










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de biodiversidad
versión On-line ISSN 2007-8706versión impresa ISSN 1870-3453
Rev. Mex. Biodiv. vol.81 no.2 México ago. 2010
Anatomía
Functional morphology of two Lepismium species (Rhipsalideae, Cactaceae)
Morfología funcional de dos especies de Lepismium (Rhipsalideae, Cactaceae)
María Regina Torres–Boeger1*, Patricia Soffiatti1, Marco Antônio Gomes–Souto2, Márcia Budchen2, Katiane Paula Bagatini1 and Manuela Dal Forno2
1 Universidade Federal do Paraná, SCB, Departamento de Botânica, Programa de Pós–Graduação em Botánica, Caixa Postal 19031, CEP 81.531.990 Curitiba, PR, Brazil.
2 Universidade Federal do Paraná, SCB, Departamento de Botânica, Programa de Pós–Graduação em Ecología e Conservação, Caixa Postal 19031, CEP 81.531.990 Curitiba, PR, Brazil.
Correspondent:
rboeger@ufpr.br
Recibido: 16 junio 2009
Aceptado: 17 septiembre 2009
Abstract
The morphology and anatomy of stem segments of 2 species of Lepismium (Cactaceae), which grow naturally in the Araucaria forest understory, in the state of Paraná, Brazil, are compared. The goal of this study was to identify morphological traits adapted to epiphytism and to the low light condition of the studied environment. Twenty–five segments of Lepismium cruciforme and L. lumbricoides were collected and various morphological and anatomical features were measured. Differences (p < 0.05) were found between the species in mean values for total volume, total photosynthetic area, epidermis and hypodermis thickness, sclerenchyma area/total transversal area proportion of the stem segments and parenchyma area/total transversal area proportion, which can be correlated to their differences in shape. The xeric features found in Lepismium, most of them typical of drought–adapted cacti, have allowed the development of the epiphytic habit and the occupation of humid forests. As epiphytes, they are subject to some extent to water scarcity, although not to severe conditions such as most terrestrial cacti. Mean values for hypodermis thickness, water content, amount of sclerenchyma and stomatal densities are more similar to mesomorphic plants, and can be correlated to epiphytism, showing that these species have lost some of their adaptations to dry environments.
Key words: stem morphology, stem anatomy, epiphytic cacti, Araucaria Forest.
Resumen
Se compara la morfología y la anatomía de los segmentos del tallo de 2 especies de Lepismium, que crecen naturalmente en el sotobosque del bosque de Araucaria, en el estado de Paraná. El objetivo de este estudio fue identificar rasgos morfológicos de adaptación para el epifitismo y para condiciones de poca luz del ambiente estudiado. Se recolectaron 25 ejemplares de L. cruciforme y de L. lumbricoides y se midieron rasgos tanto morfológicos como anatómicos de los tallos. Se encontraron diferencias (p < 0.05) entre las especies considerando los valores promedio del volumen total, del área total fotosintética, del grosor de la epidermis y de la hipodermis, del área del esclerénquima / área transversal total de los segmentos del tallo y del área del parénquima / área transversal total, caracteres que se correlacionan a sus diferencias en forma. Las características xeromórficas encontradas en Lepismium, la mayor parte de ellas típicas de cactáceas de zonas áridas, han favorecido el desarrollo del hábito epifítico y la ocupación de bosques húmedos. Como epífitas, están sujetas a un cierto grado de escasez de agua, aunque no a las condiciones tan severas como la mayoría de las cactáceas terrestres. Los valores promedio para el grosor de la hipodermis, el contenido de agua, la cantidad de esclerénquima y las densidad estomática son más similares a las plantas mesomórficas, y se pueden correlacionar al epifitismo, demostrando que estas especies perdieron algunas de sus adaptaciones para ambientes secos.
Palabras clave: morfología del tallo, anatomía del tallo, cactáceas epífitas, bosque de Araucaria.
