










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkTropical and subtropical agroecosystems
versão On-line ISSN 1870-0462
Trop. subtrop. agroecosyt vol.14 no.3 Mérida Set./Dez. 2011
Artículos de investigación
Componentes de (CO) varianza para peso al destete de ganado cebú bermejo cubano
(CO) Variance components of weaning weight in cuban zebu bermejo cattle
Narciso Ysac Ávila-Serrano1, Alejandro Palacios-Espinosa2*,José Luis Espinoza-Villavicencio2, Ariel Guillen-Trujillo2,Rafael de Luna-De la Peña2, Danilo Guerra-Iglesias3
1 Universidad del Mar, Puerto Escondido, San Pedro Mixtepec, Juquila, Oaxaca,71980, México.
2 Universidad Autónoma de Baja California Sur, carretera al sur km. 5.5 La Paz, B.C.S., 23080, México. E-mail: palacios@uabcs.mx
3 Centro de Investigaciones para el Mejoramiento Animal de la Ganadería Tropical, Ave. 101, Km 21.5, C de La Habana, Cuba.
* Corresponding Author
Submitted March 11, 2011
Accepted May 30, 2011
Revised received July 15, 2011
Resumen
Con el objetivo de estimar los componentes de (co)varianza del peso al destete en el ganado Cebú bermejo cubano se tomó una muestra de 5120 registros de animales hembras y machos que nacieron entre los años 1988 y 2007 en tres hatos ubicados en las provincias de Cienfiiegos, Camagüey y Granma. Se utilizaron seis modelos estadísticos diferentes que tenían en común el efecto fijo de grupo de contemporáneo (año-hato-época-sexo) y las covariables edad al destete lineal, edad de la madre al parto lineal y cuadrático. El modelo 1 sólo incluyó el efecto aleatorio del animal, el modelo 2 incluyó además el efecto aleatorio del ambiente permanente, el modelo 3 fue igual que el 1 pero incluyó además el efecto aleatorio genético materno, sin considerar la existencia de covarianza entre los efectos aditivos y maternos, el modelo 4 fue igual que el 3 pero consideró la existencia de covarianza entre efectos aditivos y maternos, los modelos 5 y 6 fueron iguales a los modelos 3 y 4, respectivamente, pero adicionando el efecto aleatorio del ambiente permanente. El peso al destete promedio fue de 159.7 kg, con una desviación estándar de 26.7 kg. Se utilizaron tres criterios para comparar los modelos; el logaritmo de la verosimilitud (log L), el criterio informativo de Akaike (AIC) y el criterio informativo de Bayes (BIC). El modelo 1 fue el de peor ajuste considerando los tres criterios. Teniendo en cuenta el criterio log L los modelos 5 y 6 no difieren significativamente entre ellos y resultaron los mejores con relación al resto. Los criterios AIC y BIC señalan al modelo 5 como el mejor. La heredabilidad para el efecto aditivo directo (0.32) en el modelo 1 fue superior a la estimada para con el resto de los modelos (0.02-0.06), lo que indica que ignorar los efectos maternales produjo una sobreestimación de este parámetro. Las heredabilidades estimadas para los efectos maternos en los modelos de mejor ajuste alcanzaron valores de 0.06 (modelo 5) y 0.04 (modelo 6). Los efectos del ambiente permanente determinaron entre un 10.9% (modelo 6) y 15.4% (modelo 3) de la variabilidad total. La correlación genética entre los efectos directos y maternos alcanzó valores de 0.53 y 0.61 para los modelos 4 y 6, respectivamente. Los resultados indican que en la presente población, existen pocas posibilidades de mejorar el peso al destete por selección.
Palabras claves: Ganado Cebú; parámetros genéticos; componentes de varianza; efectos maternales; peso al destete.
