










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de biodiversidad
versión On-line ISSN 2007-8706versión impresa ISSN 1870-3453
Rev. Mex. Biodiv. vol.81 no.2 México ago. 2010
Ecología
Diversidad de anfibios y reptiles de la Reserva de la Biosfera Barranca de Metztitlán, Hidalgo, México
Diversity of amphibians and reptiles from the Barranca de Metztitlán Biosphera Reserve in Hidalgo, Mexico
Victor D. Vite–Silva*, Aurelio Ramírez–Bautista y Uriel Hernández–Salinas
Laboratorio Ecología de Poblaciones, Centro de Investigaciones Biológicas, Universidad Autónoma del Estado de Hidalgo. Apartado postal 1–69, Plaza Juárez, Pachuca 42001, Hidalgo, México.
*Correspondencia:
victorv_23@yahoo.com.mx
Recibido: 24 enero 2009
Aceptado: 20 noviembre 2009
Resumen
La Reserva de la Biosfera Barranca de Metztitlán (RBBM) es una extensa área protegida (96 042.94 ha) de la zona centro del estado de Hidalgo, México. La reserva es bien conocida por su flora endémica, pero existe poca información disponible sobre su herpetofauna. En este estudio se examina la biodiversidad de la comunidad de anfibios y reptiles que habitan en cada uno de los 4 tipos de vegetación de la reserva: bosque de pino–encino, bosque tropical caducifolio, matorral submontano y matorral xerófilo. El trabajo de campo se llevó a cabo entre junio de 2006 y agosto de 2007, periodo de estación de lluvias y secas. En total, se registran 7 especies de anfibios y 31 de reptiles para la RBBM, distribuidas en 14 familias y 29 géneros. En todos los tipos de vegetación, la riqueza de especies de reptiles fue mayor en la estación de lluvias, mientras que la de especies de anfibios fue mayor en la de secas. Entre los tipos de vegetación, el bosque tropical caducifolio exhibió la diversidad y riqueza de especies más grande de anfibios y reptiles. El bosque de pino–encino presentó la mayor equidad y diversidad de especies que los otros tipos de vegetación. El matorral xerófilo y matorral submontano fueron los más similares entre sí en diversidad de especies. Este trabajo representa una aportación significativa al conocimiento de la herpetofauna de la RBBM, y una base para estudios futuros sobre historia natural de los anfibios y reptiles de esta reserva.
Palabras clave: riqueza de especies, diversidad, anfibios, reptiles, tipos de vegetación.
Abstract
The Barranca de Metztitlán Biosphere Reserve (BMBR) is an extensive protected area (96 042.94 ha) in Hidalgo state of central Mexico. The reserve is well known for its globally significant endemic flora, but relatively little information is available regarding its herpetofauna. We examined reptile and amphibian community biodiversity occurring in each of 4 vegetation types characteristic of the reserve: pine–oak forest, tropical deciduous forest, sub–montane shrubland, and arid tropical scrubland. Field work was carried out between June 2006 and August 2007, a period spanning both wet and dry seasons. Overall, we found 7 species of amphibians and 31 species of reptiles in the BMBR, distributed in 14 families and 29 genera. Across all vegetation types, reptile species richness was greater in the rainy season, whereas amphibian species richness was greater in the dry season. Among vegetation types, tropical deciduous forest exhibited the highest diversity and richness of amphibians and reptiles. Pine–oak forest exhibited greatest species evenness and species diversity than the other vegetation types. Arid tropical scrubland and sub–montane shrubland were more similar to each other in species diversity. This study represents a significant addition to our knowledge of the herpetofauna from the BMBR, and an important base for future studies on the natural history of amphibians and reptiles in this reserve.
Key words: species richness, diversity, amphibians, reptiles, vegetation types.
Introducción
México es uno de los países que cuentan con mayor riqueza florística y faunística, así como con una gran variedad de ecosistemas, producto de su topografía compleja que le confiere relieves variados (Ochoa–Ochoa y Flores–Villela, 2006). Estos factores fueron decisivos en los procesos evolutivos que dieron origen a la gran variedad de formas biológicas (Casas–Andreu et al., 2004). El conocimiento de la riqueza de especies de anfibios y reptiles de México ha sido motivo de significativas aportaciones científicas. Por sus factores taxonómicos, biogeográficos, de comportamiento y ecológicos, estos grupos resultan interesantes e indispensables en el buen funcionamiento de los ecosistemas naturales, y se les ha catalogado como organismos modelo por sus establecidos ciclos de vida y acelerados desarrollos embrionarios (Alcala y Brown, 1967; Tinkle et al., 1970; Ramírez–Bautista, 2004). México, por su inmejorable posición geográfica (ubicado entre la región neártica y neotropical), es considerado un corredor biológico que cuenta con una importante diversidad de flora y fauna (Morrone y Márquez, 2008).
El conocimiento de la herpetofauna mexicana se inicia con Francisco Hernández entre 1570 y 1577, quien menciona 68 especies para la Nueva España. Posteriormente con el trabajo de Martín del Campo (1936) y más tarde el de Smith y Taylor (1966), se incrementó significativamente lo que hasta entonces se sabía. Con estos estudios se establecieron las bases y la continuidad de las investigaciones (Smith y Smith, 1976a, 1976b, 1993), y permitieron el surgimiento de estudios que han explorado nuevas regiones para conocer la diversidad y la historia natural de este grupo de vertebrados (Ramírez–Bautista et al., 2006a).
En la actualidad, se han descrito más de 1 200 especies de anfibios y reptiles para México, aunque comparativamente con otros grupos biológicos, son los vertebrados menos conocidos en nuestro país. Esta riqueza de especies representan el 9.8% de la herpetofauna mundial (Ochoa–Ochoa y Flores–Villela, 2006), y se espera que este porcentaje aumente en el futuro, puesto que están en proceso de descripción varias especies (Ramírez–Bautista et al., 2006a). Más de la mitad de los taxones de anfibios y reptiles presentes en el territorio son endémicos de alguna provincia biogeográfica, lo que hace de México un área compleja, donde se conectan elementos con afinidades tanto neárticas como neotropicales (Flores–Villela y Canseco–Márquez, 2004; Morrone y Márquez 2008). A pesar de la diversidad de anfibios y reptiles registrada por Smith y Smith (1976a, 1976b), a la fecha no existe una obra completa y actualizada sobre estos grupos para México. Aún así, nuestro país está considerado megadiverso en anfibios y reptiles (Ramírez–Bautista et al., 2006a; Campbell y Flores–Villela, 2008).
Desde el punto de vista histórico y faunístico, existen algunas publicaciones que contemplan la riqueza de especies para algunas regiones del estado de Hidalgo (Martín del Campo, 1936; Smith y Taylor, 1966); sin embargo, los listados que contienen son limitados. Más tarde, este conocimiento fue aumentando con los estudios que se han realizado en otras áreas geográficas del estado (Mendoza–Quijano et al., 1989; Mendoza–Quijano, 1990; Camarillo–Rangel y Casas–Andreu, 1998; Ramírez–Pérez, 2008).
