











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink1 Introduction
Tuna species (Thunnus albacares) are extensively processed worldwide giving rise to a large amount of unused by-products. These by-products are composed of viscera, skin, gills, head, bones and dark meat. Despite good nutritional characteristics, dark or red meat have no commercial value due to their unpleasant taste (Graham and Dickson 2004). Several methods have been reported to draw on these by-products; some are based on chemical hydrolysis, regardless of poor final product quality, and others by biological means including hydrolysis by exogenous or endogenous enzymes in mild process conditions (Pihlanto and Korhonen 2003; Chi et al.,2015; Reyes-Mendez et al.,2015). These bio-based methodologies offer ease of control of the reaction and optimal recovery of added-value products without side products (Shirai and Ramirez-Ramirez 2011). Among them, lactic acid fermentation (LAF) proved successful owing to process stability, inhibition of undesirable bacterial growth and the production of biopolymers, valued organic acids and hydrolysates in a GRAS and chemical-free process with low energy requirement (Ramírez-Ramírez et al., 2008; Flores-Albino et al., 2012). These fermentations are widely used to process small pelagic fish or fish by-products, e.g. sauce production in heavily salted conditions. In this regard, some mechanisms have been put forth to describe antioxidant activities of peptides, including ion-chelating capacity, radical scavenging and aldehyde adduction (Mishra et al., 2011). The feed for the lactic acid bacteria (LAB) covers a wide range of substrates; nonetheless the papaya (Carica papaya) is pointed out due to its high protease content and additionally, it is intensively cultivated throughout the year in tropical countries owing to its high demand as fresh fruit and ingredient in food processing (Fabi et al., 2007). On the other hand, the lactic acid (LA) produced in situ has a significant role in fish conservation and antimicrobial effect against undesirable microflora. Generally, the efficacy of LAF is based on the rate of pH drop and its final value, which rely on water activity, temperature and process time, activity of starter and buffer capacity. Practical utilization of fish by-products by biotechnological means requires information on the buffer capacity, endogenous protease activities, and ash and protein contents. In this regard, the bioactive peptide production from tuna wastes by LAF with papaya and molasses as carbon sources has not been reported to the best of our knowledge and the present work establishes the conditions for efficient bioprocess production of antioxidant protein hydrolysates.
2 Materials and methods
2.1 Materials
Lactobacillus plantarum (APG-Eurozym) was cultivated in Man, Rogosa and Sharpe (MRS) agar at 30 °C and kept at 8 °C prior to use. Inoculum was prepared in MRS broth by inoculation from the bacterial slants and incubated at 30 °C for 24 h to reach a cell count of 108 colony-forming units per millilitre (CFU/mL). Yellowfin tuna (Thunnus albacares) wastes were obtained from a canned tuna processing plant in Mazatlan (Sinaloa State, Mexico). The wastes consisted of three factions: i) fresh viscera (V), ii) heads, bones and fins (HBF) and iii) dark meat (DM). The fish were manually gutted and abdominal cavity was washed with water. Eviscerated fish was steam cooked at 100 °C for 3 h. HBF and DM were separated from fish. Each fraction was minced separately through a 5 mm sieve using a meat mincer (Torrey 32-3, Mexico) and stored at -20 °C. Sugar cane molasses were supplied by sugar mill "El Molino S.A. de C.V." in Nayarit state (Mexico). Papaya fruit (Carica papaya L. var Maradol) at commercial maturity (skin yellowed and orange soft pulp) was harvested in the state of Veracruz (Mexico) and milled in a blender prior to use. Soluble sugars in molasses and papaya were determined by the method of phenol sulphuric acid (Dubois et al 1956). Reagents 2,2-diphenyl-1-picrylhydrazyl (DPPH), acrylamide, bisacrylamide, sodium dodecyl sulphate, Tris base, Tricine, glycerol and ultra-low molecular weight marker (1060-26600 Da) were supplied by Sigma-Aldrich (USA). Electrophoresis grade Coomasie Blue G-250, 2-mercaptoethanol and TEMED were supplied by Bio-Rad (USA).
