










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de biodiversidad
versión On-line ISSN 2007-8706versión impresa ISSN 1870-3453
Rev. Mex. Biodiv. vol.78 no.1 México jun. 2007
Ecología
Caracterización de sitios de percha del guajolote silvestre (Meleagris gallopavo mexicana) en Sierra Fría, Aguascalientes, México
Roost sites characteristics of wild turkey (Meleagris gallopavo mexicana) in Sierra Fria, Aguascalientes, Mexico
Marcelo Márquez–Olivas1*, Edmundo García–Moya1, Carlos González–Rebeles Islas2 y Humberto Vaquera–Huerta3
1 Programa de Botánica, Colegio de Postgraduados, Carretera México–Texcoco, km 36.5, Montecillo, 56230 Estado de México, México.
2 Departamento de Etología, Fauna Silvestre y Animales de Laboratorio, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México. 04510 México, D. F., México.
3Posgrado en Estadística, Campus Montecillo, Colegio de Postgraduados, Carretera México–Texcoco, km 36.5, Montecillo, 56230 Estado de México, México.
* Correspondencia: molivas@colpos.mx
Recibido: 21 noviembre 2005
Aceptado: 03 agosto 2006
Resumen
Entre octubre y noviembre del año 2001 y mayo de 2002 se localizaron 16 sitios de percha de guajolote silvestre (Meleagris gallopavo mexicana) en la Sierra Fría de Aguascalientes. El 87.5% de los sitios se encontraron en bosques de pino–encino (en rodales de pinos), en altitudes de 2500 a 2650 m. En la parte alta (2/3) de la cuesta se localizaron 8 de los sitios y 7 sobre laderas de exposición noreste, en una pendiente promedio de 38.9 ± 12.6%. El 62.5% de los sitios se usaron durante todo el periodo de estudio y el resto, de manera ocasional. La mayoría de los dormideros se encontraron cerca de comederos o cebaderos y de parcelas agrícolas. Se contabilizaron de 1 a 23 árboles de percha utilizados por sitio, distribuidos a una distancia promedio de 24.4 ± 17.8 metros. El 81.2% de los árboles (n=170) fueron pinos (Pinus teocote, P. durangensis, P. leiophylla y P. michoacana), 17.6% encinos (Quercus laurina, Q. grisea y Q. eduardii) y 1.2% madroños (Arbutus arizonica). La altura promedio de los árboles fue de 16.6 ± 4.4 m y el diámetro 43.2 ± 11.5 cm. La preferencia del guajolote silvestre por utilizar pinos como dormideros en lugar de otras especies arbóreas, se debe posiblemente a que son árboles de mayor porte o a la conformación de su dosel; sin embargo, se ha observado que en lugares donde hay pocos pinos, los guajolotes utilizan para dormir otros árboles de características semejantes, como encinos, madroños o sicomoros.
Palabras clave: especie protegida, ave cinegética, hábitat, Sierra Madre Occidental.
Abstract
Sixteen roosting sites of Gould's wild turkey (Meleagris gallopavo mexicana) were found between October and November of 2001 and May of 2002 in Sierra Fría, Aguascalientes. A large percentage (87.5%) of the roost sites was located in oak–pine forest (in pine stands), at an elevation range of 2500 to 2650 meters. Eight roost sites were on the upper portion (2/3) of the ridge, and 7 at northeast exposures with an average slope of 38.9 ± 12.6%. Most (81.2%) of the roost trees (n =170) were pines (Pinus teocote, P. durangensis, P. leiophylla and P. michoacana), 17.6% oaks (Quercus laurina, Q. grisea, and Q. eduardii), and 1.2% madrones (Arbutus arizonica). More than half (62.5%) of the roosting sites were frequently used during the period of study and the remaining sites were occasionally used. Most of the roost sites were near bait sites and small agricultural areas. The number of roost trees found at roosting sites varied from 1 to 23. The average distance within roost trees was of 24.4 ± 17.8 m. The average roost tree height was 16.6 ± 4.4 m and the average diameter at breast height (dbh) was 43.2 ± 11.5 cm. The selection of pines as roost trees by wild turkey may be due to tree structure. However, in places without pines, wild turkey used oaks, madrones and sycamore with similar characteristics of pine for roosting.
Key words: protected species, game bird, habitat, Sierra Madre Occidental
Introducción
El guajolote silvestre (Meleagris gallopavo L.) es una especie protegida por la Norma Oficial Mexicana NOM059–ECOL–2001 (Diario Oficial de la Federación, 2002), pero en las localidades donde es abundante se permite su aprovechamiento cinegético bajo un plan de manejo porque es muy atractivo para los cazadores, ya que es la gallinácea de mayor talla y el rey de las aves de caza de Norteamérica (Leopold, 1977; Eaton, 1992). Debido a su tamaño, comportamiento y requerimientos ecológicos, dicha especie es muy sensible a las alteraciones climáticas y del hábitat; por ejemplo, las sequías persistentes o la deforestación son factores que con mucha frecuencia inciden de manera negativa en su población (Austin y de Graff, 1975; Wunz y Hayden, 1975). Numerosos estudios indican que las características físico–bióticas de los sitios que los guajolotes prefieren para descansar o dormir se relacionan en buena medida con su seguridad, de tal modo que a estos espacios del medio se les considera como un componente esencial de su hábitat, sobre todo cuando las condiciones del clima no son favorables (Glover, 1948; Bailey y Rinell, 1967; Porter, 1978; Mackey, 1984; Dickson, 1992; Chamberlain et al., 2000).
Los sitios de descanso o percha, y el tamaño y la conformación de los árboles que los guajolotes eligen para dormir, varían durante el año de acuerdo a la disponibilidad de agua, alimento y condiciones meteorológicas (v.gr., temporada de frío, calor, lluvia o viento). La elección de los dormideros depende normalmente de la combinación de las características físicas del terreno y de la vegetación; sin embargo, en términos generales los guajolotes escogen rodales de bosques maduros o sobremaduros y árboles altos, de fácil acceso y con ramas plagiotrópicas o semihorizontales, para descansar (Scott y Boeker, 1977; Good, 1982; Kilpatrick et al., 1988; Zwank et al., 1988; Rumble, 1992).