Introduction
Epiphytes, as a general rule, show physiological and structural characteristics that constitute adaptive advantages in an environment in which water is the greatest constraint (Zotz and Hietz, 2001), besides the lack of nutrients (Lüttge 1989). Common adaptations among these plants are the development of succulence and crassulacean acid metabolism (Zotz and Hietz, 2001). Other adaptations are the velamen in the roots of orchids, and water tanks and specialized trichomes in bromelias (Andrade and Nobel, 1997), which allow water and nutrient uptake. Besides water and nutrient shortage, light intensity should also be a limiting factor to epiphytes growing in the understory of Atlantic forests (Lüttge, 1989).
It is estimated that there are around 20 000 to 25 000 epiphyte species (Benzing, 1990) occurring mostly in tropical forests. Orchidaceae, Bromeliaceae, and Cactaceae are among the dominant angiosperm families with epiphytic species. Out of the 2 000 Cactaceae species, around 10% (220 species) show epiphytic habits, occurring in tropical and subtropical humid forests (Bauer and Waechter, 2006). Floristic surveys in southern Brazil show the occurrence of 6 epiphytic Cactaceae species in gallery forests (Giongo and Waechter, 2004), and 9 species along the coast, on isolated fig plants (Goncalves and Waechter, 2003). In the State of Paraná, 9 species are mentioned occurring in 14 fragments of Araucaria Forest (Borgo and Silva, 2003), where the most common species belong to genera Rhipsalis and Lepismium.
Epiphytic cacti are xeromorphic plants, as are most of the family members, and are characterized by the absence of leaves, presence of areoles, and crassulacean acid metabolism (CAM), and by usually are rooting in humus on tree branches and rocks. However, it has been reported that these species have lost some of their desert adaptations and their stems are thinner and lighter (Mauseth et al., 2002), less succulent, and in some forms, they became flatter, resembling a leaf (Luttge, 1989).
Lepismium belongs to tribe Rhipsalideae, the second largest group of cacti in Brazil (Taylor and Zappi, 2004), and typically grows on tree branches, with long segmented stems. Rhipsalideae, together with Hylocereeae, represent the 2 groups of epiphytic cacti (Anderson, 2001). Lepismium cruciforme (Vell.) Miq. occurs in Argentina, Paraguay and Brazil, with a distribution from Pernambuco to Rio Grande do Sul states. It is present in most forest formations, except for savannas. The stem segments are predominantly flat with some 3–angled parts. Lepismium lumbricoides (Lem.) Barthlott is present in Argentina, Bolivia, Uruguay, and Brazil, where it occurs from Sao Paulo to Rio Grande do Sul states. It is widespread in most forest formations, although rare in the Dense Atlantic Forest. The ribbed segmented stems are cylindrical when young, but larger stems may begin to produce flat segments (Bauer and Waechter, 2006). Both species have immersed areoles, but the former is naked, while the latter has bristles (Bauer and Waechter, 2006).
The goal of this work was to identify morphological traits of Lepismium cruciforme and L. lumbricoides adapted to epiphytism and to the low light condition in the Araucaria forest understory.
Materials and methods
Study area. Samples were collected in a fragment of Araucaria Forest, located at the "Centro Politécnico" of the Federal University of Paraná, in the eastern region of Curitiba, Paraná State, Brazil (25°25'S e 49°17'W). The area has around 2.5 ha, and is located at 900 m altitude (Hirai et al., 1998). The predominant soils of the area are cambisoil and podzolic soil, and the relief is classified as softly wavy (Neto et al., 2002). The study area has 2 well–defined tree strata, with a shaded understory.
The weather of the region is mesothermic, humid and subtropical with cool summers, and winters with frequent frost and no dry season. It is classified as Cfb, following Köppen's classification. According to data from SIMEPAR (Instituto Tecnológico do Paraná, PR) for 2007, the mean annual temperature is 17 oC, and the mean annual temperatures in the hottest and coldest months are around 22 and 18 oC, respectively. The mean relative air humidity is around 85 % and the annual precipitation varies between 1 300 and 1 500 mm, with the water index varying between 60 and 100, without water deficiency along the year.