2.2 Determination of tuna wastes proportions in the mixture, starter level and papaya concentration for LAF
Tuna wastes in LAF varied upon the ratio of V and HBF (wt/wt%): 64.9:7.2; 43.8:28.3; 33.3:39.3; 14.4:42.3; 7.2: 64.9. DM was kept at 27.9 wt/wt% considering that this fraction presented the highest protein content. Mixtures were analyzed on their proteases activities (Bougatef et al., 2009) and ash content (AOAC 1997). Five grams of tuna waste mixtures were homogenized in 50 ml of deionized water. Homogenates were equilibrated at 25°C and stirred continuously during titrations with LA (0.054N) employing pH meter equipped with a glass electrode (pH 210 HANNA, Italy). Maximum bufer capacity (?max) was estimated as the slope of pH-neutralization curves in the immediate vicinity of pKa value (pKa of LA=3.86) according to equation 1 (Baicu and Taylor, 2002).
 (1)
(1)
dV is expressed in mmol/l, β max in slykes (mmol of LA required to titrate the pH of 1 g (wet wt) of tuna mixture by one pH unit). The first derivative f (X0) = |a/4b| is the slope of the tangent to the titration curve at pKa location. Limits of linearity are (y 2 - y 1 ) and (x2 - x1) of the pH-LA volume curve. Tangent and linear portion of the curve were overlapped and their slopes coincided.
The starter level was determined using 10 (wt/wt%) sugar cane molasses to the selected tuna wastes mixture and inoculated with 5,10 or 20 (v/wt% wet basis) of L. plantarum at 30 °C. The integrated Gompertz model was applied to analyze the kinetic data of LA production as function of time t according to the equation 2:
 (2)
(2)
where P max is the maximum LA produced (t → ∞), b is a constant related to the initial conditions (when t = 0, then P = Pmax exp(-b)) and k is the acidification rate constant. Kinetic constants P max , b and k were estimated by the non-linear estimation programme STATISTICA (StatSoft, Inc.). Maximum acid production rate (V max ) was calculated from parameters of the Gompertz model as V max = 0.368 kP max.
After the establishment of selected ratio of tuna wastes and inoculation level, several papaya concentrations (10, 20, 30 and 50 wt/wt%) were evaluated with 5 (wt/wt%) starter at 30 °C. Samples were withdrawn for analyses every 24 h and up to 120 h.
2.3 LAF in packed bed column reactor
LAF was conducted in a stainless steel column reactor placed in controlled temperature chamber (30 °C) and filled with 4 kg of the tuna wastes, 5 (v/wt%) of inoculum, 10 (wt/wt%) of molasses and 30 (wt/wt%) of papaya. After mixing, samples (200 g) were taken from the reactor every 24 h and up to 120 h for analyses. LAF samples were mixed in a 1:10 dilution with deionized water, homogenized at 3200 rpm for 2 min in an Ultraturrax ® T 25 homogenizer (IKA, Germany) and then centrifuged for 10 min at 15,000 g (4 °C). The pellet was discarded and the supernatant was filtered throughout 22 μm pore cellulose filter (Columbia Filter, Mexico).
2.4 Determination of pH, total titratable acidity, sugars and organic acids
The pH of samples was measured in a pH meter equipped with a glass electrode (pH 210 HANNA, Italy). Samples were diluted in distilled water (1:10) and total titratable acidity (TTA) was determined by titration with sodium hydroxide (0.1N) until a final pH of 7.5 and expressed as mmol of LA/g (Fabi et al., 2007). Sugars and organic acid were determined by HPLC according to the following methodology: samples were diluted with distilled water (1:10) and then homogenized by centrifugation for 20 min at 15,000 g (4 °C). Supernatants were analyzed in an Agilent 1260 chromatographer (USA) equipped with a refractive index detector and an Aminex® column (BioRad) using H2SO4 solution (5 mM) as mobile phase at 40 °C. Sucrose, glucose, fructose, LA and acetic acid concentrations were determined according to standard concentration curves.
2.5 Determination of degree of hydrolysis (DH) in protein hydrolysates
α-Amino acid contents were determined by the modified method of 2,4,6-trinitrobenzenesulfonic acid (TNBS) (Ramírez-Ramírez et al., 2008) using known concentrations of L -leucine and expressed as mM of α-amino acids. Percentage DH was calculated using the equation 3.