De las 5 subespecies de guajolote silvestre que habitan en Norteamérica, Meleagris gallopavo mexicana Gould, 1856 es la subespecie de mayor tamaño, pero también una de las menos estudiadas (Kennamer, s/f). Se distribuye casi de manera exclusiva en México, aunque existe una pequeña población en el sur de Arizona y Nuevo México en los Estados Unidos (Lewis, 1973), donde se encuentran amenazadas, por lo que se trabaja para su repoblación (Wakeling et al., 2001).
En México, el guajolote silvestre de Gould habita a lo largo de la Sierra Madre Occidental en los estados de Sonora, Chihuahua, Durango, Sinaloa, Zacatecas, Nayarit y Jalisco (Lafón, 1997), así como en Aguascalientes y Guanajuato donde fue reintroducido en años recientes. Se han hecho varios estudios en los que se describen las características bióticas y abióticas del hábitat que usa esta subespecie en la parte norte de su distribución (York, 1991; Garza y Servín, 1993; Zornes, 1993; Lafón, 1997; Valencia y López, 2000), pero el conocimiento sobre éste en el sur es escaso o inexistente.
En la Sierra Fría de Aguascalientes se realizan de manera temporal actividades silvícolas como la extracción de leña, el pastoreo del ganado, el desmonte de áreas de matorral, la construcción de caminos y brechas, el saneamiento del bosque de árboles plagados, secos o viejos mediante su corte, y la repoblación de pináceas. Algunas de esas ocupaciones podrían tener un impacto negativo en la dinámica de la población del guajolote silvestre y reducir su población (NWF, 1982).
El objetivo de este estudio fue describir las características físicas y bióticas de los sitios que los guajolotes usan para descansar y contribuir en el conocimiento de uno de los requerimientos vitales de esta subespecie que ha sido poco estudiada en México, y de manera muy particular en esa parte de su distribución histórica. La información que se obtenga de esta investigación deberá considerarse dentro de los planes de manejo forestal de Sierra Fría, en beneficio de la conservación del guajolote silvestre y de las condiciones ecológicas de la región.
Área de estudio. La Sierra Fría es la región más importante del estado de Aguascalientes por su biodiversidad y su papel regulador del clima y del ciclo hidrológico; además, contiene vestigios arqueológicos, por lo que se considera no sólo un patrimonio natural sino también cultural (SEDESOL, 1996). Con base en esos atributos, el 30 de enero de 1994, la Sierra Fría fue declarada Área Natural Protegida, bajo la categoría de Zona Sujeta a Conservación Ecológica, (Periódico Oficial, 1994). Dicha área se localiza al noroeste del estado de Aguascalientes, entre los 21° 52' 50'' y 22° 19' 46'' N y 102° 22' 50'' y 102° 51' 26'' O; en la figura 1 se indican las principales vías de acceso por carretera. Tiene una extensión territorial de 112 090 ha, de las cuales de manera aproximada, el 64% es propiedad privada, 23% ejidal y 13% comunal (SEDESOL, 1993).

La Sierra Fría de Aguascalientes forma parte de la Sierra Madre Occidental; sin embargo, se encuentra aislada de esta cordillera por una región seca que caracteriza la zona de transición entre las provincias fisiográficas de la Mesa Central y del Norte (INEGI, 1981). La fisiografía es escarpada y se ubica dentro de la Región Hidrológica Núm. 12, la cual abastece la cuenca del río Lerma–Santiago por medio de los ríos Verde Grande, Tlaltenango, Morcinique y Calvillo, entre otros afluentes. La hidrología superficial de Sierra Fría se compone de bordos y presas permanentes, los cuales se abastecen de los escurrimientos pluviales temporales (INEGI, 1993). Los climas semisecosemicálido, semiseco–templado y templado–subhúmedo, predominan en la región. La temperatura media anual es de 17°C y la precipitación promedio de 600 mm; enero es el mes más frío y julio el más cálido, mientras que marzo es más seco y julio el más húmedo (Villalobos, 1998). La mayor parte de los suelos son someros y con afloramiento rocoso, de color gris rojizo y gris oscuro, con texturas franco arcillosas y un pH que varía de 6 a 6.6, con algunas zonas bastante ácidas (Villalobos, 1998).
Como parte de la biota de Sierra Fría se han identificado 591 especies vegetales terrestres, las cuales pertenecen a 325 géneros y 85 familias; 37 especies acuáticas o subacuáticas que corresponden a 28 géneros y 21 familias, y 95 especies de hongos (Hesselbach, 1996). Los encinares y su asociación con otras especies cubren cerca del 97% de los bosques templados de Sierra Fría, cuya extensión abarcan el 50.8% del área total. Las especies más abundantes son Quercus resinosa Liebm., Q. potosina Trel., Q. eduardii Trel., Q. laeta, Q. grisea Liebm., Q. sideroxyla Humb. et Bonpl. y Q. rugosa Nee., y las menos conspicuas Q. chihuahuensis Trel., Q. aristata Hook. et Arn., Q. coccolobifolia Trel., Q. microphylla Nee., Q. uxoris McVaugh, Q. gentryi C.H. Muller y Q. laurina Humb. et Bonpl. Los pinares ocupan menos del 1% de los bosques templados de la región y por lo general se encuentran en algunas cañadas y en las partes altas de la Sierra; las especies más comunes son Pinus lumholtzii Rob. et Fern., P. leiophylla [Schl. et Cham.] var. chihuahuana Shaw. y P. teocote Schl. et Cham., y las menos frecuentes P. michoacana var. cornuta Mtz., P. durangensis Mtz. y P. cembroides Zucc. (SEDESOL, 1993). Otros árboles y arbustos comunes en Sierra Fría son el táscate (Juniperus deppeana Steud.) y la manzanita (Arctostaphylos pungens HBK.), especies que han proliferado debido a los diferentes disturbios ocurridos en la región y que han favorecido su establecimiento y dispersión.
