










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de biodiversidad
versión On-line ISSN 2007-8706versión impresa ISSN 1870-3453
Rev. Mex. Biodiv. vol.76 no.1 México jun. 2005
Taxonomía y sistemática
Análisis morfométrico de los peces del grupo labialis, género Profundulus (Cyprinodontiformes: Profundulidae), en Chiapas, México
Morphometric analysis of the fish included in the labialis group, genus Profundulus (Cyprinodontiformes: Profundulidae), in Chiapas, Mexico
Alfonso A. González-Díaz¹*, Edmundo Díaz-Pardo², Miriam Soria-Barreto³ y Rocío Rodiles-Hernández¹
¹El Colegio de la Frontera Sur. Carretera Panamericana y Periférico Sur s/n, A.P. 63, San Cristóbal de Las Casas, C.P. 29290. Chiapas. México. Tel. 01 967 6749000 ext. 1322.
*Correspondencia: agonzalez@sclc.ecosur.mx, aangelgd@yahoo.com.mx
²Universidad Autónoma de Querétaro. Facultad de Ciencias Naturales-Biología. Laboratorio de Conservación de Peces y Hábitats Acuáticos. Centro Universitario s/n, Cerro de las Campanas, Querétaro, 76010, México.
³Departamento de Ingeniería en Sistemas Ambientales. Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional. Carpio y Plan de Ayala s/n, Col. Plutarco Elías Calles, México, D.F., 11340, México.
Recibido: 05 de julio del 2004
Aceptado: 18 de mayo del 2005
Resumen: Se presentan los resultados de la comparación morfométrica de las especies del "complejo labialis", del género Profundulus, basada en 11 medidas tradicionales y 21 cruzadas. Mediante el análisis de componentes principales y agrupamiento se demostró que Profundulus labialis y P. candalarius muestran un patrón morfológico semejante, caracterizado por mayor altura del cuerpo, menor longitud del hocico, y menor longitud y ancho de la boca; mientras que P. hildebrandi presenta características diferentes. El análisis de varianza mostró 14 medidas morfométricas estadísticamente significativas que permiten separar, por lo menos, a una de las tres especies y además corroboran su validez taxonómica. Las relaciones de similitud establecidas en este análisis concuerdan con las relaciones filogenéticas propuestas por otros autores. Las diferencias encontradas en medidas relacionadas con estructuras tróficas y de la altura del cuerpo podrían estar relacionadas con aspectos ecológicos o asociaciones con distintos hábitats.
Palabras clave: morfometría comparada, peces mexicanos.
Abstract. Results of morphometrical comparison of the species of the "labialis complex", of the genus Profundulus, based on 11 traditional and 21 crossed measures, are herein presented. Through the principal components and cluster analyses we have found that Profundulus labialis and P. candalarius show a similar morphological pattern, characterized by the greater altitude, the shorter snout, and the shorter length and width of the mouth; while P. hildebrandi has the opposite characteristics. The analysis of variance showed 14 statistically different measures that allow to recognize at least one of the three species, and they also corroborate their taxonomical validity. The results of the present analyses of similarities are congruent with the phylogenetic relationships proposed by other authors. Some measurements associated to trophic structures seem to vary according to ecological conditions.
Key words: comparative morphometry, mexican fishes.
Introducción
El género Profundulus es un taxón representativo de la ictiofauna centroamericana. Su distribución abarca las dos vertientes continentales: sobre el Pacífico, desde Acapulco, Guerrero, México al oeste de Honduras, y sobre el Atlántico, desde el istmo de Tehuantepec hasta el río Motagua en Guatemala. Actualmente se reconocen cinco especies, pertenecientes a los grupos punctatus y labialis. El primero compuesto por Profundulus guatemalensis y P. punctatus; y el segundo por Profundulus candalarius, con distribución en la cuenca de Comitán, Chiapas, México y Río Candelaria, Guatemala, P. hildebrandi, endémica de San Cristóbal de Las Casas, Chiapas, México y P. labialis en la cuenca Grijalva-Usumacinta, Chiapas, México y Guatemala (Miller, 1955; Espinosa et al., 1993).
El conocimiento sistemático del género Profundulus es insuficiente y su clasificación ha variado según el criterio de distintos autores, ya que Profundulus y Tlaloc se han considerado alternativamente como distintos géneros o como subgéneros (Doadrio et al., 1999a). Asimismo, la separación de especies resulta difícil debido a su gran variación intra e interpoblacional.