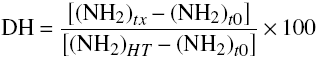 (3)
(3)
where: (NH2) tx is the amount of terminal ?-amino groups released at the time of fermentation t x ; (NH2) to is the amount of terminal a-amino groups released at the time of fermentation t 0 ; (NH2) HT is the amount of terminal α-amino groups without fermenting obtained after acid hydrolysis.
2.6 Microbiological analysis
Decimal dilutions in 0.9 (wt/v%) of sterile NaCl solution were prepared from samples in order to determine viable cell count in MRS and brilliant green bile (BD Bioxon, Mexico) agar plates for LAB and enterobacteria, respectively.
2.7 Determination of proteolytic activities
Activities of acidic and neutral proteases were determined employing hemoglobin and casein as substrates, respectively. Hemoglobin was dissolved in universal buffer (0.05 M) at pH 5 (Anson, 1938) and casein solution was prepared in phosphate bufer at pH 7 (Kunitz, 1947). A supernatant volume of 150 µL of was added to 1 mL of substrate (1 wt/v%) in its respective bufer and incubated for 1 h at 35 °C. The reaction was quenched by the addition of 1 mL of trichloroacetic acid (5 wt/v%), centrifuged and the supernatant absorbance measured at 280 nm. One unit of enzymatic activity (U) was defined as the amount of enzyme that produces an increment of 0.001 of absorbance units per min under the assay conditions.
2.8 Determination of radical scavenging activity (RSA)
Protein hydrolysate sample solutions were prepared and RSA were determined for electron scavenging activity using an Elexys Super X ® EPR spectrometer (Bruker, Germany) under the following procedure: different protein concentrations (20-1000 µg soluble protein/mg) were prepared in test tubes to which 2 mL of freshly prepared 0.1mM DPPH in methanol were added. Tubes were stirred in a vortex and incubated at 30 °C for 30 min in darkness. Then, sample was placed in a quartz flat cell to measure under 4 averaged scans, 100 kHz field modulation frequency, 9.78 GHz microwave frequency and 5 mW microwave power. Scavenging activity was calculated according to equation 4.
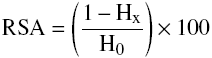 (4)
(4)
where H x and H 0 were the height on the spectra of the third resonance peak for samples with and without protein, respectively. RSA determinations were carried out by triplicate. Experimental data were fitted to Probit statistical analysis in NCSS ® programme (NCSS LLC. USA). IC50 value was the concentration of protein hydrolysates that inhibit 50% of DPPH radical scavenging activity (Bougatef et al., 2010).
2.9 Determination of molecular weight of protein hydrolysates by SDS-PAGE
LAF sample was suspended in 5 mL of acetic acid solution (0.087M) prior to injection in a Sep-Pak C-18 ® cartridge (Waters, USA) eluted with 5 mL acetonitrile solution (50%). Eluted samples were freeze dried and dissolved in bufer (12% SDS, 6% mercaptoethanol, 30% glycerol, 0.05% Coomassie blue G and 150mM TRIS HCl pH 7.0) and analyzed by denaturing electrophoresis SDS-PAGE. Gels were stained with coomassie blue (Bio-Rad, USA) and analyzed with the image processing software (ImageJ 1.41o National Institutes of Health, USA).
3 Results and discussion
3.1 Selection of dark meat:viscera:heads, bones and fins (DM:V:HBF) ratios in mixture ofTuna wastes for lactic acid fermentation
The proportion of fish waste fractions was determined upon protein, ash contents, bufer capacity and protease activity in the mixtures. The ash content increased consistently with the HFB (Figure 1), which might lead to raising the buffer capacity in fish fractions. Nevertheless, the highest bufer capacity was estimated in the mixture of 27.9:33.3:39.3 of DM:V:HBF, which also contained high protein content, 84.24 ± 2.96 % slykes. In this regard, DM presented the highest protein content (50.06 ± 1.12%) followed by V (46.22 ± 0.55%) and HBF (27.82+0.52%) (See supporting information 1 for maxima buffer capacities (?max) for mixture of Tuna wastes). The DM is a characteristic fish tissue that contains high myoglobin content, which has buffering capacity and, it enhances the diving capacity in marine animals against the accumulation of LA produced by muscle buffering under hypoxic conditions. Adequate buffering capacity is required since fast acidification during LAF can inhibit starter growth and negatively affect protease activity; however, highly-bufered substrate might slow down the pH decrease thereby favoring the development of spoilage microorganisms (Figure 1). Gongora and co-workers (2012) reported that Lactobacillus plantarum was able to reduce the pH to 4 in fish silage, however it presented inhibition after 383 min at pH 3 and inability to grow at pH 2. It is noteworthy that DM and HBF samples were previously cooked in the fishery and only V was raw, neutral protease activity was directly related to the amount of this ingredient in the mixture. The level of endogenous enzymes in the mixture with V of 33.3 wt/wt% was statistically grouped after those with V ratios of 43.8 and 64.9 wt/wt%, but higher than mixtures with V ratios of 7.21 and 14.42 wt/wt% (Figure 1). Therefore, based on the high buffering capacity and protein content, as well as active neutral digestive proteases the mixture of tuna wastes with 27.9 wt/wt% of DM, 33.3 wt/wt% of V and 39.3 wt/wt% of HBF was selected for further experimentation.