En la Sierra Fría de Aguascalientes se tiene un registro de 228 especies de vertebrados; 47.8% aves, 38.1% mamíferos y 14% reptiles y anfibios; además, 13 órdenes de insectos que corresponden a 65 familias (Hesselbach, 1996). Las especies cinegéticas de mayor importancia económica son el guajolote silvestre y el venado cola blanca (Odocoileus virginianus). También se permite la caza de jabalí de collar o pecarí (Tayassu tajacu), de coyote (Canis latrans) y otras especies menores, como conejo (Sylvilagus floridanus), paloma (Zenaida asiatica y Z. macroura) y codorniz (Cyrtonyx montezumae).
La atención de la demanda cinegética en la Sierra Fría de Aguascalientes ha requerido la introducción de especies exóticas, como el ciervo rojo (Cervus elaphus), el faisán de collar (Phasianus colchicus) y la perdiz chukar (Alectoris chukar), entre otras, que al no tener el éxito esperado, se han tenido que remover o mantener en confinamiento para exhibición o reproducción.
La mayoría de los habitantes de Sierra Fría subsisten de las actividades productivas tradicionales, como la ganadería extensiva y la agricultura de temporal; sin embargo, un grupo importante participa en el cuidado y protección de los recursos naturales y son promotores del llamado turismo cinegético, el campismo y el ecoturismo de la región.
Materiales y métodos
Entre el 12 de octubre y el 23 de noviembre del año 2001 (periodo de otoño) y del 3 al 19 de mayo del 2002 (periodo de primavera), se hicieron varios recorridos a pie y en vehículo en busca de sitios de percha o dormideros de guajolote silvestre en la Sierra Fría de Aguascalientes. Se eligió el Rancho El Gauro como campamento base, desde donde nos movilizamos de manera aleatoria hasta un máximo de 15 km a su alrededor. Para ubicar los sitios que los guajolotes utilizaron como dormideros, en ocasiones se realizaron recorridos por la noche en vehículo, con la intención de escuchar su gorgoreo o canto, registrar la ubicación, y localizarlos al día siguiente. La mayoría de las veces se hizo una búsqueda exhaustiva (árbol por árbol), durante el día, a pie, en recorridos de 3 km en promedio para detectar acumulación de excretas de los guajolotes en la vegetación arbórea y el suelo. Asimismo, durante el trabajo de campo nos apoyamos en el conocimiento de informantes de la región, quienes nos indicaron lugares donde habían observado indicios (excretas, plumas, etc.) de posibles dormideros.
En todos los sitios de percha se contó el número de árboles utilizados como dormideros y con una cinta de lona de 50 m de longitud se midió la distancia entre ellos (< a 100 m), siguiendo el procedimiento del árbol de percha más cercano. Cuando la distancia entre los árboles fue de 100 m o más se consideró como un sitio diferente. La altura total de los árboles y a la primera rama se calculó a una distancia de 20 m de la base del árbol, en dirección horizontal (en contra de la pendiente), con una pistola Blume–Leiss®, y el diámetro normal se midió con una cinta diamétrica a 1.30 m de alto. La cobertura del dosel se estimó en la parte central de cada sitio donde se encontró la mayor cantidad de árboles utilizados como dormideros. Para ello, se modificó el procedimiento propuesto por Johansson (1985); se utilizó un tubo de PVC de 20 cm de longitud y 1 cm de diámetro, dividido en 8 secciones iguales (Márquez–Olivas, 2002). La densidad de árboles y arbustos (> a 2.5 cm de diámetro) se obtuvo de una parcela circular de 0.05 ha (12.6 m de radio), ubicada en el centro de cada sitio; lugar donde también se registraron la altitud, la cual se obtuvo con un altímetro Thommen®, la exposición de la ladera con una brújula Brunton® y los grados de la pendiente con un clinómetro Sunto® (Wakeling et al., 2001a).
En cada sitio se anotó el tipo de vegetación dominante (v.gr. bosque de pino, bosque de encino, bosque de pinoencino, etc.), se determinó el número de estratos vegetales, y con un GPS (Global Positional System) marca Garmin 12–XL se estimó la distancia a los abrevaderos y comederos más cercanos (claros del bosque, milpas y cebaderos). De todos los árboles que los guajolotes utilizaron como dormideros se colectaron muestras (hojas, conos y frutos), para su posterior identificación. El material botánico se identificó bajo la supervisión y apoyo del personal a cargo del Herbario de la Universidad Autónoma Chapingo.
Resultados
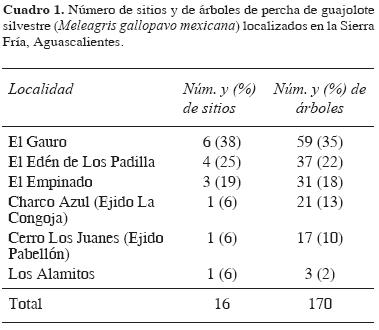
El trabajo de campo permitió la ubicación de 16 sitios de percha (7 en el año 2001 y 9 en el 2002), 10 de los cuales fueron localizados en 2 comunidades vecinas que concentraron el 56.5% de los árboles (n=170) utilizados como dormideros (Cuadro 1). El 87.5% de los sitios se encontraron en bosques de pino–encino (en rodales de pino) y el resto (12.5%) en bosques de encino, sobre la pendiente de pequeñas cañadas en una amplitud altitudinal de 2500 a 2650 metros. El 43% de los dormideros se hallaron en laderas de exposición noreste, 31% este, 12% noroeste y 7% en norte y oeste, en una pendiente promedio de 38.9 ± 12.7%. El 50% de los sitios se localizaron en la parte alta (2/3) de la cuesta, 37.5% en la parte media y 12.5% en la parte baja (1/3). Se contabilizaron de 1 a 23 árboles de percha utilizados por sitio, distribuidos a una distancia promedio de 24.4 ± 17.8 m (Cuadro 2). En el 31.3% de los sitios (n=16) se encontraron de 10 a 14 árboles dormidero; en el 25% de 5 a 9; en el 18.8% de 1 a 4 y de 15 a 19, y en el 6.3% de 20 a 24 árboles. El 62.5% de los sitios de percha se usaron frecuentemente durante todo el periodo de estudio y el resto (37.5%) de manera ocasional. Se localizaron 8 de los dormideros en una área aproximada de 300 ha y la mayor parte se usaron de manera constante durante todo el estudio. Todos los sitios de descanso o dormideros se localizaron en los rodales del bosque con mayor edad, fuste, altura y cobertura. En esos sitios la densidad promedio de árboles fue de 351 por hectárea y la cobertura del dosel del 57.5% (Cuadro 2); además se identificó algún tipo de alteración reciente (v.gr., tala de árboles o incendios forestales). Los sitios se encontraron a una distancia promedio de los abrevaderos de 246.3 ± 139.9 m y a 160 ± 73.5 m de los comederos (Cuadro 2), y relativamente cerca de viviendas (ranchos) habitadas de manera permanente (menos de 1.5 km).
