphins (Tursiops truncates) in Sarasota Bay, Florida. Journal of Mammalogy 79:1045–1059. [ Links ]
Baumgartner, M. F., K. D. Mullin, L. N. May y T. D. Leming. 2001. Cetacean habitats in the northern Gulf of Mexico. Fishery Bulletin 99:219–239. [ Links ]
Begon, M., J. L. Harper y C. R. Townsend. 1996. Ecology: individuals, populations and communities, tercera edición. Blackwell Science , Oxford . 1068 p. [ Links ]
Berazi, G., S. Agazzi,. S. Bonizzoni, M.,Costa, y A. Azzellino. 2008. Dolphins in a bottle. Abundance, residency patterns and conservation of bottlenose dolphins Tursiops truncatus in the semi–closed eitrophic Amvrakikos Gulf, Greece. Aquatic Conservation: Marine and. Freshwater Ecosystems 18:130–146. [ Links ]
Buckland, S. T. y A. E. York. 2002. Abundance estimation. In Encyclopedia of Marine Mammals, W. F. Perrin, B. Würsig y J. G. M. Thewissen (eds.). Academic, San Diego, California. p. 1–6. [ Links ]
CITES. 2008. The Convention on International Trade in Endangered Species of Wild Fauna and Flora, http://www.cites.org; 11.II.2010 [ Links ]
Connor, R. C., R. S. Wells; J. Mann y A. J. Read. 2000. The bottlenose dolphin. In Cetacean Societies, J. Mann, , R. C. Connor, P. L. Tyack y H. Whitehead (eds.). The University of Chicago Press, Illinois. 433 p. [ Links ]
Darling, J. y H. Morowitz. 1986. Census of "Hawaiian" humpback whales (Megaptera novaeangliae) by individual identification. Canadian Journal of Zoology 64:105–11. [ Links ]
Defran, R. H., G. M. Schultz y D. W. Weller. 1990. A technique for the photo–identification and cataloging of dorsal fins of the bottlenose dolphin (Tursiops truncatus). In Individual recognition of cetaceans: use of photo–identification and other techniques to estimate population parameters, P. S. Hammond, S. A. Mizroch y G. P. Donovan (eds.). Reports of the International Whaling Commission Special issue 12:53–55 p. [ Links ]
Defran, R. H., D. W. Weller, D. L. Kelly y M. A. Espinosa. 1999. Range characteristics of Pacific coast bottlenose dolphins (Tursiops truncatus) in the Southern California Bight. Marine Mammals Science.15:381–393. [ Links ]
Delgado–Estrella, A. 1991 Algunos aspectos de la ecología de poblaciones de las toninas (Tursiops truncatus Montagü, 1821) en la laguna de Términos y sonda de Campeche, México. Tesis, Escuela Nacional de Estudios Profesionales Iztacala, UNAM, México, D.F. 149 p. [ Links ]
Delgado–Estrella, A. 1996. Ecología poblacional de las toninas Tursiops truncatus, en la laguna de Yalahau, Quintana Roo, México. Tesis maestría, Facultad de Ciencias, Universidad Nacional Autónoma de México, México, D.F. 93 p. [ Links ]
Delgado–Estrella, A. 2002. Comparación de parámetros poblacionales de las toninas, Tursiops truncatus, en la región sureste del golfo de México (estados de Tabasco, Campeche, Yucatán y Quintana Roo).Tesis doctorado, Facultad de Ciencias, Universidad Nacional Autónoma de México, México, D.F. 160 p. [ Links ]
Dudzik, K.J., K.M. Baker y D.W. Weller. 2006. Mark–recapture abundance estimate of California coastal stock bottlenose dolphins: February 2004 to April 2005, SWFSC Administrative report LJ–06–02C. Southwest Fisheries Science Center, La Jolla, California. 15p. [ Links ]
Fulling, G.L., K. D. Mullin y C. W. Hubard. 2003. Abundance and distribution of cetaceans in outer continental shelf waters of the U.S. Gulf of Mexico. Fisheries Bulletin 101: 923–932. [ Links ]
Hammond, P. S., S. A. Mizroch .y G. P. Donovan. 1990. Individual recognition of cetaceans: use of photo–identification and other techniques to estimate population parameters. Reports of the International Whaling Commission. Special Issue 12. [ Links ]
Heckel, G. 1992. Fotoidentificación de tursiones Tursiops truncatus (Montagu, 1821) en la Boca de Corazones de la Laguna de Tamiahua, Veracruz, México, (Cetacea: Delphinidae) Tesis, Universidad Nacional Autónoma de México, México, D.F.164 p. [ Links ]
Irwin, L.J. y B. Würsig. 2004. A small resident community of bottlenose dolphins, Tursiops truncatus, in Texas: monitoring recommendations. G. Mex. Sci. 22:3–21. [ Links ]
IUCN. 2008. The IUCN red list of threatened species. 2008. International Union for Conservation of Nature and Natural Resources, http://www.iucnredlist.org/; 27.I.2010. [ Links ]
John, E. 2000. Bottlenose dolphins. Biology and conservation. University of Florida Press, Gainesville. 288 p. [ Links ]