Abstract
The objective of this study was to estimate the (co) variance components of weaning weight in the Cuban bright reddish Zebu cattle a sample of 5120 records of male and female animals born between 1988 and 2007 was taken from three herds located in the provinces of Cienfiiegos, Camagüey and Granma. Six different statistical models were used which had in common as fixed effects the contemporary groups (year-herd-season-sex) and the covariables, linear weaning age and mother's age at linear and quadratic calving. Model 1 only included the random effect of the animal, model 2 also included the random effect of permanent environment, model 3 was equal to 1 but also including the random maternal genetic effect, regardless of the existence of covariance between the additive and maternal effects, model 4 was equal to 3 but considering the existence of covariance between additive and maternal effects, models 5 and 6 were the same as models 3 and 4, respectively, but adding the random effect of permanent environment. Weaning weight averaged 159.7 kg, with a standard deviation of 26.7 kg. Three criteria to compare models were used the log-likelihood (log L), Akaike information criterion (AIC) and Bayesian information criterion (BIC). Model 1 was that of worst fit considering the three criteria. Taking into account the criterion log L models 5 and 6 did not differ significantly between them and were the best compared to the rest. AIC and BIC criteria indicate the model 5 as the best. Heritability for the direct additive effect (0.32) in modelo 1 was higher than that estimated for the other models (0.02 to 0.06). Heritabilities estimated for maternal effects in models of best fit reached values of 0.06 (model 5) and 0.04 (model 6), which although higher than those of the direct effect are also low.
Permanent environmental effects determined between 10.9% (model 6) and 15.4% (model 3) of the total variability. The genetic correlation between direct and maternal effects reached values of 0.53 and 0.61 for models 4 and 6, respectively. The results indicate that there is little possibility of improving the weaning weight by selection in the present population.
Key words: Zebu cattle; genetic parameters, variance components; maternal effects; weaning weight.
INTRODUCCIÓN
El ganado Bos indicus, originario de la India y Pakistán (Sanders, 1980) está adaptado a las condiciones tropicales y subtropicales (Campher et al, 1998) y puede desarrollarse en ambientes difíciles que limitan la producción de carne de razas especializadas y constituyen un reto para el mejoramiento genético. La importancia que para la ganadería tropical y especialmente para Cuba, ha tenido y tendrá la raza Cebú de origen Brahaman, ha sido destacada por Planas et al. (2004). Esta raza se encuentra distribuida por todo el país y existen reportes en los últimos años que demuestran su potencialidad (Planas y Ramos, 1994).
El peso al destete es uno de los caracteres más importantes a considerar en los programas de mejoramiento genético del ganado vacuno de carne de ahí que sea uno de los más estudiados (Bertrand y Benyshek, 1987; Cantet et al, 1988; Meyer, 1992; Waldron et al., 1993; Robinson, 1996; Meyer, 1997; Dodenhoff et al., 1999; Quintanilla y Piedrafita, 2000; Bijma, 2006).
El peso al destete está influido por los efectos maternales que deben ser considerados conjuntamente con los componentes genéticos directos, especialmente si existe una relación antagónica entre ellos (Baker, 1980; Bertrand y Benyshek, 1987). El conocimiento de estos componentes de varianza es necesario en el diseño de los programas de mejora y para la estimación del valor genético de los animales. La estimación de los efectos maternales ha sido siempre problemático, debido a que los efectos directos y maternos están generalmente confundidos. Quass y Pollack (1980), basados en el modelo biométrico de Willham (1963) desarrollaron la inclusión de los efectos maternos en el modelo animal, método que hasta ahora ha sido el más utilizado para la estimación de los efectos maternales en ganado de carne. Algunos métodos para estimar componentes de (co)varianza para caracteres influidos por efectos maternales en ganado de carne han sido propuestos también por Meyer (1992; 1997).
En Cuba existen ties variedades de ganado Cebú registrado bajo control: el blanco, el bermejo o rojo y el sardo. Estas variedades se crían de manera independiente. Los trabajos de investigación han estado dirigidos a la estimación de componentes de varianza y tendencias genéticas en el Cebú blanco (Rico y Planas, 1994; Guerra et al, 2002; Menéndez et al, 2006) y hasta ahora no existen estimaciones de los componentes de (co)varianza para el Cebú bermejo. El objetivo del presente trabajo fue estimar los componentes de (co)varianza del peso al destete en ganado Cebú bermejo cubano, mediante la utilización de seis modelos animal que alternan los efectos maternales.