El 81.2% de los árboles (n=170) que los guajolotes usaron como dormideros fueron pinos; 85 pertenecieron a Pinus teocote, 35 a P. durangensis, 11 a P. leiophylla y 7 a P. michoacana; el 17.6% fueron encinos, 12 Quercus laurina, 10 Q. grisea y 8 Q. eduardii, y el 1.2% madroños (Arbutus arizonica A. Gray). Nueve (5.3%) de los árboles de percha se encontraron secos; 6 correspondieron a árboles viejos (5 encinos y 1 pino) y 3 a pinos quemados. La altura de los árboles de percha promedió 16.6 m ± 4.4 m; a la primera rama 4.6 ± 2.4 m y el diámetro 43.2 ± 11.5 cm (Cuadro 3). El 48% de los árboles de percha tuvieron una altura de 10 a 15 m (Fig. 2) y el 63% un diámetro de 30 a 49 cm (Fig. 3 ).


Discusión
Las características físico–bióticas que se han documentado de los sitios que los guajolotes eligen para descansar, como la densidad de árboles por hectárea, la cobertura del dosel, la altura y el diámetro de los árboles de percha, las especies arbóreas utilizadas como perchaderos, y la ubicación y orientación de la pendiente, presentan algunas inconsistencias con las consignadas en esta investigación, en virtud de que estas aves se pueden encontrar en diferentes bosques, e incluso en áreas de matorral (Carrillo–Reyes, 2003), y se adaptan con facilidad a las condiciones ambientales de cada lugar, según la época del año y el grado de alteración del hábitat. Como ejemplo, York (1991) encontró el 97% de los dormideros en rodales de Pinus chihuahuana sobremaduros en la parte baja de las riberas; Valencia y López (2000) hallaron cerca del 70% de las perchas (n = 33) en bosques de encinopino (en rodales de pino), en altitudes mayores de 2650 m; Garza (2005) encontró la mayor parte de los dormideros en los encinares ya que prácticamente durante todo el año laderas con exposición sur y Hoffman et al. (1993) citan que el guajolote de Merriam descansa durante el invierno en bosques de Pinus ponderosa y en rodales de piñonero y Juniperus distribuidos en las partes inferiores de las laderas. Por su parte, Carrillo–Reyes (2003) encontró que los guajolotes de Río Grande (M. g. intermedia) no usaron no usaron los encinares ya que prácticamente durante todo el año esa subespecie percha en áreas ribereñas, en bosques de alamillos o sicomoros (Platanus occidentalis).
En el primer informe sobre el monitoreo de los guajolotes reintroducidos en Sierra Fría se cita que las áreas ribereñas con vegetación de pino–encino, en asociación con herbáceas y arbustivas, son las más utilizadas por esta especie quien prefiere los pinos de mayor tamaño para perchar (Clemente, 1993).
Se ha notado que los sitios de percha se sitúan de manera habitual en lugares estratégicos, de preferencia cerca de donde haya agua y alimento y donde los guajolotes puedan desplazarse (subir y bajar de los árboles) con poco esfuerzo y encuentren cierta protección contra depredadores y el clima (Mackey, 1984; Schemnitz et al., 1985; Lutz y Crawford, 1987; Valencia y López, 2000). Mellink (1989) refiere que los guajolotes requieren para pernoctar bosques maduros incoetáneos, en lugares protegidos del viento, a menos de 800 m del agua. La altura de los árboles, la orientación de la pendiente y la parte en que se ubican los dormideros (alta, media o baja de la pendiente) es fundamental para la seguridad de los guajolotes. Garza (2005) halló que, en el invierno, cuando los dormideros se ubicaron en laderas con exposición distinta a la exposición sur éstos siempre estuvieron protegidos por riscos y laderas de mayor altura.
En este estudio la mayoría de los dormideros se localizaron en la parte noreste de las cañadas, lo que coincide con las observaciones de otros autores quienes citan que los guajolotes utilizan esos lugares de las riberas para protegerse del viento, sobre todo cuando los dormideros se ubican en lo alto de la cuesta, así como para recibir más temprano el amanecer y porque los árboles que prefieren para descansar se encuentran regularmente en esas áreas (Jonas, 1966; Boeker y Scott 1969; Rumble, 1992).
La orientación de los dormideros se ha relacionado más con el estado del tiempo que con la distribución de la vegetación y las condiciones microclimáticas en el hábitat, lo cual podría explicar también la ubicación de las áreas que utiliza el guajolote silvestre para dormir. Como ejemplo, en el oeste de los Estados Unidos de América, el árbol que más comúnmente selecciona el guajolote de Merriam para descansar es Pinus ponderosa que crece o se asocia a sitios deficientes en lluvia, típico de la parte este de las laderas (Fowells, 1965). Se ha notado también que durante el invierno esa subespecie prefiere el sur de las laderas, porque son áreas más cálidas y la nieve se derrite con mayor rapidez (Rumble y Anderson, 1996).