Twenty–five specimens from each species were collected with clipper poles. One complete individual was considered as a specimen that posses a stem which hangs from a rooted horizontal axis on the tree branch. The samples were kept in plastic bags and taken to the laboratory for the analysis.
Morphological study. For each individual, the longest stem, composed of a set of branches, was used in this study. Stem length was measured with a metric tape. Thickness and width and/or diameter were measured with a digital caliper at the very base. Afterwards, each stem was weighted on an analytical balance to determine fresh weight (FW) and dry weight (DW) after drying for 48 hours at 50oC in an oven. Water content was estimated as the difference between FW and DW. The stem area (cm2) of L. cruciforme was estimated using a scanned digitized image, generated by the program SIGMASCAN–PRO (v. 5, SPSS Inc. Chicago, USA). Total volume for the species was calculated from these data. For L. cruciforme area was calculated as (ALC) = stem greatest width*stem smallest width/2, and the volume as (VLC) = ALC * total length. For L. lumbricoides, the area was calculated as (ALL) = 2*π*(1/2 diameter)2, and the volume as (VLC) = ALL* stem length. For calculating sclerenchyma area/ total area, we considered all lignified tissue, including sclereid caps below the hypodermis, scattered sclereids, sclereid caps outside the vascular cylinder and the xylem cylinder. Stomatal density (number/mm2) was measured from nail polish epidermal peels. For each species, 75 sites were counted, 25 from the apical, 25 from the median and 25 from the basal portion of the stem. The same procedure was used for areole density.
Anatomical study. Transverse sections of fresh living material of 10 individuals were made using free–hand sectioning, and were taken from the middle of the stem, cleared by sodium hypochlorite, stained with 0.05 % toluidine blue (Feder and O'Brien, 1968) and mounted in Entelan®. The starch was identified using Lugol and cuticle using Sudan III (Johansen, 1940). Transverse sections of 7 μηι thick obtained with a rotary microtome were also made, from samples dehydrated through an ethanol series and embedded in Leica® synthetic resin, according to the manufacturer's instructions, stained with 0.05 % toluidine blue (Feder and O'Brien, 1968) and mounted in Entelan®.
The following measurements were obtained using the preparations: cuticle, epidermis, and hypodermis thickness; and sclerenchyma and parenchyma tissue percentage. Sclerenchyma/parenchyma tissue percentage was estimated using SIGMASCAN–PRO software (4.0 version, SPSS Inc., Chicago Illinois, USA). Descriptions and measurements were made using bright–field microscopy and an optical scale.
For all quantitative values, means and standard deviations were calculated. Comparisons of the quantitative means between species and comparisons of stomata density among sites in the same species were made through a t–test and ANOVA, respectively, with 5 % significance, using STATISTICA (6.0 version, StatSoft Inc., USA). All quantitative data are given in Table 1.
Results
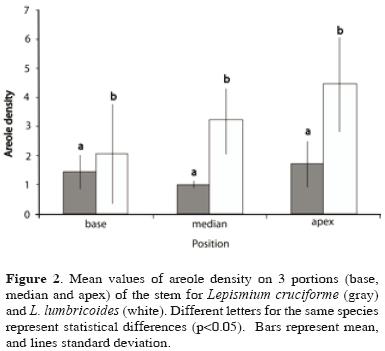
Morphological description. Lepismium cruciforme has a flat stem 12.65 (± 2.42) mm wide, 2.67 (±0.56) mm thick, and 108.7 (±36.31) mm in length, which is not ribbed though 3–angled in some parts. The total volume of the stem is 12. 27 (±5.39) cm3 and the total photosynthetic area is 143.70 (± 59.48) cm2. The stem contains 80.25 (±1.97) % water content. Lepismium lumbricoides has 6.9 (±0.88) ribs in the segmented cylindrical stem with 3.12 (±0.33) mm diameter and 113.98 (±32.56) mm in length. The total volume of the stem is 8.48 (±2.77) cm3 and the total photosynthetic area is 106.18 (± 32.82) cm2. The stem contains 80.08 (±1.95) % water content. Among these characteristics, only total volume and total photosynthetic area were significantly higher (p<0.05) in L. cruciforme. Sclerenchyma area/total transversal area proportion (4.76±2.2; 10.51±5.10) and parenchyma area/transversal area proportion (95.53±2.20; 89.81±4.95) are significantly different (p<0.05) between L. cruciforme and L. lumbricoides, respectively. Stomata density differs between the base segment and the other segments (Fig. 1) but not between species (Table 1) and the numbers of areoles differs among segments of stem only in L. lumbricoides (Fig. 2) and between species (Table 1).