Debido a sus características naturales, la Reserva de la Biosfera Barranca de Metztitlán (RBBM) ha sido objeto de muchos trabajos científicos, destacando principalmente estudios sobre cactáceas (Hernández–Aguilar et al., 2007; López–López y Collazo–Ortega, 2007), climáticos (Hernández et al., 2007), y de insectos (Delgado y Márquez, 2006; Moreno et al., 2007). Sin embargo, en el conocimiento de la herpetofauna se muestra la misma situación que para otras áreas del estado de Hidalgo; los inventarios que se han generado son muy escasos y no representan cuantitativamente la composición taxonómica de la reserva. Este trabajo tiene como objetivo dar a conocer la composición de la herpetofauna de la RBBM, como una forma de incrementar el conocimiento de estos grupos que habitan en las principales comunidades vegetales presentes en la reserva; así también, pretende sentar las bases para estudios posteriores sobre historia natural de estos vertebrados.
Materiales y métodos
Área de estudio. La Reserva de la Biosfera Barranca de Metztitlán (RBBM) fue declarada como tal el 27 de noviembre del año 2000. Se localiza al este del estado de Hidalgo (98° 23' 00" O y 98° 57' 08" N y 20° 14' 15" O y 20° 45' 26" N), con una superficie de 96 042.94 ha, y una elevación de 1 000 a 2 000 m snm (Fig. 1). El clima predominante en la mayor parte de la reserva es semiseco. La temperatura va de 18 a 22°C, aunque en la zona NE, SE y en las partes más altas, las temperaturas medias anuales corresponden al intervalo de 16 a 18°C (CONANP, 2003).
La RBBM se encuentra entre la sierra Madre Oriental y la Faja Volcánica Transmexicana; esta ubicación geográfica y las características geológicas de la región le permiten tener una alta diversidad de tipos de vegetación (Rzedowski, 1978; Morrone, 2001), como el bosque de pino–encino (BPE), bosque tropical caducifolio (BTC), matorral submontano (MSM) y matorral xerófilo (MX). Esta clasificación se basa en los criterios florísticos, fisonómicos y ambientales de Rzedowski (1978), Flores–Mata et al. (1971) y Zamudio et al. (1992).
Para la elaboración de la lista de especies de anfibios y reptiles, se revisó la literatura previamente publicada sobre el tema y se consultó una base de datos de colecciones nacionales y extranjeras (Apéndice 1). La información obtenida de esta base fue generada en un periodo de 10 años (1995–2005), y se obtuvo de los datos proporcionados en formato impreso por los curadores de las colecciones de cada una de las universidades mencionadas en el Apéndice 1. Esta información permitió formar una lista preliminar de las especies de anfibios y reptiles de la RBBM y, a la vez , eliminar los registros de especies que se consideraron inciertas para la reserva, con base en la literatura sobre la distribución de cada especie (e.g., Smith y Taylor, 1966; Flores–Villela, 1993; Duellman, 2001; Campbell y Lamar, 2004; Canseco–Márquez et al., 2004; Parra–Olea et al., 2005; Mendoza–Quijano et al., 2006; Flores–Villela y Canseco–Márquez, 2007). Es importante aclarar que se utilizaron los registros de esta base de datos sólo para representar la riqueza de especies para el área en estudio y no para generar análisis de diversidad.
Trabajo de campo. El trabajo de campo en la RBBM consistió en muestreos sistemáticos, es decir, siguiendo un orden en cuanto a tiempo y esfuerzo horas/hombre, aplicado para cada uno. Los muestreos se realizaron entre los días 15 y 20 de cada mes; el primer periodo (temporada de secas) abarcó de febrero a abril de 2007, y el segundo (temporada de lluvias), de junio a agosto de 2007. Previo a éstos, se realizaron 2 muestreos adicionales dentro de la época de lluvias, en los meses de junio y septiembre de 2006, ya que durante esta época se acentúa la diversidad y abundancia de estos grupos y las especies de anfibios y reptiles son más visibles, puesto que inician su actividad reproductora (Ramírez–Bautista, 2004). En este trabajo se muestrearon 4 tipos de vegetación: BPE, BTC, MSM y MX, que fueron elegidos por tratarse de los más representativos y de mayor extensión territorial dentro de la reserva.
Los datos de riqueza de especies, así como los de diversidad y abundancia obtenidos durante todo el trabajo de campo sirvieron para generar una base de datos que se utilizó para formar la lista de especies que fue analizada con índices de diversidad basados en las especies registradas en los distintos tipos de vegetación antes mencionados.
Para homogeneizar los muestreos, el esfuerzo de éstos en cada tipo de vegetación fue el mismo, es decir, fue realizado por 3 personas y considerado como horas/ hombre (por ejemplo, una búsqueda = 2 horas por 2 personas = 4 horas persona), mediante este método se pudo ejemplificar la riqueza de especies y evaluar la completitud del inventario en una curva de acumulación (Southwood y Henderson, 2000; Vellend, 2001).
Los anfibios y reptiles se recolectaron realizando recorridos (caminatas) en cada tipo de vegetación, uno durante el día y otro por la noche, hasta completar 6 muestreos para cada tipo de vegetación, y obteniendo al final del trabajo un total de 22 muestreos. Durante el trabajo de campo se contó con el permiso de colecta SEMARNAT–08–017–A, con Número de Registro Ambiental (NRA): HESSX1304811.
Los muestreos diurnos se realizaron según la técnica descrita por Casas–Andreu et al. (1991), modificada de acuerdo con los hábitos de cada especie (hora de inicio y fin de la actividad de los organismos). El horario de muestreo para especies con actividad diurna, que generalmente son los reptiles, fue de 09:00 a 13:00 h y de 14:00 a 18:00 h (Ramírez–Bautista, 1994; Uribe–Peña et al., 1999). Algunos organismos con actividad diurna fueron recolectados directamente con la mano, aunque para especies evasivas o muy rápidas (por ejemplo, Aspidoscelis gularis) fue necesario hacerlo con ligas. Para las especies venenosas (por ejemplo, género Crotalus) se usó un gancho herpetológico. Los organismos se buscaron en los diferentes tipos de microhábitats; bajo rocas, sobre y bajo troncos y a orillas de los cuerpos de agua, entre otros (Casas–Andreu et al., 1991). El horario para las especies con actividad nocturna se estableció a partir de las 19:00 a las 23:00 h, y fue aplicado especialmente para los anfibios (Casas–Andreu et al., 1991; Ramírez–Bautista, 1994; Uribe–Peña et al., 1999). Para la recolecta de especies de este grupo que habitan dentro de cuerpos de agua, se utilizó una red de malla con un mango, así como también lámparas de mano para facilitar su captura.
Los datos obtenidos directamente en el campo fueron: fecha y hora de recolecta, temporada en el que fueron encontrados (secas o lluvias), tipo de vegetación, altitud y coordenadas geográficas. Esta información se utilizó para elaborar una base de datos en el programa Excel, y de esta forma estimar la riqueza potencial de especies en cada tipo de vegetación. Todos los ejemplares fueron reconocidos hasta especie con las diferentes claves dicotómicas para cada grupo; por ejemplo, Smith y Taylor (1966) para grupos específicos de anfibios y reptiles (Rossman et al., 1996; Duellman, 2001; Bezy y Camarillo, 2002; Campbell y Lamar, 2004), así como con las descripciones originales de las especies (por ejemplo, Pérez–Ramos et al., 2000). Además, se actualizaron los nombres científicos de acuerdo a la literatura más reciente para cada grupo taxonómico (Ramírez–Bautista et al. 2006a; Frost et al., 2008).