El porcentaje de la pendiente de los sitios de percha fue menor que el que citan Valencia y López (2000), pero coincide con el de Lafón (1997), 38.9 vs. 57.9 y 36.5%, respectivamente. Usualmente el guajolote silvestre selecciona sitios de pendientes inclinadas (> a 35%), pero también utiliza pendientes moderadas (20 a 30%) y de manera ocasional menores del 15% (Jonas, 1966; Scott y Boeker, 1975; Lutz y Crawford, 1987; Kilpatrick et al., 1988), especialmente durante el verano. Las laderas más inclinadas favorecen a los guajolotes para trepar a los árboles cuando van a dormir y les permite movilizarse con poco esfuerzo en las mañanas, cuando dejan las perchas para acudir a los sitios donde se alimentan o beben agua, mientras que los dormideros ubicados en pendientes suaves o planas dificultan esa labor y sólo se usan cuando facilitan el acceso a las perchas.
Los abrevaderos son un componente esencial del hábitat del guajolote silvestre durante el invierno o sequía. Boeker y Scott (1969) citan que la disponibilidad de agua puede influir en la selección de los sitios de descanso, pues regularmente se ve a los guajolotes beber agua poco tiempo después de abandonar los dormideros. Se ha encontrado que los sitios de percha se sitúan más cerca del agua en invierno que en el verano, ya que durante esta estación del año los alimentos no contienen el agua suficiente para satisfacer sus necesidades metabólicas. Como ejemplo, en el verano Goerndt (1983) encontró los dormideros a una distancia promedio del agua de 1670 m y en el invierno de 322 m.
En este estudio los dormideros de los guajolotes se encontraron en áreas con pocos arbustos y una alta densidad de árboles adultos de altura variable. La presencia de tocones y árboles caídos, algunos utilizados como acceso a los árboles de percha, fueron comunes en estos sitios. El estrato herbáceo en los sitios localizados en propiedades privadas, como El Edén de Los Padilla, El Gauro y Los Alamitos, se observó en buena condición, mientras que en áreas de acceso común o ejidales fue prácticamente nulo.
El número de árboles utilizados como dormideros por sitio fue muy semejante al que encontraron Mackey (1984) y Valencia y López (2000), 1–24 y 1–21 respectivamente, pero mayor que el consignado por Kilpatrick et al. (1988) y Rumble (1992), 1–7 y 1–9, y menor al que registran Hoffman (1968) y Boeker y Scott (1969), 4–44 y 1–37 de manera respectiva. Los sitios donde se encontraron más árboles de percha se usaron con mayor frecuencia; pero, según la acumulación de heces, no todos los árboles se usaron con la misma intensidad, ya que unos cuantos albergaron más guajolotes que otros y se usaron de manera más constante. Suponemos, aunque es poco probable, que hay cierta jerarquía en la interacción de estas aves para ocupar los dormideros, de tal suerte que los guajolotes dominantes podrían ocupar las mejores perchas y los demás dispersarse en árboles de alrededor; por consiguiente, entre más grande la parvada mayor el número de árboles utilizados. En el cuadro 2 se presenta la distancia promedio de los árboles de percha encontrados por sitio, así como la de los que se localizan más cerca y la de los más alejados, lo que muestra la dispersión de los árboles utilizados en cada lugar.
Se ha observado que en invierno, más que en verano, los guajolotes regresan a los mismos sitios de descanso, ya que durante esta parte del año se dispersan y aumenta su área de actividad. Durante el invierno, los sitios de descanso son más grandes (se concentra un mayor número de guajolotes en un mismo lugar) y se utilizan con mayor frecuencia, mientras que en el verano son pequeños, se eligen al azar y rara vez se usan de forma continua (Boeker y Scott, 1969; Lutz y Crawford, 1987; Lafón, 1997). Como ejemplo, Porter (1980) encontró que en el verano los dormideros se usaron con una frecuencia de 20%, York (1991) 49% y Zornes (1993) 33%.
El uso constante de los mismos sitios y árboles de descanso puede ser letal para los guajolotes, pues atrae a los depredadores (entre los que se incluye a los humanos), que esperan pacientemente el momento de atraparlos, tal como se pudo corroborar durante esta investigación, ya que en un porcentaje elevado de los sitios más frecuentados se encontraron evidencias de depredación.
La mayoría de los sitios que se usaron con más frecuencia se ubicaron cerca de comederos que los administradores de los ranchos abastecen con granos de maíz y de pequeñas parcelas donde siembran maíz, avena y otras gramíneas, de manera exclusiva para la fauna silvestre. Esta tendencia también fue observada por Rumble (1992), quien cita que parvadas grandes de guajolotes duermen cerca de ranchos y unidades habitacionales, donde estas aves obtienen granos domésticos o se les facilita encontrar alimento. La concentración y el uso cotidiano de los dormideros en esas áreas demuestra que los guajolotes se acostumbran, más rápidamente de lo que nos imaginamos (Holbrook et al., 1987), a las intervenciones del hábitat, dado que en todos esos sitios es común encontrar algún tipo de alteración reciente, como apacentamiento de animales domésticos, actividad humana intensa, tala de árboles o incendios forestales, disturbios de los que se observaron señales claras en este trabajo.