Anatomical description. Lepismium cruciforme: the stem has a uniseriate epidermis (29.97±6.63 μm) (Figs. 3, 4), composed of irregular cells, varying from rectangular to triangular, with thick external periclinal walls and thinner anticlinal ones in transverse view, covered by a cuticle (10.73±2.73 μm) (Figs. 3, 4). Stomata are parallelocytic and at the same level as the other epidermal cells (Fig. 3). Internal to the epidermis, there is a hypodermis composed of 1 layer of collenchymatic cells (75.33±10.55 μm) of irregular size (Figs. 3, 4), with thicker walls than adjacent parenchyma cells (Fig. 4). The substomatal chambers cross hypodermis. The cortex is composed mainly of isodiametric parenchyma cells with thin walls (Figs. 3, 4). Sclereids, mucilaginous cells, and prismatic crystals are found dispersed in the cortex. Isolated and scattered vascular bundles occur in the inner areas of the cortex. Chloroplasts and starch occur through the entire cortex and pith. Fiber caps are present around the vascular cylinder, forming an incomplete ring (Fig. 5).
Lepismium lumbricoides: the stem has a uniseriate epidermis (11.15±6.18 μm), composed of irregular cells, varying from rectangular to triangular, with thick periclinal walls in transverse sections, covered by a thick cuticle (11.37±1.15 μm) (Figs. 6, 7). Stomata are parallelocytic and located slightly below epidermal cells level (Fig. 7). Internal to the epidermis is a hypodermis composed of 1 layer of collenchymatous cells (22.62±3.33 μm) of irregular sizes, with thicker walls than adjacent parenchyma cells (Fig. 7). The substomatal chambers cross the hypodermis. The cortex is composed mainly of isodiametric parenchyma cells with thin walls (Fig. 6). Fiber bundles form a non–continuous ring below hypodermis (Fig. 6). Isolated and scattered vascular bundles occur in the inner areas of the cortex. Mucilage cells are dispersed in the cortex. Chloroplasts and starch occur through the entire cortex and pith. Fiber caps are present around the vascular cylinder (Fig. 8).
Discussion
In this study, most of the characters observed in these Lepismium species corroborate the pattern found in Cactaceae. The uniseriate epidermis covered by a cuticle, presence of a hypodermis, a chlorophyllous cortical parenchyma, and cortical bundles, were already described for other species of cacti (Terrazas and Mauseth, 2002; Soffiatti and Angyalossy, 2007; Calvente et al., 2008).
The cuticle has an important function in protection, being the primary barrier between the plant body and the environment. In cacti, especially on those species occurring in very arid habitats, external walls of epidermal cells are highly cutinized (Mauseth et al., 1998; Loza–Cornejo and Terrazas, 2003). However, it is a feature influenced by environmental conditions and can have a lot of variation in thickness in Cactoideae (Gibson and Horak, 1978; Mauseth, 1996; Nyffeler and Eggli, 1997; Mauseth et al., 1998; Loza–Cornejo and Terrazas, 2003; Terrazas and Arias, 2003; Mauseth, 2006). According to Terrazas and Arias (2003), cuticle thickness may vary between 1–25 μm thick in Cactoideae; on the other hand, Mauseth (2006) mentions that cuticle in cacti are generally around 1.0–10 μm thick, which is corroborated by Terrazas et al. (2005) in Stenocereus, a genus of Cactoideae, and Terrazas and Loza–Cornejo (2003) for Cactoideae as a whole, although the latter observed it can reach up to 200 μm in a species of Ariocarpus, a tiny globose cacti. Cuticle of both species of Lepismium (< 12 μm, Table 1) is considered thin for a xerophyte (Mauseth, 2006). Even considering the low light conditions these species are subject to, their cuticle thickness corroborates with nonepiphytic cacti in general.