Recientemente se analizaron las relaciones filogenéticas entre los géneros de la familia Profundulidae, mediante el empleo de marcadores moleculares y electroforesis de aloenzimas. Se estableció que los grupos punctatus y labialis son monofiléticos, siendo P. hildebrandi la especie basal de este último, entre tanto P. candalarius y P. labialis aparecieron como grupos hermanos. En el grupo punctatus se propusieron dos especies nuevas con base en caracteres genéticos, lo cual comprueba que la variación morfológica observada en estos peces tiene un sustento genético (Doadrio et al., 1999a, com. pers.). La variabilidad observada en el grupo labialis también sugiere la existencia de nuevas especies o formas en el estado de Chiapas, México (Miller, 1955).
Estos trabajos destacan la necesidad de buscar nuevas características morfológicas que avalen las diferencias establecidas sobre la base de datos moleculares, asimismo sugieren que es preciso realizar estudios más detallados para reconstruir las relaciones filogenéticas dentro del género Profundulus (Doadrio et al., 1999a, com. pers.).
La bondad del uso de las medidas tradicionales en el reconocimiento de especies es ampliamente conocida; sin embargo, el empleo de medidas cruzadas, a pesar de dar buenos resultados, no se ha presentado con frecuencia en estudios taxonómicos de peces mexicanos. La falta de caracteres diagnósticos en el nivel específico dificulta la tarea taxonómica, el presente estudio tiene por objeto determinar la existencia de atributos morfométricos discriminantes que contribuyan al reconocimiento de las especies del grupo labialis.
Materiales y métodos
El material biológico analizado procede de ejemplares depositados en las Colecciones de Peces del Colegio de la Frontera Sur, Unidad San Cristóbal (ECO-SC-P) y del Museo de Zoología de la Universidad de Ciencias y Artes de Chiapas (MZ), con los siguientes números de registro; Profundulus candalarius: laguna de Teopisca, Chiapas, ECO-SC-P3997, ECO-SC-P3998, MZ066 y MZ014; P. hildebrandi: cuenca de San Cristóbal, San Cristóbal de Las Casas, Chiapas, ECO-SC-P2821, P2827, P2829, P3824 al P3832, P3834, P3835, P3838, P3924, P3925, P3926, P3829 y P4007; P. labialis: ríos Blanco y Trapiche, Chiapas, ECO-SC-P3927 y P3991.
La identidad taxonómica de estos peces fue corroborada empleando las claves de Miller (1955) y Álvarez (1970) y se seleccionaron 45 ejemplares adultos de cada especie con tallas de 40 a 100 mm de longitud patrón (LP). De cada ejemplar se obtuvieron 11 de las medidas "tradicionales" propuestas por Hubbs y Lagler (1958), empleando un vernier electrónico con precisión de 0.01 mm: 1, longitud cefálica; 2, longitud de la boca; 3, diámetro ocular; 4, distancia interorbital, 5, distancia predorsal; 6, altura máxima del cuerpo; 7, altura del pedúnculo caudal; 8, ancho de la cabeza; 9, distancia interpectoral; 10, ancho de la boca, y 11, longitud del hocico. Las medidas se expresaron en porcentaje con respecto a la longitud patrón, con el propósito de reducir el factor alométrico causado por la diferencia en el tamaño de los ejemplares (Crisci y López-Armengol, 1983). Estos datos fueron empleados en el análisis de agrupamiento aplicando el índice de disimilitud de distancia euclidiana, cuyos resultados se observan mediante la construcción de un dendrograma a través del método de ligamiento promedio no ponderado (UPGMA), con el fin de evidenciar relaciones de similitud morfológica entre las especies (Sneath y Sokal, 1973).
Posteriormente, las medidas lineales fueron transformadas a logaritmos base diez. Estos datos se usaron en el análisis de componentes principales (ACP) con la matriz de varianza-covarianza, a través del programa ANACOM Versión 3.0 (De la Cruz, 1994), para establecer el patrón de variación morfológica entre las especies e identificar las medidas que determinan la formación de grupos dentro del espacio multivariado (Crisci y López-Armengol, 1983). Las medidas más importantes se identificaron por sobrepasar el valor absoluto 0.3 en los eigenvalores (Soria, 1999; González, 2001; Catena et al., 2003). A cada medida, expresada en proporciones, se le aplicó un análisis de varianza (ANOVA, programa SPSS Versión 10.0.1) con un nivel de significancia de 0.05 y una prueba de DHS (Diferencia Honestamente Significativa) de Tukey, (SPSS Versión 10.0.1) con la finalidad de confirmar la presencia de diferencias interespecíficas estadísticamente significativas.