Como se ha observado en numerosos estudios (Goerndt, 1983; Mackey, 1984; Schemnitz et al., 1985; York, 1991; Hoffman et al., 1993; Miller et al., 1999; Valencia y López, 2000), en Sierra Fría el guajolote silvestre eligió para descansar árboles del género Pinus, tal vez porque son árboles de mayor porte o por la estructuración y el tamaño de las ramas, lo que les permite posarse de manera cómoda y moverse con cierta facilidad (Haucke, 1975). Sin embargo, cuando hay pocos pinos, los guajolotes utilizan para dormir otros árboles de características semejantes, como se muestra en nuestros resultados, donde el 17.6% de los árboles dormideros fueron encinos (Quercus spp.). Asimismo, Scott y Muller–Using (1992) citan 71% de los dormideros en Platanus occidentalis y 29% en Pinus teocote y Kilpatrick et al. (1988) 43.7% en Pinus strobus y 24.2% en Quercus spp.; por el contrario, Schemnitz et al. (1985) encontraron todos los dormideros en pinos; información que coincide con lo registrado por Valencia y López (2000), quienes indican que a pesar de haber encontrado encinos del tamaño de los pinos, cerca de los sitios de descanso, los guajolotes no los usaron. Estos últimos autores encontraron el 26% (n=180) de los árboles dormideros secos, cifra muy superior a la presentada en este estudio que fue de 5.3%.
La abundancia y distribución de los pinares en Sierra Fría es muy reducida, ya que los puros encinares cubren cerca del 72% de los bosques templados de la región (Márquez–Olivas et al., 2005), y especies como Pinus michoacana y P. durangensis, que durante este estudio fueron utilizadas como dormideros, se encuentran representadas por árboles solitarios o poblaciones muy bajas y generalmente mezcladas con otras especies arbóreas. Aunado a lo anterior, del año 1995 al 2000, en la Sierra Fría de Aguascalientes se talaron entre 1500 y 2000 pinos plagados por el gusano descortezador (Dendroctonus mexicanus); en 2001, los árboles talados fueron 10 343 (Nuestro campo, 2002), lo que significa una disminución considerable de los árboles que los guajolotes prefieren para descansar y afecta la calidad de su hábitat.
Las características que posiblemente influyen más en la selección de los árboles que los guajolotes escogen como dormideros son el tamaño, la estructuración del dosel y la altura a la primera rama, aunque también se ha relacionado con otros aspectos de la vegetación que pueden intervenir en la regulación térmica (ganancia o pérdida de calor) de las aves (Mackey, 1984; Rumble, 1992). La altura promedio de los árboles de percha fue menor a la que citan Goerndt (1983), Lafón (1997) y Wakeling et al. (2001a), quienes encontraron los dormideros en árboles de más de 23 m de alto; mayor que la registrada por Valencia y López (2000) de 13.4 m, y semejante a la consignada por Kilpatrick et al. (1988), 18.9 ± 3.9 m, lo que demuestra el grado de crecimiento del estrato arbóreo según la especie, edad y condiciones ecológicas de cada sitio o región.
En el área de estudio es característico que los pinares maduros o sobremaduros tengan las primeras ramas colgantes y próximas al suelo. La primera rama es el acceso principal de los guajolotes a los árboles de percha, de tal modo que entre más distante se encuentre del suelo más trabajo les costará subir; sobre todo en pendientes moderadas o planas. La altura a la primera rama también es importante para la seguridad de los guajolotes; árboles con ramas cercanas al suelo, facilitan la labor de los depredadores y propician que estas aves sean más vulnerables. En este estudio, la altura promedio a la primera rama de los árboles de percha es semejante a la que citan Wakeling et al. (2001a), pero difiere de la mencionada por Valencia y López (2000): 4.5 vs. 5.9 y 8.7 m, respectivamente.
Wakeling et al. (2001a) encontraron que el área basal (m2 / ha) y el diámetro de los árboles fueron las variables que más influyeron en la selección de los sitios de descanso; Mackey (1984) cita que los guajolotes abandonan los dormideros cuando el área basal se reduce a 16.8 m2 / ha y Rumble (1992) menciona que el diámetro no es un criterio de selección apropiado. Sin embargo, los guajolotes acostumbran descansar en árboles altos y maduros (Hoffman, 1991) que regularmente tienen diámetros gruesos. En este estudio, el diámetro de los árboles presentó variación entre sitios y dentro de los sitios, producto de la heterogeneidad de la vegetación. El valor promedio (43.2 cm) del diámetro de los árboles de percha muestra muy poca diferencia con el de Goerndt (1983), Mackey (1984), Kilpatrick et al. (1988) y Valencia y López (2000), pero se diferencia de los presentados por Lafón (1997) y Wakeling et al. (2001a); 43.2 vs. 20 y 71 cm de manera respectiva, aunque el 8% de los árboles de percha (n=170) tuvieron más de 60 cm de diámetro y 10% entre 20 y 29 cm (Fig. 3).
La información consignada en este estudio es limitada, pero puede servir de base para identificar áreas con características de vegetación que son preferidas por el guajolote silvestre para dormir y excluirlas de la explotación forestal. Otras recomendaciones sobre el manejo del bosque para conservar y proveer de estos sitios a dichas especies se pueden consultar en Hoffman et al. (1993).
Agradecimientos
Esta investigación se llevó a cabo en parte con el apoyo del Instituto de Recursos Naturales del Colegio de Postgraduados. El primer autor agradece al CONACYT la beca crédito que le otorgó para hacer el doctorado. Asimismo, al maestro Diódoro Granados por permitir el uso del Herbario de la División de Ciencias Forestales de la Universidad Autónoma Chapingo y otorgar un espacio para guardar el material botánico recolectado y a Francisco Ramos Marchena por la asistencia en la identificación de las especies. Se agradece al señor Alberto Guerrero Pérez, propietario del Rancho El Gauro, por su hospitalidad durante la fase de campo y a Don Alfredo y familia, Cuco, Lupe y Rafael, por su ayuda e información. Los autores agradecen las observaciones de los revisores anónimos que mejoraron el contenido del manuscrito.