The epidermis of both species is uniseriate, common in the great majority of cacti (Gibson and Nobel, 1986; Nyffeler and Eggli, 1997; Terrazas and Arias, 2003). Lepismium cruciforme showed significantly higher values of thickness of the epidermis as well as the hypodermis compared to L. lumbricoides (Table 1). The number of layers and thickness of hypodermis are highly variable in cacti (Terrazas and Arias, 2003; Mauseth, 2006). In Cactaceae, the epidermis and hypodermis play important roles giving support to the succulent stem by preventing water loss and protecting inner tissues against sunlight, in addition to providing defense against microscopic organisms (Gibson and Nobel, 1986). Gibson and Nobel (1986) showed that the collenchymatic hypodermis found in most Cactaceae gives flexibility allowing the contraction or expansion of the stem according to the water availability. Both studied species have an uniseriate hypodermis, composed of 1 layer of thick–walled collenchyma cells when compared to the surrounding parenchyma. In L. cruciforme, this layer is thicker when compared to L. lumbricoides, where it is the only supporting tissue present close to surface. Overall, both species here studied have a thin hypodermis, compared with other cacti (Terrazas et al., 2005; Mauseth, 2006; Soffiatti and Angyalossy, 2007). Calvente et al. (2008) mentioned the presence of either thick– or thin–walled hypodermal cells in Rhipsalis species. The hypodermis also plays a role in maintaining rib integrity (Gibson and Nobel, 1986). However in the studied species, ribs are absent or small and in low number, which could explain the presence of a not very well developed hypodermis in both species. In addition to the collenchymatic hypodermis, there are caps of sclereids forming an almost continuous band below the hypodermis in L. lumbricoides, which certainly play an additional role in support due to its cylindrical shape. The presence of sclereid caps is reflected in the higher percentage of sclerenchyma area in L. lumbricoides (Table 1). Both species present caps of sclereids with very thick–walls around the vascular cylinder.
The stomata density differs along the stem in both species increasing significantly toward the stem apex (Fig. 1, Table 1), which corroborates the study of Hernández et al. (2007). According to these authors, this difference in density along the stem has to do with epidermal cells maturation toward the base. The stomata density in both Lepismium is 6–11 stomata.mm2, which is considered low when compared to other Cactoideae species (20–80 st.mm–2, Terrazas and Mauseth, 2002). The values of stomata density are already considered low when compared to what is found in leaves of other C3 and C4 plants (Terrazas and Mauseth, 2002). The results here also do not agree with Calvente et al. (2008) who studied Rhipsalis species. In the Lepismium species, the low stomata density may be related to the relatively high humidity of the air, associated with the high local precipitation, which allows the stomata to stay open longer without facing the typical desiccation of xeric environments The stomata type is parallelocytic, a common type of stomata found in Cactaceae (Terrazas and Mauseth, 2002; Terrazas and Arias, 2003; Hernández et al., 2007; Calvente et al., 2008).
A chlorenchymatous palisade parenchyma was not observed in either species, as previously described for most Cactoideae species (Terrazas and Arias, 2003). It corroborates with what was found by Calvente et al. (2008), in their anatomical study of 5 Rhipsalis species. The presence of a homogenous chlorophyll parenchyma may be related to the amount of incident light in the occurring area of the species, which is more shaded than the xeric environments where Cactaceae species are commonly found. The shape of the homogenous parenchyma cells, associated with intercellular spaces, could, hypothetically, reflect and refract the light, increasing the chances of absorption (Taiz and Zeiger, 2004). As a consequence, this parenchyma seems to be more efficient in diffuse light distribution in the interior of the stem than the palisade parenchyma cells (Vogelmann et al., 1996) and the higher number of homogenous parenchyma layers increases even more this efficiency; this could mean that their pendent stems facilitate light capture (Mauseth et al., 2002).