Los organismos recolectados para este trabajo, y durante su identificación se depositaron en el laboratorio de Ecología de Poblaciones, perteneciente al Centro de Investigaciones Biológicas (CIB) de la Universidad Autónoma del Estado de Hidalgo (UAEH). Posteriormente, al finalizar este trabajo todos serán depositados en la colección de anfibios y reptiles del CIB–UAEH.
Análisis de datos. Se elaboró una curva de acumulación de especies con la información obtenida en campo. Esta es una herramienta importante para evaluar la biodiversidad y el esfuerzo de muestreo con base a las especies observadas y esperadas; así también, señala en qué número de muestreo se alcanza la asíntota, indicando que ya es momento de finalizar el trabajo de campo en el sitio o región, o bien, la curva de acumulación puede indicar que aún no se alcanza la completitud del inventario (Moreno y Halffter, 2000; Colwell et al., 2005).
En este análisis se utilizó el estimador no paramétrico Chao 2, en el que se utilizaron datos de incidencia (presencia y ausencia). Adicionalmente, se tomaron en cuenta algoritmos que emplean proporciones de especies raras (uniques y duplicates; que aparecen en 2 muestras, respectivamente), en el supuesto de que cuando estas 2 líneas se cruzan, el inventario está completo (Jiménez–Valverde y Hortal, 2003); la curva de acumulación de especies se generó con ayuda del programa EstimateS v 8.0.0 (Colwell, 2006).
La riqueza de especies para cada tipo de vegetación se evaluó empleando el índice de Shannon–Wiener, el cual suele utilizarse entre los ecólogos por ser uno de los índices clásicos y fáciles de interpretar. Este índice considera estadísticamente a la comunidad como una muestra finita y es sensible al cambio entre el número de especies (Halffter et al., 2001; Feisinger, 2003); además, actúa bajo la suposición de que los individuos se muestrean al azar a partir de una población inmensamente grande, y que todas las poblaciones de las especies están presentes en una sola muestra (Magurran, 1988, 2004).
También se reconoció el valor de equidad, utilizando el índice de Pielou que estima el valor de equidad en términos de abundancia, tomando como base las especies más representativas (dominantes) y raras (poco comunes) presentes en las diferentes comunidades analizadas (Halffter et al., 2001; Henderson y Seaby, 2002).
Para ambos índices se realizó un remuestreo (bootstrap) que permite conocer si sus valores son estadísticamente diferentes. Los resultados de estos análisis fueron generados con el programa Species Diversity and Richness III (Henderson y Seaby, 2002). Simultáneamente, para poder comparar los patrones de abundancia de especies entre los diferentes tipos de vegetación, se elaboraron curvas de Whittaker o de rango–abundancia, indicando sobre el eje de las X el número de especies registradas o representadas en cada tipo de vegetación, mientras que en el eje de las Y se refleja la abundancia relativa de cada especie en cada comunidad vegetal (Magurran, 1988; Feinsinger, 2003).
Finalmente, se calculó el grado de similitud de la riqueza de especies entre tipos de vegetación, elaborando un dendograma de similitud mediante un análisis cluster empleando el índice de Jaccard con ayuda del programa BioDiversity Professional (Magurran, 2004).
Resultados
Con 462 horas de esfuerzo de muestreo, aplicado en los 22 muestreos realizados en la RBBM, se registró un total de 26 especies; 5 correspondieron a anfibios y 21 a reptiles (Cuadro 1). Con la revisión de la literatura y base de datos, se agregaron 12 especies más (2 anfibios y 10 reptiles); la lista final fue de 38 especies (18.4% anfibios) y (81.5% reptiles) para la región (Cuadro 2). El grupo de anfibios de la reserva está formado por 7 géneros, los que se encuentran dentro de 5 familias (Cuadro 2). El suborden Sauria se encuentra representado con 6 géneros dentro de 4 familias, y el suborden Serpentes está representado con 16 géneros, ubicados en 5 familias (Cuadro 2).
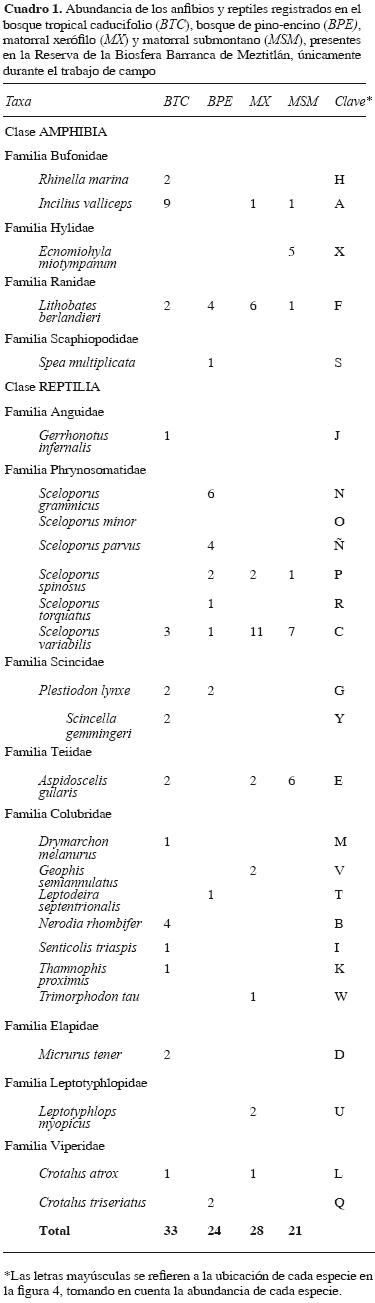

Riqueza de especies. Tomando en cuenta los datos obtenidos en campo, la riqueza de especies de anfibios y reptiles varió por 6 especies entre temporadas. En la temporada de secas se registraron 16 especies de anfibios y de reptiles, mientras que en la de lluvias fueron 22. En la temporada de secas, las más abundantes fueron los saurios, con 6 especies, superando con 1 a las serpientes y a los anuros. En contraste, en la temporada de lluvias se presentó un incremento en el número de especies para el grupo de los reptiles, mostrando a los saurios como el grupo más abundante junto con las serpientes, con una frecuencia de encuentro de 9 especies en ambos grupos; contrario a los anuros, de los que sólo se registraron 4 especies (1menos que en la época de secas).
En la curva de acumulación se observó que el número de especies aumenta en función del esfuerzo de muestreo, sin embargo, con los datos obtenidos en campo (26 especies de anfibios y reptiles), la curva comienza una fase asintótica, aproximadamente en el muestreo 20, con un total de 17 a 19 especies, indicando que aún hacen falta especies y muestreos que agregar al inventario (Fig. 2). El estimador empleado (Chao 2) predice un potencial de 33 especies esperadas para la RBBM (Fig. 2), lo que indica que falta por registrar cerca del 24% para completar el inventario. Por su parte, los estimadores uniques y duplicates no llegan a sobreponerse, ya que están identificando en todo el inventario especies raras o únicas (Cuadro 1). Esto significa que el listado está próximo a completarse; sin embargo, los estimadores deben identificar sólo especies repetidas o duplicates en todo el inventario para que éstos inicien un entrecruce y la curva de riqueza observada alcance una fase completamente asintótica, indicando que ya no es posible registrar más especies (Fig. 2).