Literatura citada
Austin, D.E. y L.W. De Graff. 1975. Winter survival of wild turkey in the southern Adirondacks. Proceedings of the National Wild Turkey Symposium 3:55–60. [ Links ]
Bailey, R.W. y K.T. Rinell. 1967. Management of the eastern turkey in the northern hardwoods. In The wild turkey and its management, O.H. Hewitt (ed.). The Wildlife Society, Washington, D.C., p. 261–302. [ Links ]
Boeker, E.L. y V.E. Scott. 1969. Roost tree characteristics for Merriam's turkey. Journal of Wildlife Management 33:121–124. [ Links ]
Carrillo–Reyes, A. 2003. Uso de hábitat del guajolote silvestre (Meleagris gallopavo intermedia) en Lampazos de Naranjo, Nuevo León. Tesis de Maestría. Facultad de Ciencias Forestales. Universidad Autónoma de Nuevo León. Linares. 85 p. [ Links ]
Chamberlain, M.J., B.D. Leopold y L.W. Burger. 2000. Characteristics of roost sites of adult wild turkey female. Journal of Wildlife Management 64:1025–1032. [ Links ]
Clemente, S. F. 1993. Primer informe de avances sobre el monitoreo de una población de guajolote silvestre liberada en la serranía de Aguascalientes. Consejo Nacional de la Fauna, Delegación de Aguascalientes. 5 p. [ Links ]
Diario Oficial de la Federación. 2002. Norma Oficial Mexicana (NOM–059–ECOL–2001). Protección ambiental–Especies nativas de México de flora y fauna silvestres–Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio–Lista de especies en riesgo. Secretaría de Medio Ambiente y Recursos Naturales. Segunda sección. 6 de marzo de 2002. Poder Ejecutivo Federal, México, D. F. p. 21. [ Links ]
Dickson, J.G. 1992. The wild turkey: biology and management. National Wild turkey Federation, USDA Forest Service. Stackpole Books, Harrisburg, Pennsylvania. 463 p. [ Links ]
Eaton, S.W. 1992. Wild turkey. In The birds of North American. A. Poole, P. Stettenheim y F. Gill (eds.). Academy of Natural Sciences and American Ornithologists´ Union, Washington, D.C., p. 121–130. [ Links ]
Fowells, H.A. 1965. Silvics of forest trees of the United States. USDA Agricultural Handbook. 271. Washington, D.C. 762 p. [ Links ]
Garza, A y J. Servín. 1993. Estimación de la población y utilización del hábitat del cócono silvestre (Meleagris gallopavo, Aves: Phasianidae) en Durango, México. Ecología Austral 3:15–23. [ Links ]
Garza, A. 2005. Biología y ecología del cócono o guajolote silvestre en Durango (Aves: Meleagris gallopavo). Tesis Maestría, Facultad de Ciencias, Universidad Nacional Autónoma de México. México, D.F. 87 p. [ Links ]
Glover, F.A. 1948. Winter activities of wild turkey in west Virginia. Journal of Wildlife Management 12:416–427. [ Links ]
Goerndt, D.L. 1983. Merriam's turkey habitat in relation to grazing and timber management of a mixed conifer forest in southcentral New Mexico. M.S. thesis. New Mexico State University, Las Cruces. 96 p. [ Links ]
Good, P.L. 1982. Winter habitat of the wild turkey in southeastern New Hampshire. M.S. thesis, University of New Hampshire, Durham. 61 p. [ Links ]
Haucke, H.H. 1975. Winter roost characteristics of the Rio Grande turkey in south Texas. Proceedings of the National Wild Turkey Symposium 3:164–169. [ Links ]
Hesselbach, M.H. 1996. Diversidad biológica de la Sierra Fría. Cuaderno de Trabajo 44: Agricultura y Recursos Naturales. Gobierno del Estado de Aguascalientes. 19 p. [ Links ]
Hoffman, R.W. 1991. Spring movements, roosting activities, and home–range characteristics of male Merriam's wild turkey. Southwestern Naturalist 36:332–337. [ Links ]
Hoffman, R.W., H.G. Shaw, M.A. Rumble, B.F. Wakeling, C.M. Mollohan, S.D. Schemnitz, R.E. Wilson y D.A Hengel. 1993. Management guidelines for Merriam's wild turkeys. Colorado Division of Wildlife in cooperation with USDA Forest Service, Rocky Mountain Forest Range Experiment Station, Division Game Report No. 18. Fort Collins, Colorado. 24 p. [ Links ]
Hoffman, D.M. 1968. Roosting sites and habitats of Merriam's turkeys in Colorado. Journal of Wildlife Management 32:859–866. [ Links ]
Holbrook, H.T, M.F. Vaughan y P.T. Bromley. 1987. Wild turkey habitat preferences and recruitment in intensively managed piedmont forest. Journal of Wildlife Management 51:182–187. [ Links ]
INEGI (Instituto Nacional de Estadística Geografía e Informática), 1981. Síntesis geográfica del estado de Aguascalientes. México, D.F. 98 p. [ Links ]
INEGI (Instituto Nacional de Estadística Geografía e Informática), 1993. Estudios Hidrológicos del estado de Aguascalientes. México, D.F. 164 p. [ Links ]
Johansson, T. 1985. Estimating canopy density by the vertical tube method. Forest Ecology and Management 11:139–144. [ Links ]
Jonas, R. 1966. Merriam's turkeys in southeastern Montana. Montana Game and Fish Department. Technical Bulletin No. 3. 36 p. [ Links ]
Kennamer, M.C. (s/f). Gould's wild turkey (Meleagris gallopavo mexicana). National Wild Turkey Federation, Wildlife Bulletin. http://www.nwtf.org/conservation/bulletins/bulletin_5.pdf [ Links ]
Kilpatrick, H.J., T.P. Husband y C.A. Pringle. 1988. Winter roost site characteristics of eastern wild turkey. Journal of Wildlife Management 52:461–463. [ Links ]
Lafón, A. 1997. Distribution, habitat use and ecology of Gould's turkey in Chihuahua, Mexico. Ph. D. Thesis, New Mexico State University, Las Cruces. 155 p. [ Links ]