Spines act not only protecting against herbivores but also against insolation (Gibson and Nobel, 1986; Mauseth, 2006). The lack of spines helps in maximizing photosynthetically active radiation interception, and it is very common in epiphytes, where they are reduced or completely lost in an environment of low light (Gibson and Nobel, 1986), which is seen in Lepismium species, where L. cruciforme completely lacks them and L. lumbricoides has only bristles.
Ribs are another striking character present in several species of Cactaceae, important in the process of adjusting the volume/surface rate of succulent stems according to water availability. The expansivity of the stems depends on the number and size of the ribs (Mauseth, 2000) and it is primarily important in environments of extreme drought. As ribs are mainly related to light interception, in general epiphytic cacti have smaller numbers of ribs (Gibson and Nobel, 1986), which is corroborated in the present study. Lespimium lumbricoides has a small number of small ribs (less than 8), which indicates a moderate capacity of water retention. Lespimium cruciforme has no ribs at all. The absence or small number of ribs may be a selected character in epiphyte cacti (Gibson and Nobel, 1986). Associated to the short dry season in the Araucaria forest and high air humidity, these plants do not need to adjust their stems as there are no strong water availability fluctuations.
Water content in Lespimium was 80 %, similar in both species and lower than other cacti (Rizzini, 1987). According to Zotz & Hietz (2001), water deficiency is the most important abiotic restriction for epiphytes, in comparison to the scarce availability of nutrients. Although living in humid forests, they are also subject to a certain degree of drought, as they are rooted over tree barks, which do not hold water for very long. Therefore, succulence is an important trait for the maintenance of water storage, even inside a tropical forest. In the studied species, the water–storage is performed by the large isodiametric parenchyma cells of the cortex. The fact that there is relatively little water–storage tissue in these species is reflected by the lower water content compared to other cacti, which means drought is not as limiting as light in this kind of environment. However, their water content is higher than any other eudicotyledons (Boeger et al., 2006), indicating that they are still succulents.
The species differed significantly regarding mean values of surface and volume, total photosynthetic area, sclerenchyma area/total transversal area proportion, and parenchyma area/total transversal area proportion. Lepismium cruciforme shows higher values of surface, volume and total photosynthetic area, probably due to the flat shape of its stems compared to the cylindrical ones of L. lumbricoides. The flat stem of L. cruciforme may be an additional advantage enabling greater light capture in shaded environments because of its more exposed surface (Mauseth, 2000), resembling a large leaf which maximize light interception. However, the ratio between these 2 attributes (total volume/total surface) is similar between species, indicating that they have similar surface areas per unit volume, independently of the shape of the stem. The ratio of volume/surface area is relatively high when compared to the mean values (0.169) found for 200 Cactaceae species of different environments (Mauseth, 2000). The values found in the studied Lepismium species are, probably, the result of the size of the stem, which are much longer than wide (length/width rate of L. cruciforme = 86.26; length/width rate of L. lumbricoides = 365.32). On the other hand, the sclerenchyma tissue area/total transversal area proportion and parenchyma area/total transversal area proportion are higher in L. lumbricoides, which has a cylindrical stem, where more supporting tissue is needed. Overall, because of their slender and pendent stems, they do not need much supporting tissue.