El tipo de vegetación con mayor número de especies fue el BTC, con 14 especies, seguido por el BPE y el MX, con 10 y 9, respectivamente, superando al MSM que presentó 6 especies. La diversidad calculada en los 4 tipos de vegetación de acuerdo con el índice de Shannon–Wiener, mostró que el BTC presentó el valor más alto de diversidad (H=2.3) y el MSM el más bajo (H'=1.7, Fig. 3). Por otra parte, el grado de equidad en la distribución de la abundancia de las especies entre los tipos de vegetación, utilizando el índice de Pielou (J'), indicó que la equidad entre los tipos de vegetación fue muy poco homogénea (Fig. 3); sin embargo, el valor más alto fue para el BPE (J'= 0.92), y el más bajo para el MX (J'= 0.80), indicando que en el BPE, las especies presentan valores de abundancia muy similares, mientras que la abundancia entre las especies del MX es más heterogénea (Cuadro 1).
Patrones de abundancia. De un total de 106 registros de anfibios y reptiles en la zona de estudio, el 31.1% se presentó en el BTC, 22.6% en el BPE, 26.4% en el MX, y el 19.8% para el MSM (Cuadro 1). Visualmente, las curvas de rango–abundancia están claramente dominadas por especies raras (especies con las letras IJKLM, RST, WLA y FAP; Cuadro 1 y Fig. 4). Sólo Lithobates berlandieri y Sceloporus variabilis estuvieron presentes en los 4 tipos de vegetación, mostrándose moderadamente abundantes. Sin embargo, no se mantiene un orden jerárquico en los diferentes tipos de vegetación, es decir, no son completamente dominantes, ni raras (Fig. 4). Spea multiplicata fue la especie de anfibio poco frecuente en las 4 comunidades vegetales, puesto que sólo se registró en una ocasión dentro del BPE (Cuadro 1). Mientras que Incilius valliceps fue moderadamente frecuente en el BTC y MX, pero rara en MSM (Fig. 4).
En relación con los reptiles, el mayor número de especies raras se registró en el BTC. Gerrhonotus infernalis, Drymarchon melanurus, Senticolis triaspis, Thamnophis proximus y Crotalus atrox, se ubican como poco frecuentes en este tipo de vegetación (Cuadro 1). Finalmente, Aspidoscelis gularis fue la única especie poco frecuente en el BTC y MX, y con una abundancia moderadamente alta en el MSM (Cuadro 1, Fig. 4).
Por su parte, el índice de similitud indicó que los tipos de vegetación con mayor similitud en composición herpetofaunística fueron el MX y MSM, con un porcentaje de 38%, mientras que los menos similares fueron el MX y BPE, con un porcentaje de 17% (Fig. 5).
Discusión
El estado de Hidalgo representa el 1.1% de la superficie total del país (CONANP, 2003); el área de estudio tiene una extensión de 961.63 km2 (CONANP, 2003), es decir, el 0.05% de la superficie total de México. Por lo que la extensión que ocupa la RBBM presenta una importante composición de familias, géneros y especies de anfibios y reptiles con respecto a las que habitan en nuestro país (Flores–Villela y Canseco–Márquez, 2004).
Del total de especies (38) que aquí se registran, 26 son del trabajo de campo y el resto (12) de la literatura y de la base de datos. Es un número taxonómico confiable para esta reserva y una información de la que se carecía (Mendoza–Quijano et al., 2006). Para la RBBM se tenía un número de 29 especies provenientes de la literatura (CONANP, 2003), de las que algunas era dudosa su presencia en la reserva. En este trabajo se corrobora la presencia de algunas especies previamente mencionadas en el Programa de Manejo de la RBBM (CONANP, 2003). Se agregan al listado 1 especie de anfibio (Rhinella marina) y 9 de reptiles, entre las que destaca Micrurus tener, siendo ésta un registro nuevo para el área de estudio, con el que amplía su área de distribución en el estado de Hidalgo (Hernández–Salinas et al., 2008).
Como se advierte en la figura 4, las curvas de rango–abundancia estuvieron marcadamente dominadas por las especies poco comunes o raras. En las curvas de esta figura existe poca equidad en cuanto a la abundancia de todas las especies, ya que pocas son dominantes (parte superior de las curvas) y muchas son raras (parte inferior de las curvas). Sin embargo, en el caso de los anfibios sólo Incilius valliceps y Lithobates berlandieri se distribuyen en casi todos los tipos de vegetación, por lo que es probable que las condiciones de humedad, temperatura, disponibilidad de alimento y la altitud ofrezcan los requerimientos necesarios para que éstas tengan éxito en estos ambientes. Este mismo patrón observaron Urbina–Cardona y Reynoso (2005) en un gradiente potrero–borde–interior de la Reserva de Los Tuxtlas, Veracruz, al reconocer los patrones de abundancia de anfibios y reptiles de esa región.
En los reptiles, las lagartijas fue el grupo que mejor se representó en la reserva; por ejemplo, Aspidoscelis gularis se encontró en todos los tipos de vegetación, excepto en el MX, Sceloporus spinosus, en todos excepto en BTC, y Sceloporus variabilis en todos los tipos de vegetación. Lo anterior podría indicar que los factores climáticos, como temperatura, precipitación y fotoperiodo que imperan durante la estación de secas y lluvias en estos tipos de vegetación pueden favorecer o limitar la presencia de estas especies en los diferentes ambientes (Ramírez–Bautista et al., 2006b).
La baja riqueza de especies de anfibios en la región podría ser una respuesta a las altas temperaturas que predominan durante la estación de secas y lluvias; además, en esta reserva, los cuerpos de agua permanentes son muy escasos, sólo se establecen de forma temporal en la época de lluvias (CONANP, 2003). A pesar de que se sabe que las zonas agrícolas de la región, las fertilizan con agroquímicos que indudablemente contaminan el ambiente de los anfibios, a la fecha no existe información documentada que indique que es un factor que en parte determina la declinación de algunas poblaciones y diversidad de este grupo de vertebrados (Moreno et al., 2007).
El clima y la temperatura, así como la altitud son factores que promueven la distribución, diversidad y abundancia de los anfibios y reptiles, por lo que estos grupos pudieran estar respondiendo de manera similar, como ocurre en otros estudios y con otros grupos (Ramírez–Bautista y Moreno, 2006; Moreno et al., 2007). La Reserva de Metztitlán presenta 2 estaciones bien marcadas, una de secas y otra de lluvias (CONANP, 2003), esto permite que varíen la humedad y los intervalos de temperatura a lo largo del año, al igual que otras áreas naturales de México (Ramírez–Bautista, 1994). La variación de factores como los antes mencionados podría estar determinando la disponibilidad del recurso alimenticio, por lo tanto, la abundancia y distribución a diferentes escalas, espacial y temporal en poblaciones de anfibios y reptiles de la zona, es de manera similar a los resultados de diferentes trabajos (Vilá, 1998; Arita y Rodríguez, 2001; Pineda et al., 2005).