MATERIALES Y MÉTODOS
Se analizó el peso al destete (PD) a los 7 meses de edad de 6321 terneros hembras y machos de la raza Cebú bermejo que nacieron entre los años 1988 y 2007 en ties hatos ubicados en las provincias de Cienfuegos, Camagüey y Granma. La información fue suministrada por el Centro Nacional de Control Pecuario (CENCOP). En la edición de los datos se eliminaron aquellos grupos de contemporáneas (hato-año-cuatrimestre de nacimiento-sexo de la cría) con menos de cinco observaciones. También se desechó la información de los individuos con madre desconocida y todos aquellos que carecieran de la información relacionada con la fecha de nacimiento, edad de la madre, hato y edad. Finalmente, la muestra quedó conformada por 5120 registros. La Tabla 1 presenta una descripción general de la muestra analizada. La estructura de los datos cumple con las recomendaciones mínimas de 10% de madres con datos propios como becerras y más de 1.5 crías por vaca, para la correcta partición de los efectos maternos y ambientales permanentes (Maniatis y Pollot, 2003). Estructuras similares han sido consideradas como adecuadas para la evaluación genética del peso al destete por Diaz et al. (2002) y Cinachi et al. (2006), en ganado Limousine Español y en ganado Caracú Brasileño, respectivamente.

Los datos fueron analizados mediante el programa 'WOMBAT' de Meyer (2006) utilizando seis modelos matemáticos. Como efectos fijos comunes a todos los modelos se consideraron al grupo de contemporáneos, la edad de la cría al destete como covariable lineal y la edad de la madre como covariable lineal y cuadrática. El modelo 1, fue un modelo animal simple con los efectos genéticos aditivos como único efecto aleatorio. El modelo 2 fue igual que el 1 pero adicionándole los efectos del ambiente permanente debidos a la madre y no correlacionado con los efectos aditivos en el modelo. El modelo 3 fue como el 1 pero adicionando los efectos maternales genéticos y asumiendo una covarianza igual a cero entre los efectos directos y maternos. El modelo 4 fue igual al 3 pero permitió una covarianza entre los efectos directos y maternos. Los modelos 5 y 6 fueron similares al 3 y 4, respectivamente, pero adicionando los efectos del ambiente permanente. En forma matricial estos modelos se representan de la siguiente forma:
Modelo 1: y = Xb + Za a+ e
Modelo 2:y = Xb + Zaa + Zcc + e
Modelo 3:y = Xb + Zaa + Zmm + e con cov(a, m) = 0
Modelo 4: y = Xb + Za a + Zm m + e con cov(a, m) ≠ 0
Modelo 5: y = Xb + Za a + Zm m + Zc c + e con cov(a, m) = 0
Modelo 6: y = Xb + Za a + Zm m + Zc c + e con cov(a, m) ≠ 0
En donde:
y = es el vector de las observaciones del peso al
destete.
b = vector de los efectos fijos que incluye el efecto del grupo contemporáneo y la regresión (lineal y cuadrática) de la edad de la madre y de la edad de la cría al destete (lineal) sobre el PD.
a = vector de los efectos aleatorios genéticos aditivos.
m = vector de los efectos aleatorios genéticos maternos.
c = vector de los efectos aleatorios del ambiente permanente.
e = vector de los efectos aleatorios residuales.
X, Zd, Zm,y Zc son las matrices de incidencia que relacionan las observaciones con los efectos fijos, aditivos directos, aditivos maternos y del ambiente permanente, respectivamente.