Considering the set of analyzed morphological attributes, the studied species of Lepismum show some characteristics that were selected by the environmental conditions, despite the historical xeromorphic component of the group. These species became thinner and lighter, and lost some desert–adapted characters, such as large storing tissues and ribs which are no longer necessary in an environment with plenty of rainfall and low light. The more striking characteristics in these species are the presence of a homogenous parenchyma instead the cortical palisade parenchyma and the poorly developed hypodermis. Because of the epiphytic conditions, it was expected that some xeromorphic characteristics were more evident, such as higher stomata density, higher succulence and better developed sclerenchyma tissues, besides the above mentioned characteristics. However, the mean values for hypodermis thickness, water content, amount of sclerenchyma, and stomata density are more similar to those of mesomorphic plants, which suggest the influence of the mesic environment conditions in the structure of the stems. Additional studies under different environmental conditions, such as light and water availability may elucidate the degree of morphological plasticity in the analyzed characteristics and may help the understanding of adaptive strategies of epiphytic Cactaceae species. The xeric features found in Lepismium, most of them typical of drought–adapted cacti, have favored the development of epyphitic habit and the occupation of humid forests. Although they live in a rainy environments, as epiphytes they do not have immediate access to water absorption, being also subject to water deficit even living inside a rain forest.
Literature cited
Anderson, E. F. 2001. The cactus family. Timber Press, Portland, Oregon. 776 p. [ Links ]
Andrade, J. L. and P. S. Nobel. 1997. Microhabitats water relations of epiphytic cacti ferns in a low neotropical forest. Biotropica 29:261–270. [ Links ]
Bauer, D. and J. L.Waechter. 2006. Sinopse taxonómica de Cactaceae epifíticas no Rio Grande do Sul, Brasil. Acta Botanica Brasilica 20:225–239. [ Links ]
Benzing, D. H. 1990. Vascular epiphytes: general biology and related biota Cambridge University Press, Cambridge. 354 p. [ Links ]
Boeger, M. R. T., M. Kaehler, J. C. F. Melo Júnior, M. Z. Gomes, L. S. Oliveira, C. R. M. Chaves and E. S. Schottz. 2006. Estrutura foliar de seis espécies do subosque de um remanescente de Floresta Ombrófila Mista. Hoehnea 33:521–531. [ Links ]
Borgo, M and S. M. Silva. 2003. Epífitos vasculares em fragmentos de Floresta Ombrófila Mista, Curitiba, Paraná, Brasil. Revista Brasileira de Botânica 26:391–401. [ Links ]
Calvente, A. M.; E. R. H. P. Andreata and R. C.Vieira. 2008. Stem anatomy of Rhipsalis (Cactaceae) and its relevance for taxonomy. Plant Systematic and Evolution 276:271–277. [ Links ]
Feder, N. and T. P. O'Brien. 1968. Plant microtechnique: some principles new methods. American Journal of Botany 55:123–142. [ Links ]
Giongo, C. and J. L.Waechter. 2004. Composição florística e estrutura comunitária de epífitos vasculares em uma floresta de galeria na Depressão Central do Rio Grande do Sul. Revista Brasileira de Botânica 27:563–572. [ Links ]
Gibson A. C. and K. E. Horak.1978. Systematic anatomy and phylogeny of Mexican columnar cacti. Annals of the Missouri Botanical Garden 65:999–1057. [ Links ]
Gibson, A. C. and P. S. Nobel. 1986. The Cactus Primer. Harvad University Press, Cambridge. 286 p. [ Links ]
Gonçalves, C. N. and J. L.Waechter. 2003. Aspectos florísticos e ecológicos de epífitos vasculares sobre figueiras isoladas no norte da planície costeira do Rio Grande do Sul. Acta Botanica Brasilica 17:89–100. [ Links ]