Considerando ambas estaciones, en la de secas se registró menor riqueza de especies (11) que en la de lluvias (18 especies); esta variación podría explicarse con respecto a la parte más húmeda del año cuando el recurso alimento es más abundante que en la parte más seca, por lo que los organismos en general, y en este caso los anfibios y reptiles, se ven favorecidos por el acceso al alimento, como se informa en otros estudios (Hernández–Ibarra, 2005; Estrada–Rodríguez et al., 2006; Garza–Castro et al., 2006).
En la figura 2 se muestra que el estimador Chao 2 presenta una fase asintótica, la cual debería ser simulada por la curva de riqueza observada. Sin embargo, como se indicó en la sección de resultados, el número de muestreos y el de especies raras hacen que aún no sea posible que la curva de riqueza observada alcance una fase asintótica definida, considerando que aún se requiere de un mayor esfuerzo de muestreo, el cual debe aplicarse a los pequeños manchones de vegetación aislados de la reserva que no fueron considerados en este estudio (Fig. 2). En cuanto a las proporciones de especies raras (uniques y duplicates) que no se sobreponen, indica que el inventario aún está incompleto (Fig. 2); por lo tanto, es muy probable la existencia de otras especies por registrar. Bajo este supuesto, podemos aceptar que mientras más especies raras haya, mayor será el número de especies que quedan por registrar en el inventario (Jiménez–Valverde y Hortal, 2003). Sin embargo, las 12 especies que se tomaron de la literatura y que no se incluyen en los análisis, podrían representar las especies que durante el trabajo de campo no se encontraron.
El índice de Shannon–Wiener mostró que el BTC fue el de mayor diversidad y riqueza de especies, mientras que el MSM fue el menos diverso. Esta diferencia de riqueza de especies podría explicarse bajo un enfoque ecológico, similar a lo mostrado por Pianka (1966) y Kucera (1978), indicando que la mayor riqueza de especies de diferentes grupos suelen encontrarse en bosques tropicales. Nuestros resultados suelen concordar con la idea anterior y con trabajos similares de selvas tropicales (Vargas–Contreras y Hernández–Huerta, 2001).
Finalmente, los valores de equidad mostraron diferencias entre los 4 tipos de vegetación, indicando que los individuos de las especies registradas no se encuentran distribuidos de manera uniforme en los tipos de vegetación analizados (Broker y Zar, 1979), lo que puede atribuirse a que cada uno representa hábitats heterogéneos, donde cada formación vegetal alberga una diversidad de recursos disponibles (microhábitats, alimento, espacio, etc.), por lo tanto, las especies que habitan en estos ambientes heterogéneos, probablemente no tendrán una repartición homogénea de recursos entre los individuos y especies. Este patrón fue semejante al de un estudio sobre el ensamble de anfibios de un bosque mesófilo de montaña (Pineda et al., 2005) y a otro sobre la comparación de la herpetofauna de la zona centro de México (Ramírez–Bautista y Moreno, 2006).
En la figura 3 y cuadro 1, se muestran las diferencias de similitud entre tipos de vegetación, mostrando que las formaciones vegetales con menos especies exclusivas son el MSM y MX, 3 para el MX (Geophis semiannulatus, Trimorphodon tau y Leptotyphlops dulcis) y sólo una para el MSM (Ecnomiohyla miotympanum), siendo ésta la única especie que habita de forma exclusiva en un ambiente árido (Cuadro 1). Esto indica que las condiciones ambientales que imperan tanto en el MX como en el MSM suelen ser poco favorables para algunas especies de anfibios, puesto que en estas comunidades vegetales existe una mayor aridez y una concentración más elevada de sales, acompañada por pocos cuerpos de agua, los cuales son necesarios para llevar a cabo los ciclos biológicos y desarrollo embrionario de los anfibios (Duellman y Trueb, 1986; Duellman y Campbell, 1992; Duellman, 2001; Zug et al., 2001). En contraste, en el cuadro 1 se muestra una mayor cantidad de especies de reptiles exclusivas para el MX y MSM. Las condiciones ambientales que prevalecen en estas formaciones vegetales suelen ser favorables para los reptiles. En general este grupo es menos vulnerable a estos tipos de ambientes que los anfibios, principalmente por 3 condiciones: 1) los reptiles tiene una piel altamente impermeable, lo que les ayuda a perder poca agua; 2) sus huevos (especies ovíparas) presentan un cascarón que los protege de condiciones áridas, lo contrario a los anfibios, que en toda su historia de vida, necesariamente debe estar ligada a lugares con suficiente humedad, y 3) las temperaturas que existen en ambientes áridos aceleran significativamente al desarrollo embrionario de las especies vivíparas (Shine y Bull, 1979; Blackburn, 1984; Wyman, 1990; Blaustein y Wake, 1995; Sánchez, 1999).
Como se mencionó en la parte introductoria, la RBBM se encuentra ubicada entre 2 regiones biogeográficas importantes, la neártica (donde se ubican los tipos de vegetación templado–cálidos y fríos, que alcanzan altitudes de los 500 a más de 4 000 m snm; Morrone, 2001) y la neotropical (donde se ubican los principales tipos de vegetación de clima tropical, alcanzando altitudes que van desde pocos metros al nivel del mar hasta aproximadamente 3000 m snm). Por lo tanto, la RBBM es un área geográfica con una importante heterogeneidad de tipos de vegetación y rangos altitudinales variables que como consecuencia presenta una alta diversidad de flora y fauna (Moreno et al., 2007). Así que las especies que se ubican en el BTC estarían representando especies con afinidades neotropicales, y las que están en el BPE, MX y MSM, a la herpetofauna con afinidades neárticas (Fig. 5). Lo anterior indica que la heterogeneidad de tipos de vegetación, humedad y temperatura, refleja una herpetofauna con mayor complejidad a la observada en otras áreas naturales con ambientes similares de México; por ejemplo, el Parque Nacional Malinche (Gómez–Álvarez y Reyes–Gómez, 2006), Parque Nacional de El Mineral de El Chico (Ramírez–Pérez, 2008) y las Áreas Naturales protegidas de Chichinautzin y Sierra de Huautla (Castro–Franco y Bustos–Zagal, 2006).
Las especies hasta ahora registradas en este estudio, son un acercamiento al conocimiento de la distribución geográfica de la herpetofauna de México, y en particular, de los anfibios y reptiles del estado de Hidalgo. Sin embargo, este conocimiento debe ser acompañado por otros estudios de estructura, dinámica de poblaciones y de ecología de comunidades para poder entender claramente la importancia que tienen estos grupos biológicos en esta zona, y promover la necesidad de estructurar estrategias de conservación más finas para protegerlos en diversas área naturales (Lips, 1998; Berger et al., 1998; Sánchez, 1999).
Agradecimientos
A las autoridades de la Reserva de la Biosfera Barranca de Metztitlán (RBBM) por el apoyo brindado para realizar este estudio. A B. P. Stephenson, J. Roberto Hernández, Alejandro Ramírez Pérez y Adrián Leyte Manrique por el apoyo que nos otorgaron durante el trabajo de campo. A los proyecto de PROMEP, Cuerpo Académico de Ecología, CONACYT S52552–Q y Diversidad Biológica del Estado de Hidalgo FOMIX–CONACYT 43761, FOMIX–HGO–2008–95828.