El primero y segundo momento de los componentes aleatorios para el modelo 6 que incluyó todos los efectos son: [ a' m' c' e' y'] ~N {[0'0'0'0' (Xb)']', Σ}
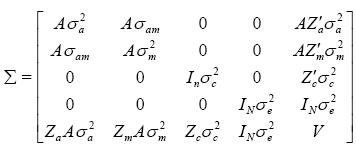
Donde A es la matriz de relaciones aditivas entre todos los individuos en el pedigrí, I es la matriz identidad, n es el número de madres, N es el número total de observaciones 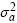 es la varianza genética aditiva de los efectos directos,
es la varianza genética aditiva de los efectos directos,  es la varianza genética aditiva de los efectos maternales,
es la varianza genética aditiva de los efectos maternales,  es la covarianza genética aditiva entre los efectos directos y los efectos maternales,
es la covarianza genética aditiva entre los efectos directos y los efectos maternales, 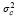 es la varianza de los efectos del ambiente permanente,
es la varianza de los efectos del ambiente permanente,  es la varianza del error y
es la varianza del error y
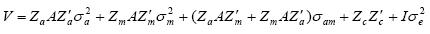
Para determinar el modelo más apropiado se utilizó la prueba del logaritmo de la verosimilitud (Meyer, 1992):

donde log Lr y log Lc son los logaritmos de la verosimilitud para el modelo reducido y completo, respectivamente. En esta prueba, la distribución Chi-cuadrada tiene grados de libertad igual al número de parámetros en el modelo completo menos el número de parámetros en el modelo reducido. También se utilizaron el criterio informativo de Akaike (Akaike, 1974). El objetivo es buscar el modelo aproximado, partiendo del modelo completo, que proporcione la menor pérdida de información posible. La expresión de AIC es - 2 log L+ 2 p, donde L es la función de verosimilitud y p es el número de parámetros independientes estimados en el modelo. La función de verosimilitud es una medida de la capacidad de ajuste del modelo, mientras que 2 p representa una penalizacion debida al número de parámetros (Sorensen y Gianola, 2002). El criterio informativo de Bayes (Schwarz, 1978), BIC= - 2 log L + p log (N - r), donde N es el número de observaciones y r es el rango de la matriz de incidencia para los efectos fijos. Estas pruebas (AIC y BIC) pueden ser usadas para comparar modelos con los mismos efectos fijos pero diferentes estructuras de (co)varianzas. Los valores más bajos son preferidos.
RESULTADOS Y DISCUSIÓN
En la Tabla 2 se presentan los diferentes criterios considerados para determinar la bondad de ajuste del modelo. El modelo 1 fue el que menos bondad mostró de conformidad con los tres criterios considerados, también presentó el mayor componente aditivo y del error. Teniendo en cuenta el criterio log L, los modelos 5 y 6 no difieren significativamente entre ellos y resultaron los mejores con relación al resto. Los modelos 3 y 4 tampoco presentaron diferencias significativas entre ellos. Bajo los criterios AIC y BIC el modelo 5 fue el de mejor ajuste. Este mismo modelo fue seleccionado bajo los mismos criterios por Domínguez et al. (2009) en un estudio realizado con ganado Salers.

En aquellos modelos donde se tuvieron en cuenta los componentes para los efectos maternales genéticos y/o maternales ambientales (modelos 2 al 6), toda la varianza materna fue considerada, coincidiendo con Meyer (1992), quien señala que cuando se considera uno solo de estos efectos (genético o ambiental), toda la variación maternal es tenida en cuenta. Lo anterior permite una mejor estimación de la heredabilidad genética aditiva, ya que esta misma autora demostró, que los modelos que no toman en cuenta los efectos maternales producen substancialmente estimaciones altas de la varianza genética aditiva directa y en consecuencia estimaciones altas de la heredabilidad genética aditiva. Lo anterior coincide con lo observado en el presente estudio, en donde en el modelo 1 que ignora efectos maternales, se presenta una estimación elevada de la varianza genética aditiva (Tabla 3) y en consecuencia una estimación elevada de la heredabilidad de los efectos genéticos directos (Tabla 4), coincidiendo con lo reportado por otros autores (Meyer, 1992; Domínguez et al., 2003; Prayaga y Henshall, 2005). De igual manera los criterios para determinar el ajuste de los modelos favorecieron a aquellos con una mayor influencia del componente ambiental materno que el componente genético materno (Tabla 3). Resultados similares fueron publicados por Prayaga y Henshall (2005) en cruzas de Cebú con razas británicas y continentales, criados en una estación experimental en Australia.
Con relación a los efectos del ambiente permanente (considerados en los modelos 2, 5 y 6), se observa que  y
y 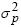 son muy similares y de valores más bajos que en el resto de los modelos (Tabla 3).
son muy similares y de valores más bajos que en el resto de los modelos (Tabla 3).