Leopold, A. S. 1977. Fauna silvestre de México. IMERNAR,. México, D.F. 641 p. [ Links ]
Lewis, J.C. 1973. The world of the wild turkey. Lippincott, New York. 158 p. [ Links ]
Lutz, R.S. y J.A. Crawford. 1987. Seasonal use of roost sites by Merriam's wild turkey hens and hen–poult flocks in Oregon. Northwest Science 61:174–178. [ Links ]
Mackey, D. L. 1984. Roosting habitat of Merriam's turkeys in south–central Washington. Journal of Wildlife Management 48:1377–1382. [ Links ]
Márquez–Olivas, M. 2002. Caracterización de hábitat del tecolote moteado mexicano (Strix occidentalis lucida (X) Nelson, 1903) en Sierra Fría, Aguascalientes. Agrociencia 36:541–546. [ Links ]
Márquez–Olivas, M., E. García, C. González–Rebeles y L. Tarango. 2005. Composición de la dieta del guajolote silvestre (Meleagris gallopavo mexicana Gould, 1856) reintroducido en la 'Sierra Fría' de Aguascalientes, México. Veterinaria México 63(4):395–409. [ Links ]
Mellink, E.1989. Estudio faunístico de la Unidad Industrial de Explotación Forestal de Atenquique. In Plan de Manejo Integral Forestal de la Región de Atenquique (PMIFRA), R.L.J. Flores, C.S. Juárez, B.J. Gallegos y B.M.A. Mendoza (Redactores). Memoria General, PMIFRA. Compañía Industrial de Atenquique, Jalisco, p. 148–164. [ Links ]
Miller, D.A., G.A. Hurst y B.D. Leopold. 1999. Habitat use of eastern wild turkeys in central Mississippi. Journal of Wildlife Management 63:210–222. [ Links ]
Nuestro Campo. 2002. Revista Informativa de la Comisión para el Desarrollo Agropecuario del Estado de Aguascalientes. Año 1, No. 5. [ Links ]
NWF (National Wildlife Federation). 1982. Assessment of the Weyerhaeuser Company's forestry operations in southwestern Arkansas and southeastern Oklahoma. National Wildlife Federation. Washington, D.C. 77 p. [ Links ]
Periódico oficial. 1994. Decreto 88 de la LV Legislatura. Órgano del Gobierno Constitucional del Estado de Aguascalientes. Decreto 88 de la LV Legislatura, 30 de enero de 1994.
Porter, W.F. 1978. The ecology and behavior of the wild turkey (Meleagris gallopavo) in southeastern Minnesota. Ph. D. Thesis, University of Minnesota, St. Paul. 122 p. [ Links ]
Porter, W.F. 1980. An evaluation of wild turkey brood habitat in southeastern Minnesota. Proceeding National Wild Turkey Symposium 4:203–212. [ Links ]
Rumble, M.A. 1992. Roosting habitat of Merriam's turkeys in the Black Hill, South Dakota. Journal of Wildlife Management 56:750–759. [ Links ]
Rumble, M.A. y S.H. Anderson. 1996. Microhabitats of Merriam's turkeys in the Black Hill, South Dakota. Ecological Applications 6:326–334. [ Links ]
Schemnitz, S.D., D.L. Goerndt y K,H. Jones. 1985. Habitat needs and management of Merriam's turkey in southcentral New Mexico. Proceeding National Wild Turkey Symposium 5:199–231. [ Links ]
SEDESOL (Secretaría de Desarrollo Social). 1993. Estudio para la declaratoria de la Sierra Fría como Área Natural Protegida. Informe interno. Subsecretaría de Ecología del Estado de Aguascalientes. 286 p. [ Links ]
SEDESOL (Secretaría de Desarrollo Social). 1996. Sierra Fría, Área Natural Protegida. Tríptico divulgativo. Subsecretaría de Ecología del Estado de Aguascalientes. [ Links ]
Scott, V.E. y E.L. Boeker. 1977. Responses of Merriam's turkey to pinyon–juniper control. Journal of Range Management 30:220–223. [ Links ]
Scott, L. y B. Muller–Using. 1992. Aspectos ecológicos de una población de guajolote silvestre (Meleagris gallopavo) al sureste de Nuevo León, México. Reporte científico 30. Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León, Linares. 38 p. [ Links ]
Valencia, O.G. y S.E. López. 2000. Uso–disponibilidad de hábitat y presencia de enfermedades en el guajolote de Gould en el suroeste del estado de Sonora. Instituto del Medio Ambiente y el Desarrollo Sustentable del Estado de Sonora (IMADES). Reporte final. 24 p. [ Links ]
Villalobos, S.V. 1998. El venado cola blanca en la Sierra Fría de Aguascalientes. Cuaderno de Trabajo 89: Agricultura y Recursos Naturales. Gobierno del Estado de Aguascalientes. 33 p. [ Links ]
Wakeling, B.F., S.R. Boe, M.M. Koloszar y T.D. Rogers. 2001. Gould's turkey survival and habitat selection modeling in southeastern Arizona. Proceedings of the National Wild Turkey Symposium 8:101–108. [ Links ]
Wakeling, B.F., S. Weimann, T. Jackson, B. Dowler, M. Eacret y J. Gillis. 2001a. Ponderosa pine tree selection by roosting Merriam's turkeys in north–central Arizona. Biennial Conference of Research on the Colorado Plateau 5:119–124. [ Links ]
Wunz, G.A. y A.H. Hayden. 1975. Winter mortality and supplemental feeding of turkeys in Pennsylvania. Proceedings of the National Wild Turkey Symposium 3:61–69. [ Links ]
York, D.L. 1991. Habitat use, diet, movements and home range of Gould's turkey in the Peloncillo Mountains. M.S. Thesis. New Mexico State University, Las Cruces. 104 p. [ Links ]
Zornes, M.L. 1993. Ecology and habitat evaluation of Gould's turkey in the Peloncillo Mountains. M.S. Thesis New Mexico State University, Las Cruces. 117p. [ Links ]
Zwank, P.J., T.H. Jr. Whitre y F.G. Kimmel. 1988. Female turkey habitat use in Mississippi River Batture. Journal of Wildlife Management 52:253–260. [ Links ]