Para cada ejemplar se tomaron 21 medidas, siguiendo la metodología de cuadriláteros contiguos propuesta por Strauss y Bookstein (1982), donde se construye la forma del cuerpo mediante líneas cruzadas (Fig. 1): 1, margen anterior del premaxilar a la articulación del cleitro; 2, articulación del cleitro al origen de la aleta pélvica; 3, margen anterior del premaxilar al origen de la aleta pélvica; 4, cleitro al occipucio; 5, margen anterior del premaxilar al occipucio; 6, occipucio al origen de la aleta pélvica; 7, origen de la aleta pélvica al origen de la aleta anal; 8, occipucio al origen de la aleta anal; 9, origen de la aleta anal al origen de la aleta dorsal; 10, occipucio al origen de la aleta dorsal; 11, origen de la aleta dorsal al origen de la aleta anal; 12, base de la aleta anal; 13, origen de la aleta dorsal a la inserción posterior de la aleta anal; 14, origen de la aleta anal a la inserción posterior de la aleta dorsal; 15, base de la aleta dorsal; 16, inserción posterior de la aleta dorsal a la inserción posterior de la aleta anal; 17, inserción posterior de la aleta anal al radio inferior de la aleta caudal; 18, inserción posterior de la aleta dorsal al radio ventral de la aleta caudal; 19, inserción posterior de la aleta anal al radio dorsal de la aleta caudal; 20, inserción posterior de la aleta dorsal al radio dorsal de la aleta caudal; 21, radio dorsal de la aleta caudal al radio ventral de la aleta caudal. Estas medidas fueron expresadas en porcentaje de la longitud patrón y posteriormente sometidas a un análisis de varianza para conocer aquellas estadísticamente significativas, las cuales se señalan en los dibujos de las especies.

Figura 1. Medidas utilizadas en el método de cuadriláteros contiguos.
Resultados
A partir del análisis de similitud se formaron dos grupos principales, el más pequeño integrado básicamente por los ejemplares de P. hildebrandi, quienes mostraron menor parecido morfológico con los de las otras dos especies (5.6 unidades de disimilitud). Por otra parte, el segundo grupo incluyó los organismos de P. labialis y P. candalarius; esta última especie mostró la mayor variación morfológica, incluso algunos de sus representantes se unieron al primer grupo (Fig. 2).

Figura 2. Dendrograma producto del índice de disimilitud de distancia euclidiana de los individuos de Profundulus. P. candalarius, P. labialis, P. hildebrandi.
Los primeros dos componentes del ACP evidenciaron mejor la separación entre las especies, explicando el 77.02% de la varianza. En el primero la longitud y ancho de la boca (2 y 10), longitud del hocico (11) y altura máxima del cuerpo (6) tuvieron las medidas con los eigenvalores más altos, explicando el 65.91% de la varianza (Cuadro 1). En la región positiva de este eje se ubicaron los organismos de P. hildebrandi, que se caracterizan por por la longitud del hocico y la longitud y ancho de la boca mayores, así como menor altura del cuerpo. En la región negativa se encontró a P. labialis, que se distingue por valores mayores de altura del cuerpo, pero medidas más pequeñas en la longitud del hocico y longitud y ancho de la boca. Los ejemplares de P. candalarius están en el medio y sus individuos se superponen con los de las otras dos especies, por eso resulta difícil separarlos (Fig. 3).
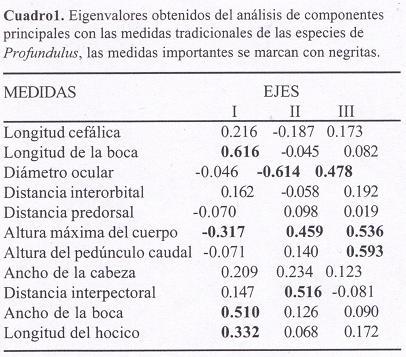

Figura 3. Análisis de componentes principales de las medidas tradicionales en logaritmo de las especies de Profundulus.
El segundo componente explicó el 11.11% de la varianza y las medidas más importantes fueron el diámetro ocular (3), la distancia interpectoral (9) y la altura máxima del cuerpo (6) (Cuadro 1). Los organismos de P. labialis se hallan hacia la parte positiva, diferenciados por tener mayor la distancia interpectoral y altura máxima, así como menor diámetro ocular. Por su parte, los individuos de P. candalarius tienden a ubicarse hacia la región negativa, caracterizados por que la distancia interpectoral y la altura máxima son más pequeñas y el diámetro ocular es mayor. Los representantes de P. hildebrandi se distribuyeron a todo lo largo del eje, registrando valores intermedios entre las medidas mencionadas anteriormente (Fig. 3). El tercer componente no tuvo un patrón claro en la delimitación de los grupos al explicar sólo el 8.26 % de la varianza, por lo tanto no se graficó.