Los índices de herencia estimados por los modelos 2, 3 4, 5 y 6 (Tabla 4) oscilaron en un rango entre 0.023±0.017 (modelo 6) y 0.024±0.061 (modelo2). Estas estimaciones se consideran bajas, dejando pocas posibilidades para la mejora por selección para el peso al destete en las poblaciones estudiadas. Mercadante et al. (1995), reportan una heredabilidad superior para el efecto directo en ganado de carne de países tropicales. Por su parte Ríos (2008), en una revisión sobre el tema, en trabajos publicados entre los años 1940 y 2006, señaló un valor promedio no ponderado de 0.27 para la heredabilidad de los efectos directos en todas las razas y de 0.26 para la Brahman y la Nelore. Sin embargo, también se encuentran en la literatura valores similares a los estimados en el presente estudio (Elzo et al, 2001; Plasse et al, 2002; Nuñez-Dominguez et al., 2006).
Las heredabilidades estimadas para los efectos maternos (Tabla 4) en los modelos de mejor ajuste alcanzaron valores de 0.059±0.023 (modelo 5) y 0.045±0.022 (modelo 6), las cuales, aunque mayores que las del efecto directo también resultan bajas. Ríos (2008) en su revisión destaca un valor promedio de 0.17 para todas las razas, de 0.13 para Brahman y 0.12 en Nelore. Sin embargo, Pico et al. (2004) y Nuñez-Dominguez et al. (2006) encontraron valores de heredabilidad para los efectos maternales, similares a los del presente trabajo.
Como se observa en la Tabla 4 los efectos del ambiente permanente determinaron entre un 10.9% (modelo 6) y un 15.4% (modelo 3) de la variabilidad total. Estos valores coinciden con los promedios publicados por Ríos (2008) del 14% para todas las razas y del 12% para Brahman y Nelore.
La correlación genética entre los efectos directos y maternos se tuvo en cuenta en los modelos 4 y 6, alcanzando valores de 0.534 ±0.425 y 0.614±0.557, respectivamente. Sin embargo, entre los modelos 3 y 4 y los modelos 5 y 6 no existieron diferencias significativas y en virtud de que la única diferencia entre ellos es la covarianza genética entre los efectos directos y maternos, podría considerarse que la inclusión de dicha covarianza no es importante. Sin embargo, debido a los altos errores estándar obtenidos, lo anterior debe ser considerado con precaución. Los estimados de la correlación genética entre los efectos directos y maternos que aparecen en la literatura difieren entre ellos, correlaciones negativas (-0.68, -0.37, -0.46 ) han sido reportadas en otros estudios (Pimenta et al, 2001; Domínguez et al, 2003, Estrada et al, 2008, respectivamente). Muchos de los estimados altos y negativos han sido atribuido al síndrome de las ubres grasas (Koch, 1972; Willham, 1972). Heydarpour et al. (2008) señalaron que una estructura poblacional pobre podría generar sesgos en los estimados de la correlación genética entre los efectos directos y maternos y explicar algunos de los estimados altos y negativos frecuentemente obtenidos, así también, cuando las correlaciones son calculadas sobre datos obtenidos en rebaños de universidades o estaciones experimentales donde existe un buen control de la genealogía estas son pequeñas y positivas, pero en el caso de datos de campo donde se pierde la conexión entre la identificación del animal cuando se desteta y cuando es madre, estas correlaciones tienden a ser fuertemente negativas en dependencia del porcentaje de pérdida de la información.
CONCLUSIONES
La exclusión de los efectos maternales genéticos y/o ambientales provocó una sobreestimación del componente aditivo directo. Sin embargo, cuando estos efectos fueron considerados, las heredabilidades resultaron bajas y consecuentemente con pocas posibilidades de respuesta a la selección. La variabilidad de los efectos maternos resultó superior a la de los efectos aditivos directos, fundamentalmente la ambiental. Las correlaciones genéticas entre los efectos directos y maternos fueron altas y positivas pero no resultaron significativas. Los resultados indican que existen pocas posibilidades de mejorar el peso al destete por selección en la presente población.