Literatura citada
Alcala, A. C. y W. C. Brown. 1967. Population ecology of the tropical scincoid lizard, Emoia atrocostata, in the Philippines. Copeia 3:596–604. [ Links ]
Arita, H. T. y P. Rodríguez. 2001. Ecología geográfica y macroecología. In Introducción a la biogeografía en Latinoamérica: teorías, conceptos, métodos y aplicaciones, J. Llorente Bousquets y J. J. Morrone (eds.). Comisión Nacional para el Conocimiento y Uso de la Biodiversidad y Universidad Nacional Autónoma de México. México D.F. p. 63–80. [ Links ]
Berger, L., R. Speare, P. Daszak, D. E. Green, A. A. Cunningham, C. L Goggin, R. Slocombe, M. A. Ragan, A. D. Hyatt, K. R. McDonald, H. B. Bines, K. R. Lips, G. Maranteli y H. Parkes. 1998. Chytridiomucosis causes amphibian mortality associate with population declines in the rainforests of Australia and Central America. Proccedings of the National Academy pf Sciences 95:9031–9036. [ Links ]
Bezy, R. L. y J. L. Camarillo. 2002. Systematics of xantusiid lizards of the genus Lepidophyma. Scientific Contribution Natural History Museum of Los Angeles 493:1–41. [ Links ]
Blackburn, D. G. 1984. Euterian like reproductive specialization in viviparous reptile. Proceedings of the Natural Academic Society 81:4860–4863. [ Links ]
Blaustein, A. R. y D. B. Wake. 1995. The puzzle of declining amphibian population. Scientific American 272:56–61. [ Links ]
Brower, J. E. y J. H. Zar. 1979. Field and laboratory methods for general ecology. Brown, USA. 194 p. [ Links ]
Canseco–Márquez, L., F. Mendoza–Quijano y M. G. Gutiérrez–Mayén. 2004. Análisis de la distribución de la herpetofauna. In Biodiversidad de la sierra Madre Oriental, I. Luna–Vega, J. J. Morrone y D. Esparza (eds.). Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Universidad Nacional Autónoma de México, México, D. F. p. 417–438. [ Links ]
Camarillo–Rangel, J. L. y G. Casas–Andreu. 1998. Notas sobre la herpetofauna del área comprendida entre Zacualtipán, Hidalgo y Huayacocotla, Veracruz. Anales del Instituto de Biología, Universidad Nacional Autónoma de México. Serie Zoología 69:231–237. [ Links ]
Campbell, J. A. y O. Flores–Villela. 2008. A new long–tailed rattlesnake (Viperidae) from Guerrero, México. Herpetologica 64:246–257. [ Links ]
Campbell, J. A. y W. W. Lamar. 2004. The venemous reptiles of the western hemisphere, vol. I y II. Cornell University, Ithaca, New York. [ Links ]
Casas–Andreu, G., F. R. Méndez de la Cruz y X. Aguilar–Miguel. 2004. Anfibios y reptiles. In Biodiversidad de Oaxaca, A. J. García–Mendoza, M. J. Ordóñez y M. Briones–Salas (eds.). Instituto de Biología, Universidad Nacional Autónoma de México/ Fondo Oaxaqueño para la Conservación de la Naturaleza / World Wildlife Fund, México, D.F. p. 375–390. [ Links ]
Casas–Andreu, G., G. Valenzuela–López y A. Ramírez–Bautista. 1991. Cómo hacer una colección de anfibios y reptiles. Cuadernos del Instituto de Biología, UNAM, México, D.F. 68 p. [ Links ]
Castro–Franco, R. y M. G. Bustos–Zagal. 2006. Herpetofauna de las áreas naturales protegidas corredor biológico Chichinautzin y la sierra de Huautla Morelos, México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, D.F. / Universidad Autónoma del Estado de Morelos, Cuernavaca. [ Links ]
Colwell, R. K. 2006. EstimateS: Statistical estimation of species richness and shared species from samples. Versión 8.2. Users guide and application. http://purl.oclc.or/estimates; última consulta: 14.IV.2007 [ Links ]
Colwell, R. K., C. Xuan–Mao y Jing–Chang. 2005. Interpolando, extrapolando y comparando las curvas de acumulación de especies basadas en su incidencia. In Sobre diversidad biológica: el significado de las diversidades alfa, beta y gamma, G. Halffter, J. Soberón, P. Koleff y A. Melic (eds.). Monografías 3er Milenio. SEA/CONABIO/Diversitas/CONACYT/, Zaragoza. p. 73–84. [ Links ]
CONANP (Comisión Nacional de Áreas Naturales Protegidas). 2003. Programa de manejo Reserva de la Biosfera Barranca de Meztitlán. SEMARNAT. México, D.F. 202 p. [ Links ]
Delgado, L. y J. Márquez. 2006. Estado del conocimiento y conservación de los coleópteros Scarabaenidae (insecta) del estado de Hidalgo. Acta Zoológica Mexicana (n.s.) 22:57–108. [ Links ]
Duellman, W. E. y L. Trueb. 1986. Biology of amphibians. McGraw–Hill, New York. [ Links ]
Duellman, W. E. y J. A. Campbell. 1992. Hylid frogs of the genus Plectrohyla: Sistematics and phylogenetics relationships. Miscellaneous Publication Museum of Zoology University of Michigan 181:1–32. [ Links ]
Duellman, W. E. 2001. The hylid frogs of Middle America. Museum of Natural History, University of Kansas, Lawrence. 1159 p. [ Links ]
Estrada–Rodríguez, J. L., H. Gadsden, S. V. Leyva–Pacheco y T. U. Morones–Long. 2006. Herpetofauna del cañón "Piedras Encimadas", sierra "El Sarnoso", Durango, México. 123. In Inventarios herpetofaunísticos de México: avances en el conocimiento de su biodiversidad, 3. A. Ramírez–Bautista, L. Canseco–Márquez y F. Mendoza Quijano (eds.). Publicaciones de la Sociedad Herpetológica Mexicana. México, D. F. p. 1–23. [ Links ]
Feinsinger, P. 2003. El diseño de estudio de campo para la conservación de la biodiversidad. FAN, Santa Cruz de la Sierra [Bolivia]. 242 p. [ Links ]
Flores–Mata, G., J. Jiménez–López, X. Madrigal–Sánchez, F. Moncayo–Ruíz y F. Takaki–Takaki. 1971. Memoria del mapa de tipos de vegetación de la República Mexicana. Dirección de Agrología, Secretaría de Recursos Hidráulicos, México, D. F. [ Links ]
Flores–Villela, O. A. 1993. Herpetofauna mexicana: lista anotada de las especies de anfibios y reptiles de México, cambios taxonómicos recientes, y nuevas especies. Special Publication 17. Carnegie Museum of Natural History. Pittsburgh, Pennsylvania. 73 p. [ Links ]
Flores–Villela, O. y L. Canseco–Márquez. 2004. Nuevas especies y cambios taxonómicos para la herpetofauna de México. Acta Zoológica Mexicana (n.s.) 20:1–31. [ Links ]
Flores–Villela, O. y L. Canseco–Márquez. 2007. Riqueza de la herpetofauna. In Biodiversidad de la Faja Volcánica Transmexicana, I. Luna–Vega, J. J. Morrone y D. Espinosa (eds.). Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Universidad Nacional Autónoma de México. p 407–420. [ Links ]