Hernandez, M., T. Terrazas, A. D. Alvarado and M. L. Cavazos. 2007. Los estomas de Myrtillocactus geometrizans (Mart. Ex. Pfeiff.) Console (Cactaceae): variación e su área de distribución. Revista Fitotecnia Mexicana 30:235–240. [ Links ]
Hirai, R. Y., O. Yano and M. E. G. Ribas. 1998. Musgos da mata residual do Centro Politécnico (Capão da Ed. Física), Curitiba, Paraná, Brasil. Boletim do Instituto de Botânica 11:81–118. [ Links ]
Johansen, D. A. 1940. Plant microtechnique. McGraw–Hill Book Co. Inc. New York. 523 p. [ Links ]
Loza–Cornejo, S. and T. Terrazas. 2003. Epidermal and hypodermal characteristics in North American Cactoideae (Cactaceae). Journal of Plant Research 116:27–35. [ Links ]
Lüttge, U. 1989. Vascular epiphytes: setting the scene. In Ecological Studies 79: Vascular plants as ephiphytes. U. Lüttge (ed.). Springer–Verlag. New York. p.1–12. [ Links ]
Mauseth, J. D. 1996. Comparative anatomy of tribes Cereeae and Browningieae (Cactaceae). Bradleya 14:66–81. [ Links ]
Mauseth J. D., T. Terrazas and S. Loza–Cornejo. 1998. Anatomy of relictual members of subfamily Cactoideae, IOS Group 1a (Cactaceae). Bradleya 16:31–43. [ Links ]
Mauseth, J. D. 2000. Theorethical aspects of surface–to–volume ratios water storage capacities of succulent shoots. American Journal of Botany 87:1107–1115. [ Links ]
Mauseth J. D., R. Kiesling and C. Ostolaza. 2002. A cactus odyssey: journeys in the wilds of Bolivia, Peru, and Argentina. Portland, OR: Timber Press. 306 p. [ Links ]
Mauseth, J. D. 2006. Structure–function relationships in highly modified shoots of Cactaceae. Annals of Botany 98:901–926. [ Links ]
Mauseth, J. D. 2006. Structure–function relationships in highly modified shoots of Cactaceae. Annals of Botany 98:901–926. [ Links ]
Neto, R. M. R., C. Kozera, R. do R. de Andrade, A. T. Cecy, A. P. Hummes, E. Fritzsons, M. V. W. Caldeira, M. de N. M. Maciel and M. K. F. de Souza. 2002. Caracterizaçao florística e estrutural de um fragmento de Floresta Ombrófila Mista, em Curitiba, PR–Brasil. Revista Floresta 32:3–16. [ Links ]
Nyffeler, R. and E. Eggli. 1997. Comparative stem anatomy systematic of Eriosyce sensu lato (Cactaceae). Annals of Botany 80:767–786. [ Links ]
Rizzini, C. T. 1987. Cactáceas – Os segredos da sobrevivência. Ciência Hoje 5:30. [ Links ]
Soffiatti, P. and V. Angyalossy. 2007. Anatomy of brazilian Cereeae (subfamily Cactoideae, Cactaceae): Arrojadoa Britton & Rose, Stephanocereus A. Berger and Brasilicereus Backeberg. Acta Botânica Brasilica 21:813–822. [ Links ]
Taylor, N. P. and D. C. Zappi. 2004. Cacti of Eastern Brazil. Royal Botanic Gardens. Kew. UK. 499 p. [ Links ]
Taiz, L. and E. Zeiger. 2004. Fisiologia Vegetal. 3°. ed. Porto Alegre: Artmed. 720 p. [ Links ]
Terrazas, T. and J. D. Mauseth. 2002. Chapter 2. Shoot anatomy and morphology. In Cacti: biology and uses, P. S. Nobel (ed.). University of California Press, Berkeley. p. 23–40. [ Links ]
Terrazas, T. and S. Arias. 2003. Comparative stem anatomy in the subfamily Cactoideae. The Botanical Review 68:444–473. [ Links ]
Terrazas, T., S. Loza–Cornejo, and H. J. Arreola–Nava. 2005. Anatomía caulinar de las especies del género Stenocereus (Cactaceae). Acta Botanica Venezuelica 28:321–336. [ Links ]
Vogelmann, T. C., J. N. Nishio and W. K. Smith. 1996. Leaves light capture: light propagation gradients of carbon fixation within leaves. Trends in Plant Science 1:65–70. [ Links ]
Zotz, G. and P. Hietz. 2001. The physiological ecology of vascular epiphytes: current knowledge, open questions. Journal of Experimental Botany 52:2067–2078. [ Links ]