REFERENCIAS
Akaike, H.A.1974. New look at statistical model identification. Institute of Statistical Mathematics, Minato-Ku, Tokyo, Japan. IEEE Trans. Automat. Contr. AC-19:716-723. [ Links ]
Baker, R.L. 1980. The role of maternal effects on the efficiency of selection in beef cattle: A review. Proceedings of the New Zealand Society of Animal Production. 40, 285-303. [ Links ]
Bertrand, J.K. and Benyshek, L.L. 1987. Variance and covariance estimates for maternally influenced beef growth traits. Journal of Animal Science. 64:728-734. [ Links ]
Bijma, P. 2006. Estimating maternal genetic effects in livestock. Journal of Animal Science. 84: 800-806. [ Links ]
Campher, J.P., Hunlun, C. and Van Zyl, G.J. 1998. South African Livestock Breeding. South African Stud Book and Livestock Improvements Association, Bloemfontein. p56-57. [ Links ]
Cantet, R.J.C., Kress, D.D., Anderson, D.C., Doornbos, D.E., Burfening, P.J. and Blackwell, R.L. 1988. Direct and maternal variances and covariances and maternal Phenotypics effects on preweaning growth of beef cattle. Journal of Animal Science. 66: 648-660. [ Links ]
Cinachi, P.M., Mercadante, M.E.Z., Galao, L., Razook, A.G. e Andrade, L. 2006. Estimativas de parámetros genéticos de características de crecimiento em urn rebanho Caracú seleccionado para peso ao sobreano. Revista Brasileira de Zootecnia. 3: 1669-1679. [ Links ]
Diaz, C, Moreno, A. and Carabaño, M.J. 2002. Effects of genetic groups on estimates of variance components for maternally influence traits in Spanish Limousine. Journal of Animal Breeding and Genetics. 119: 325-334. [ Links ]
Dodenhoff, J., Van Vleck, L.D. and Wilson, D.E. 1999. Comparison of models to estimate genetic effects for weaning weight of Angus cattle. Journal of Animal Science. 77: 3176-3184. [ Links ]
Domínguez V.J., Nuñez, D.R., Ramírez, V.R. y Ruiz, F.A. 2003. Evaluación genética de variables de crecimiento en bovinos Tropicarne : Selección de modelos. Agrociencia. 37: 323-335. [ Links ]
Domínguez V.J., Rodríguez, A.F., Ortega, G.J.A. y Flores, M.A. 2009. Selección de modelos, parámetros genéticos y tendencias genéticas en las evaluaciones genéticas nacionales de bovinos Brangus Salers. Agrociencia. 43: 107-117. [ Links ]
Elzo, M.A., Martinez, G., Gonzalez, F. y Huertas, H. 2001. Variabilidad y predicciones genéticas aditivas, no aditivas y totales para la producción de Ganado de carne en el rebaño multiracial Sanmartinero-Cebú de La Libertad. R. Corpoica. 3: 51-64. [ Links ]
Estrada, L.R.J., Monforte, J. y Segura, C.J.C. 2008. Comparación de modelos en la evaluación genética de caracteres de crecimiento del ganado Brahman en el sureste de México. Archivos Latinoamericanos de Producción Animal. 4:221-231. [ Links ]
Guerra, D., Rodriguez, M., Ramos, F., Planas, T., Ortiz, J., Torres, J.A. and Falcon, R. 2002. Evaluación genética de las razas de carne en Cuba. II Congreso Internacional de Mejora Animal. La Haba, Cuba. 561; 4. [ Links ]
Heydarpour, M., Schaeffer, L.R. and Yazdi, M.H. 2008. Influence of population structure on estimates of direct and maternal parameters. Journal of Animal Breeding and Genetics. 125: 89-99. [ Links ]
Koch, R.M. 1972. The role of maternal effects in animal breeding. VI Maternal effects in beef cattle. Journal of Animal Science. 35: 1316-1323. [ Links ]
Maniatis, N. and Pollot, G.E. 2003. The impact of data structure on genetic (co)variance components of early growth in sheep, estimated using an animal model with maternal effects. Journal of Animal Science. 81: 101-108. [ Links ]
Menendez, A., Guerra, D., Planas, T. and Ramos, F. 2006. Factors affecting the growth of young bulls of the Zebu breeds in the performance test in grazing conditions. Cuban Journal of Agriculture Science. 40: 371-377. [ Links ]
Mercadante, M.E.Z., Lobo, R.B. and De los Reyes, A. 1995. Parámetros genéticos para características de crecimiento en cebuinos de carne. Archivos Latinoamericanos de Producción Animal. 3: 45-89. [ Links ]