El análisis de varianza señala que entre las medidas empleadas en el ACP, las que se consideran estadísticamente discriminantes para las tres especies (P<0.05) son: la longitud de la boca (2) (F=184.103), la altura máxima del cuerpo (6) (F = 112.271), la distancia interpectoral (9) (F = 20.988), el ancho de la boca (10) (F = 157.726) y la longitud del hocico (11) (F = 99.577); por otra parte, el mayor diámetro ocular (3) discrimina a P. candalarius (F = 25.87) y la altura mínima del pedúnculo caudal (7) a P. hildebrandi (F = 19.036) (Cuadro 2).
La reconstrucción morfológica de las especies, hace evidente que los individuos de P. labialis y P. candalarius son más altos que los de P. hildebrandi. Esta condición se refleja en la forma más alargada de P. hildebrandi, cuya altura del pedúnculo caudal se aproxima a la del cuerpo. El perfil dorsal de la cabeza también muestra diferencias; en P. hildebrandi está dirigido hacia arriba, casi paralelo al margen dorsal del cuerpo, mientras que en P. labialis y P. candalarius se orienta ligeramente hacia la región ventral (Fig. 4).

Figura 4. Reconstrucción morfológica de las especies de Profundulus a partir el método de cuadriláteros contiguos.
Entre las medidas cruzadas (método de los cuadriláteros contiguos), las estadísticamente significativas según el análisis de ANOVA fueron las siguientes (Cuadro 3): la distancia del origen de la aleta dorsal al origen de la aleta anal (11) (F = 117.812) y del margen anterior del premaxilar a la articulación del cleitro (1) (F = 125.824), son estadísticamente significativas para las tres especies; asimismo, P. hildebrandi se discrimina por la distancia del origen de la aleta dorsal a la inserción posterior de la aleta anal (13) (F = 77.976), la distancia de la inserción posterior de la aleta dorsal al radio dorsal de la aleta caudal (19) (F = 36.281), la longitud de la base de la aleta anal (12) (F = 37.810) y del radio dorsal de la aleta caudal al radio ventral (21) (F = 8.981); mientras que P. labialis se diferencia por la longitud de la base de la aleta dorsal (15)
(F = 24.853) (Cuadro 3, Fig. 4).
Discusión
El análisis de agrupamiento mostró que la semejanza morfológica es mayor entre Profundulus candalarius y P. labialis; en cambio, P. hildebrandi fue el taxón menos parecido. Estos resultados fueron corroborados con el ACP, en donde se evidenció la formación de tres grupos. Dentro del patrón morfológico observado en el espacio multivariado es evidente la separación entre los individuos de P. labialis y P. hildebrandi. Esta situación no es tan clara en P. candalarius, debido a que la dispersión de las medidas dentro de la especie hacen que su variación se superponga con la de las otras dos especies.
Algunos de los caracteres empleados en estudios filogenéticos para establecer las relaciones entre las especies de la familia Profundulidae, coinciden con los de mayores valores discriminatorios encontrados en el presente estudio, principalmente aquellos relacionados con la boca. De esta manera, probablemente el parecido morfológico entre P. labialis y P. candalarius sea producto del aislamiento geográfico reciente y de las estrechas relaciones de parentesco, factores que también estarían explicando algunas de las diferencias con P. hildebrandi.
Al respecto, Miller (1955) indicó que las diferencias morfológicas que se observan en la forma de la forma de la boca y del maxilar de P. hildebrandi han sido reforzadas, además, por procesos ecológicos relacionados con la alimentación y representan convergencias evolutivas con las especies del grupo punctatus. Esto podría corroborar y explicar las evidentes diferencias morfométricas encontradas en este estudio entre la última especie con P. labialis y P. candalarius, las cuales se relacionan con el tamaño de la boca y del hocico (Cuadro 2).
Los patrones morfológicos que aquí se presentan y los de Miller (1955) encuentran sustento en las relaciones filogenéticas obtenidas a través de datos moleculares, que señalan a P. labialis y P. candalarius como especies hermanas, mientras que P. hildebrandi representa la especie basal (Doadrio et al., com. pers.), que, además, ha sido fuertemente afectada por factores ambientales. También se corrobora la supuesta relación entre los cambios genéticos de estas especies y sus características morfológicas, tal como se menciona en los taxa del grupo punctatus del género Profundulus (Doadrio et al. 1999a, com. pers.). Los resultados aquí presentados son preliminares y es preciso realizar estudios de otras poblaciones, principalmente de las especies con amplias distribuciones geográficas como P. candalarius y P. labialis, que se registran desde Chiapas hasta Guatemala (Miller, 1950, 1955) y pueden presentar mayor variación que modifique los resultados aquí mostrados.