Kelly, D. 1983. Photo–identification of bottlenose dolphins in southern California. Whalewatcher. 17:6–8. [ Links ]
Leatherwood, S. y R.R. Reeves. 1990. The bottlenose dolphin. Academic, New York, 653 p. [ Links ]
López Hernández, I. 1997. Ecología poblacional de las toninas Tursiops truncatus en la costa de Tabasco, México. Tesis, Facultad de Ciencias, Universidad Nacional Autónoma de México, México, D.F. 77 p. [ Links ]
Lusseau, D. 2005. The residency pattern of bottlenose dolphins (Tursiops spp.) in Milford Sound, New Zealand, is related to boat traffic. Marine Ecology Progress Series 295:265–272. [ Links ]
Mazzoil, M., S.. D. McCulloch, R. H. Defran, y M. E. Murdoch. 2004. The use of digital photography and analysis for dorsal fins photoidentification of bottlenose dolphins. Aquatic Mammals 30:209–219. [ Links ]
Merriman, M. G. 2007. The occurrence and behavioural ecology of bottlenose dolphins (Tursiops truncatus) in the Marlborough Sounds, New Zealand. MSc thesis, Massey University, Auckland. [ Links ]
Ortega–Ortiz, J. G. Delgado–Estrella A. y A. Ortega–Argueta. 2004 Mamíferos marinos del golfo de México, estado actual del conocimiento y recomendaciones para su conservación. In Diagnóstico ambiental del golfo de México, M. Caso, I. Pisanty y E. Exequiel (eds.). Secretaría de Medio Ambiente y Recursos Naturales/ Instituto Nacional de Ecología/ Instituto de Ecología/Harte Research Institute for Gulf of Mexico Studies, México, D.F. p 137–162. [ Links ]
Pollock, K.H., J.D. Nichols, C. Brownie y J.E. Hines. 1990. Statistical inference for capture–recapture experiments. Wildlife Monographs 107:1–97. [ Links ]
Reynolds, J. E., R. S. Wells y S. D. Eide. 2000. The bottlenose dolphin: biology and conservation. University Press of Florida, Gainesville. 304 p. [ Links ]
Schramm, Y. 1993. Distribución, movimientos, abundancia e identificación del delfín Tursiops truncatus (Montagu, 1821) en el Sur de la Laguna de Tamiahua, Ver. y aguas adyacentes (Cetacea: Delphinidae) Tesis, Universidad Autónoma de Guadalajara., Jalisco. 164 p. [ Links ]
Secretaría de Medio Ambiente y Recursos Naturales. 2002. Norma Oficial Mexicana NOM–059–ECOL–2001, Protección ambiental–Especies nativas de México de flora y fauna silvestre Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio–Lista de especies en riesgo. Diario Oficial de la Federación, 6 de marzo. [ Links ]
Serrano, A., L. Zavaleta–Lizárraga, I. Martínez–Serrano. 2007. Uso de delfines y tortugas marinas como indicadores del estado de salud del ecosistema marino en la zona norte del golfo de México. Informe técnico. Universidad Veracruzana–PEMEX Exploración y Producción, Tuxpan, Veracruz. 197 p. [ Links ]
Seber, G.A.F. 1982. The estimation of animal abundance and related parameters. segunda edición, Charles Griffin, Nueva York. 654 p. [ Links ]
Shane, S. H., R. S. Wells y B. Würsig. 1986. Ecology, behavior, and social organization of the bottlenose dolphin: a review. Marine Mammal Science 2:34–63. [ Links ]
Turner, E. R., 1977. Intertidal vegetation and commercial yields of peneids shrimp. Transactions of the American Fisheries Society. 106:411–416 p. [ Links ]
Weller, D. W. 1991. The social ecology of Pacific coast bottlenose dolphins. M. A. thesis, San Diego State University, California. 93 p. [ Links ]
Wells, R. y M. Scott. 1990. Estimating bottlenose dolphin population parameters from individual identification and capture–release techniques. In Individual recognition of cetaceans: use of photo–identification and other techniques to estimate population parameters, P. S. Hammond, S. A. Mizroch y G. P. Donovan (eds.). Reports of the International Whaling Commission Special issue 12:407–416 [ Links ]
Wells, R. y M. D Scott. 1999. Botlenose dolphins Tursiops truncatus (Montagu, 1821) In Handbook of marine mammals, vol. 6, The second book of dolphins and porpoises, Ridgway, S. H. y R. Harrison (eds.). Academic, San Diego, California. [ Links ].
Würsig, B. y T. A. Jefferson. 1990. Methods of photo–identification for small cetaceans. In Individual recognition of cetaceans: use of photo–identification and other techniques to estimate population parameters, P. S. Hammond, S. A. Mizroch y G. P. Donovan (eds.). Reports of the International Whaling Commission Special issue 12:43–52. [ Links ]