Frost, D. R., T. Grant, J. Faivovich, R. H. Bain, A. Haas, C. F. B. Haddad, R. O. de Sa, A. Channing, M. Wilkinson, S. C. Donnellan, C. J. Raxworthy, J. A. Campbell, B. L. Blotto, P. Moler, R. C. Drewes, R. A. Nussbaum, J. D. Lynch, D. M. Green y W. C. Wheeler. 2008. Is the Amphibian tree of life really fatally flawed?. Cladistics 24:385–395. [ Links ]
Garza–Castro, J. M., F. H. Carmona–Torres y A. J. González–Hernández. 2006. Anfibios y reptiles del ejido San Juan Raya, Municipio de Zapotitlán de las Salinas, Puebla. In Inventarios herpetofaunísticos de México: avances en el conocimiento de su biodiversidad 3, A. Ramírez–Bautista, L. Canseco–Márquez y F. Mendoza Quijano (eds.). Publicaciones de la Sociedad Herpetológica Mexicana, México, D. F. p. 170–179. [ Links ]
Gómez–Álvarez, G. y S. R. Reyes–Gómez. 2006. Anfibios y reptiles del Parque Nacional La Malinche, estado de Tlaxcala. In Inventarios herpetofaunísticos de México: avances en el conocimiento de su biodiversidad 3, A. Ramírez–Bautista, L. Canseco–Márquez y F. Mendoza Quijano (eds.). Publicaciones de la Sociedad Herpetológica Mexicana, México, D. F. p. 241–250. [ Links ]
Halffter, G., C. E. Moreno y E. O. Pineda. 2001. Manual para evaluación de la biodiversidad en reservas de la biosfera, vol. 2, M & T–Manuales y Tesis, Sociedad Entomológica Aragonesa (SEA), Zaragoza. 80 p. [ Links ]
Henderson, P. A. y R. M. H. Seaby. 2002. Species diversity and richness III. Versión 3.02. Pisces Conservation, Ltd. http://www.pisces–conservation.com/softdiversity.html; última consulta: 14.IV.2007. [ Links ]
Hernández–Aguilar, A., G. Ramírez–Ramírez y M. Collazo–Ortega. 2007. Ecofisiología de Echinocactus platyacanthus (región Los Venados), en la Reserva de la Biosfera Barranca de Metztitlán, Hidalgo. In IV Foro de investigación por la conservación y II Simposio de áreas naturales protegidas del estado de Hidalgo, G. Pulido–Flores y A. L. López–Escamilla (eds.). Comisión Nacional de Áreas Naturales Protegidas y Universidad Autónoma del Estado de Hidalgo, 16–17 octubre del 2007. p. 37–43. [ Links ]
Hernández, L., J. D. Gómez Díaz y A. I. Montero Rivas.2007. Estimación del balance de humedad para la Cuenca de Amajac que incluye la Reserva de la Biosfera Barranca de Metztitlán bajo condiciones actuales y escenario de cambio climático. In IV Foro de investigación por la conservación y II Simposio de áreas naturales protegidas del estado de Hidalgo. G. Pulido–Flores y A. L. López–Escamilla (eds.). Comisión Nacional de Áreas Naturales Protegidas y Universidad Autónoma del Estado de Hidalgo, 16–17 octubre del 2007. p. 53–61. [ Links ]
Hernández–Ibarra, X. 2005. Biodiversidad de la herpetofauna del Municipio de Guadalcázar, San Luis Potosí, México. Tesis Facultad de Ciencias, Universidad Nacional Autónoma de México, México, D.F. 271 p. [ Links ]
Hernández–Salinas, U., V. D. Vite–Silva y A. Ramírez–Bautista. 2008. Micrurus tener (Texas coralsnake). Herpetological Review 39:242. [ Links ]
Jiménez–Valverde, A. y J. Hortal. 2003. Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de Aracnología 8:151–161. [ Links ]
Kucera, C. L. 1978. The challenge of ecology, segunda edición. C. V. Mosby, Saint Louis, Missouri. 326 p. [ Links ]
Lips, K. R. 1998. Decline of a tropical montane amphibian fauna. Conservation Biology 12:106–117. [ Links ]
López–López, J. y V. M. Collazo–Ortega. 2007. Banco de semillas de Echinocactus platyacanthus Link y Otto (Cactacea) y especies asociadas en una región semiárida de la Reserva de la Biosfera Barranca de Metztitlán, Hidalgo. In IV Foro de investigación por la conservación y II Simposio de áreas naturales protegidas del estado de Hidalgo. G. Pulido–Flores y A. L. López–Escamilla (eds.). Comisión Nacional de Áreas Naturales Protegidas y Universidad Autónoma del Estado de Hidalgo, 16–17 octubre del 2007. p. 45–51. [ Links ]
Magurran, A. E. 1988. Ecological biodiversity and its mesurement, primera edición. Princeton University Press, New Jersey . 179 p. [ Links ]
Magurran, A. E. 2004. Measuring biological diversity. Blackwell, Oxford. 256 p. [ Links ]
Martín del Campo, R. 1936. Contribución al conocimiento de la fauna de Actopan, Hidalgo. IV. Vertebrados observados en la época de secas. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología 7:271–286. [ Links ]
Mendoza–Quijano, F. 1990. Estudio herpetofaunístico en el transecto Zacualtipán– Zoquizoquiapan– San Juan Metztitlán, Hidalgo. Tesis, Escuela Nacional de Estudios Profesionales Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México. 94 p. [ Links ]
Mendoza–Quijano, F., A. Gonzáles–Alonso y M. Mancilla Moreno. 1989. Notas sobre algunos anfibios y reptiles del oeste de Tulancingo, Hidalgo. Memorias del IX Coloquio de Investigación. Escuela Nacional de Estudios Profesionales Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México. 4–16 diciembre de 1989. [ Links ]
Mendoza–Quijano, F., G. Quijano–Manilla y R. F. Mendoza–Paz. 2006. Análisis fenético de la herpetofauna de los bosques mesófilos de montaña del este de Hidalgo. In Inventarios herpetofaunísticos de México: avances en el conocimiento de su biodiversidad, A. Ramírez–Bautista, L. Canseco–Márquez y F. Mendoza Quijano (eds.). Publicaciones de la Sociedad Herpetológica Mexicana 3, México, D. F. p. 99–109. [ Links ]
Moreno, C. E. y G. Halffter. 2000. Assessing the completeness of bat biodiversity inventories using species accumulation curves. Journal of Applied Ecology 37:149–158. [ Links ]
Moreno, E. C., G. Sánchez–Rojas, J. R. Verdu, C. Numa, M. A. Marcos–García, A. P. Martínez–Falcón, E. Galante y G. Halffter. 2007. Biodiversidad en ambientes agropecuarios semiáridos en la Reserva de la Biosfera Barranca de Metztitlán, México. In Hacia una cultura de la conservación de la diversidad biológica, vol. 6, G. Halffter, S. Guevara y A. Melic (eds.). Sociedad Entomológica Aragonesa (SEA), Zaragoza. p. 97–107. [ Links ]