Meyer, K. 2006. 'WOMBAT'. Digging deep for quantitative genetics analyses by restricted maximum likelihood. Proc. 8th World Congress on Genetics Applied to Livestock Production. Belo Horizonte, Brasil. [ Links ]
Meyer, K. 1997. Estimates of genetic parameters for weaning weight of beef cattle accounting for direct-maternal environmental covariances. Livestock Production Science. 52: 187-199. [ Links ]
Meyer, K. 1992. Variance components due to direct and maternal effects for growth traits of Australian beef cattle. Livestock Production Science. 31: 179-204. [ Links ]
Núñez-Domínguez R., Ramírez, V.R., Hernández, A.O. Ruiz, F.A., Domínguez, V.V. and García, M.J.G. 2006. Single vs. Bivariate analisys for the genetic evaluation of groth traits in Mexican cattle populations. In: Proc. 8th World Congress on Genetics Applied to Livestock Production. Belo Horizonte, Minas Gerais, Brasil, Comm. No. 03-67. [ Links ]
Pico, B.A., Neser, F.W.C. and Van-Wyk, J.B. 2004. Genetic parameters for growth traits in South African Brahman cattle. S. Afr. Journal of Animal Science. 34: 44-46. [ Links ]
Pimenta, F.E., Martins, A.G., Sarmentó, R.J., Ribeiro, M.N., e Martins, F.R. 2001. Estimativas de heredabilidade de efeitos directo e materno de características de crecimiento de bovinos Guzerá no estado de Paraiba. Revista Brasileira de Zootecnia. 30: 1220-1223. [ Links ]
Planas, T., y Ramos, F. 2004. Cebú Cubano. Origen y principales resultados. Revista de la Asociación Cubana de Producción Animal. 1: 10-18. [ Links ]
Planas, T., Rico, C, Ribas, M., Pérez, T., Castro, H., Pérez, M., López, D. y Ponce de León, R. 2004. La genética en manos del criador. Cap. 1, 11-20. [ Links ]
Plasse, D., Verde, O., Fossi, H., Romero, R., Hoogesteijn, R., Bastidas, P. and Bastardo, J. 2002. (Co)variance components, genetic parameters and annual trends for calf weights in a pedigree Brahman herd under selection for three decades. Journal of Animal Breeding and Genetics. 119: 141-153. [ Links ]
Prayaga, K.C. and Henshall, J.M. 2005. Adaptability in tropical beef cattle: genetic parameters of growth, adaptive and temperament traits in a crossbred population. Australian Journal of Experimental Agriculture. 45: 971-983. [ Links ]
Quaas, R.L and Pollack, E.J. 1980. Mixed model methodology for farm and ranch beef cattle testing programs. Journal of Animal Science. 51: 1277-1287. [ Links ]
Quintanilla, R. y Piedrafita J. 2000. Efectos maternos en el peso al destete del ganado vacuno de carne: Una revisión. ITEA 96: 7-39. [ Links ]
Rico, C. y Planas, T. 1994. Influencias ambientales y genéticas en el desarrollo de bovinos Cebú. Revista Cubana de Ciencias Agrícolas. 28: 265-272. [ Links ]
Ríos, A. 2008. Estimadores de parámetros genéticos para características de crecimiento predestete de bovinos. Revisión. Técnica Pecuaria en México. 46: 37-67. [ Links ]
Robinson, D.L. 1996. Estimation and interpretation of direct and maternal genetic parameters for weights of Australian Angus cattle. Livestock Production Science. 45: 1-11. [ Links ]
Sanders, J.O. 1980. History and development of Zebu cattle in the United States. Journal of Animal Science. 50: 1188-1200. [ Links ]
Schwarz, G. 1993. Estimating the dimension of a model. Annals of Stat. 1978: 6:461-464. [ Links ] Waldron DF, Morris CA, Baker RL, Johnson DL. Maternal effects for growth traits in beef cattle. Livestock Production Science. 34: 57-70. [ Links ]
Sorensen, D. and Gianola, D. 2002. Likelihood, Bayesian and MCMC Methods in Quantitative Genetics. Springer. New York, U.S.A. 742 p. [ Links ]
Willham, R.L. 1963. The covariance between relatives for characters composed of components contributed by related individuals. Biometrics. 19: 18-27. [ Links ]
Willham, R.L. 1972. The role of maternal effects in animal breeding. Ill Biometrical aspects of maternal effects. Journal of Animal Science. 35: 1288-1293. [ Links ]