El uso de medidas cruzadas permitió encontrar nuevas medidas discriminantes, que junto con las tradicionales corroboran la identidad taxonómica de las tres especies. Entre las medidas tradicionales destacan las de la región cefálica y en particular de la boca; mientras que en las cruzadas son evidentes las diferencias en aquellas relacionadas con la altura del cuerpo y orientación de la boca. Estas medidas pueden ser utilizadas en la determinación taxonómica y servir como punto de referencia en estudios de variación entre poblaciones, principalmente de P. labialis y P. candalarius, que presentan distribuciones amplias, entre las cuales es probable la existencia de otras especies.
La construcción de formas del cuerpo a partir de cuadriláteros contiguos, además de ser útil en la búsqueda de caracteres discriminantes, desde el punto de vista ecomorfológico, podría ser una herramienta importante para entender y explicar algunos de los procesos ecológicos que favorecieron o determinaron las características morfológicas de estas especies (Matthews, 1998). Esto adquiere valor si se considera que muchas de las diferencias observadas están asociadas con la altura y perfil del cuerpo, así como, a la posición y tamaño de la boca.
Aparentemente la morfología de las especies está siendo definida por factores históricos, como el aislamiento geográfico y las relaciones de parentesco, pero las evidencias morfométricas encontradas sugieren la existencia de un componente adaptativo, probablemente vinculado con la captura del alimento y la selección del hábitat, que quizá esté reforzando las diferencias entre las especies del grupo.
Agradecimientos
Al M. en C. Ernesto Velázquez Velázquez y la Universidad de Ciencias y Artes de Chiapas por facilitar el material de Profundulus candalarius. A Celedonio Chan Sala y Yamín de Vega López por su ayuda en la captura del material biológico.
Literatura citada
Álvarez, J. 1970. Peces Mexicanos (Claves). Serie de Investigaciones Pesqueras, Instituto Nacional de Investigaciones Pesqueras. México. 166 p. [ Links ]
Catena, A., Ramos, M. M. y H. M. Trujillo. 2003. Análisis Multivariado. Un manual para investigadores. Biblioteca Nueva, S. L. Madrid. 413 p. [ Links ]
Crisci, J. V. y M. F. López-Armengol. 1983. Introducción a la Teoría y Práctica de la taxonomía numérica. Organización de Estados Americanos. Washington, D. C. 132 p. [ Links ]
De la Cruz, A. G. 1994. ANACOM. Sistema para el ANAlisis de COMunidades. Versión 3.0 CICIMAR. IPN. México. 99 p. [ Links ]
Doadrio, I., Carmona J. A., Martínez E. y A. De Sostoa. 1999a. Genetic variation and taxonomic analysis of the subgenus Profundulus. Journal of Fish Biology 55: 751- 766. [ Links ]
Espinosa, P. H., M. T. Gaspar D. y P. Fuentes M. 1993. Listados faunísticos de México. III: Los peces dulceacuícolas de México. Instituto de Biología, UNAM. México. 98 p. [ Links ]
González, D. A. A. 2001. Osteología comparada del aparato branquial en especies del género Chirostoma Swainson (Pisces: Atherinopsidae). Tesis de maestría. Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional. México, D.F. 78 p. [ Links ]
Hubbs, C. L. y K. L. Lagler. 1958. Fishes of the Great Lakes Region. Second edition. Canbrook Institute of Science Bulletin 26:1- 213. [ Links ]
Matthews, W. J. 1998. Patterns in freshwater fish ecology. Second edition. Chapman and Hall. USA. 756 p. [ Links ]
Miller, R. R. 1950. Profundulus hildebrandi, a new cyprinodontid fish from Chiapas, Mexico. Copeia 1:22- 30. [ Links ]
Miller. R. R. 1955. A systematic review of the Middle American fishes of the genus Profundulus. Miscellaneous Publications Museum of Zoology, University of Michigan 92:1- 64. [ Links ]
Sneath, P. H. A. y R. R. Sokal. 1973. Numerical taxonomy. The principles and practice of numerical clasification. Freeman. San Francisco. 573 p. [ Links ]
Soria, B. M. 1999. Morfometría comparada del aparato mandibular en las especies de Chirostoma del Lago de Pátzcuaro, Mich. Tesis de licenciatura. Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional. México, D.F. 88 p. [ Links ]
Strauss, R. y L. F. Bookstein. 1982. The truss: body form reconstructions in morphometrics. Systematic Zoology 31: 113- 135. [ Links ]