Morrone, J. J. 2001. Biogeografía de América Latina y el Caribe, Manuales y tesis 3. SEA , Zaragoza. 152 p. [ Links ]
Morrone, J. J. y J. Márquez. 2008. Biodiversity of mexican terrestrial arthropods (Arachnida and Hexapoda): a biogeographical puzzle. Acta Zoológica Mexicana (n.s.) 24:15–41. [ Links ]
Ochoa–Ochoa, L. M. y O. A. Flores–Villela. 2006. Áreas de diversidad y endemismo de la herpetofauna mexicana. Universidad Nacional Autónoma de México / Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, D. F. 211 p. [ Links ]
Parra–Olea, G., M. García–París, J. Hanke y D. B. Wake. 2005a. Two new species of Pseudoeurycea (Caudata: Plethodontidae) from the mountains of northern Oaxaca, Mexico. Copeia 2005:461–469. [ Links ]
Pérez–Ramos, E., L. Sadaña de la Riva y J. A. Campbell. 2000. A new allopatric species of Xenosaurus (Squamata: Xenosauridae) from Guerrero, México. Herpetológica 56:500–506. [ Links ]
Pianka, E. R. 1966. Latitudinal gradients in species diversity: a review of concepts. American Naturalist 100:33–46. [ Links ]
Pineda, E., C. E. Moreno, F. Escobar y G. Halffter. 2005. Frog, bat and dung beetle diversity in the cloud forest and coffee agroecosystems of Veracruz, México. Biological Conservation 19:400–410. [ Links ]
Ramírez–Bautista, A. 1994. Manual y claves ilustradas de los anfibios y reptiles de la región de Chamela, Jalisco, México. Cuadernos 23. Instituto de Biología, UNAM, México, D.F. 127 p. [ Links ]
Ramírez–Bautista. 2004. Diversidad de estrategias reproductivas en un ensamble de lagartijas de una región tropical estacional de las costas del Pacifico de México. Boletín de la Sociedad Herpetológica Mexicana 12:7–16. [ Links ]
Ramírez–Bautista, A. y C. Moreno. 2006. Análisis comparativo de la herpetofauna de cuatro regiones geográficas de México. In Inventarios herpetofaunísticos de México: avances en el conocimiento de su biodiversidad 3, A. Ramírez–Bautista, L. Canseco–Márquez y F. Mendoza–Quijano (eds.), Publicaciones Sociedad Herpetológica Mexicana, México, D.F. 74–98. [ Links ]
Ramírez–Bautista A., L. Canseco–Márquez y F. Mendoza Quijano (eds.). 2006a. Inventarios herpetofaunísticos de México: avances en el conocimiento de su biodiversidad, Publicaciones Especiales 3. Sociedad Herpetológica Mexicana, México, D. F. 346 p. [ Links ]
Ramírez–Bautista, A., R. García–Collazo y L. J. Guillette Jr. 2006b. Reproductive, fat, and liver cycles of male and female rose–bellied lizards, Sceloporus variabilis, from coastal areas of southern Veracruz, Mexico. Southwestern Naturalist 51:163–171. [ Links ]
Ramírez–Pérez, A. 2008. Herpetofauna del Parque Nacional El Chico y su zona de influencia, Hidalgo, México. Tesis, Biología, Universidad Autónoma del Estado de Hidalgo. 100 p. [ Links ]
Rossman, D. A., N. B. Ford y R. A. Seigel. 1996. The garter snakes, evolution and ecology. University of Oklahoma, Norman. 332 p. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa –Wiley, México, D.F. 432 p. [ Links ]
Sánchez, O. 1999. Biodiversidad, conservación y manejo de vida silvestre. In Diplomado en manejo de vida silvestre, Conservación y manejo de vertebrados del norte árido y semiárido de México, O. Sánchez y E. Vázquez–Domínguez (eds.). CONABIO / DGVS–INE (SEMARNAP)/ U. S. Fish & Wildlife Service/ Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León, Monterrey, 247 p. [ Links ]
Shine. R. y J. J. Bull. 1979. The evolution of live–bearing in lizards and snakes. Western North American Naturalist.113:905–923. [ Links ]
Smith, H. M. y R. B. Smith. 1976a. Synopsis of the herpetofauna of Mexico. vol. III, Source analysis and index for Mexican reptiles. Johnson, North Bennington, Vermont. 997 p. [ Links ]
Smith, H. M. y R. B. Smith. 1976b. Synopsis of the herpetofauna of Mexico. vol, IV, Source analysis and index for Mexican amphibians. Johnson, North Bennington, Vermont. 260 p. [ Links ]
Smith, H. M. y R. B. Smith. 1993. Sinopsis of the herpetofauna of Mexico, vol. VII (Bibliographic addendum IV and index, bibliographic addenda II–IV, 1979–1991). University of Colorado, Boulder. 1082 p. [ Links ]
Smith, H. M. y E. H. Taylor. 1966. Annotated checklist and keys to the amphibians and reptiles. A reprint of bulletins 187, 194 y 199. Eric Lundberg, Ashton, Maryland. 429 p. [ Links ]
Southwood, T. R. E. y P. A. Henderson. 2000. Ecological methods. Blackwell Science. Oxford. 592 p. [ Links ]
Tinkle, D. W., H. W. Wilbur y S. G. Tilley. 1970. Evolutionary strategies in lizard reproduction. Evolution 24:55–74. [ Links ]
Urbina–Cardona, J. N. y V. H. Reynoso. 2005. Recambio de anfibios y reptiles en el gradiente potrero–borde–interior de la selva en la Reserva de Los Tuxtlas, Veracruz, México. In Sobre diversidad biológica: el significado de las diversidades alfa, beta y gamma, G. Halffter, J. Soberón, P. Koleff y A. Melic (eds.). Monografías 3er Milenio, CONABIO/ CONACYT, México, D.F. /SEA / Diversitas, Zaragoza. p 191–208. [ Links ]
Uribe–Peña Z., A. Ramírez–Bautista y G. Casas–Andreu. 1999. Anfibios y reptiles de las serranías del Distrito Federal, Cuadernos 32, Instituto de Biología, UNAM, México, D.F. 119 p. [ Links ]
Vargas–Contreras, J. A. y A. Hernández–Huerta. 2001. Distribución altitudinal de la mastofauna en la Reserva de la Biosfera "El Cielo", Tamaulipas, México. Acta Zoológica Mexicana (n.s.) 82:83–109. [ Links ]
Vellend, M. 2001. Do commonly–used indices of beta diversity meassures species turnover?. Journal of Vegetation Science 12:545–552. [ Links ]
Vilá. M. 1998. Efectos de la diversidad de especies en el funcionamiento de los ecosistemas. Orsis 13:105–117. [ Links ]
Wyman, R. L. 1990. What's happening to the amphibians?. Conservation Biological 4:350–352. [ Links ]
Zamudio, R., J. Rzedowski, G. Carranza y G. Calderón de Rzedowski. 1992. La vegetación del estado de Querétaro. Instituto de Ecología, Pátzcuaro, Michoacán. 92 p. [ Links ]
Zug, G. R., L. J. Vitt y J. P. Caldwell. 2001. Herpetology: An introductory biology of amphibians and reptiles, second edition. Academic, New York. 630 p. [ Links ]