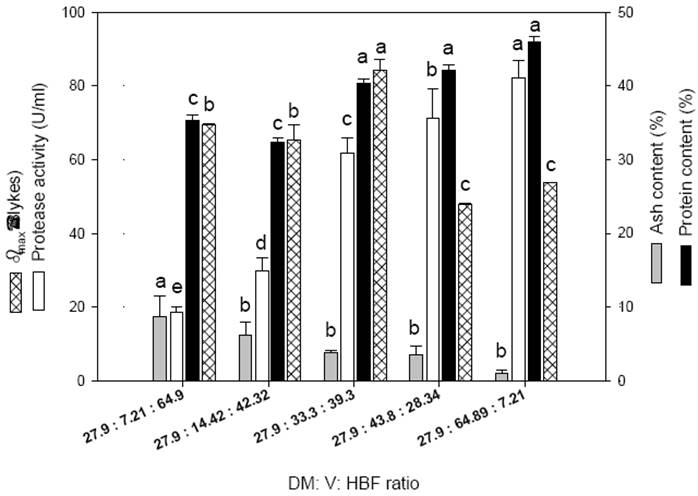
Fig. 1 Criteria employed for the selection of dark meat:viscera:heads, bones and fins (DM:V:HBF) ratios in mixture of tuna wastes for lactic acid fermentation expressed as maxima buffer capacities (βmax), proteases activities, protein and ash contents.
Table 1 Kinetic constants of lactic acid in Tuna by-products with added 10(wt/wt%) of molasses fermentation for production of protein hydrolysates with Lactobacillus plantarum as inoculum.
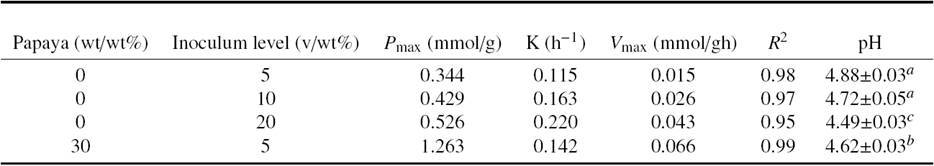
Values with different superscript letters in the same column showed significant differences (p < 0.05) according to Tukey-Kramer multiple comparison test.
3.2 Effect of inoculum level and papaya concentration on acidification in LAF
LA production (TTA) in tuna by-products fermentations with 10% of molasses and starter levels are shown in Figure 2. LA production presented significant differences (α < 0.95) at all tested levels of inoculum. The P max and V max increased with inoculation level, especially at 20%, whereas at 5% and 10% the acidification rate (k) increment was not significant, as well as pH after 120 h of fermentation. Therefore, 5 v/wt% of starter was chosen for further experimentation considering its cost in a scaled up process.

Fig. 2 Profile of acid production for fermentations and their adjustment to the Gompertz model with 10 (wt/wt%) molasses varying starter level (•) 5, (o) 10, (▼) 20 (v/wt%) and with 5 (wt/wt%) of inoculum and (△) with added 30 (wt/wt%) of papaya.
Noteworthy, the ripeness of papaya would not be a constraint when scaling up because the average sugar contents in ripe papaya (43.5 ± 0.39 mg sugars/g) determined in eight samples taken on March and December of 2007, 2012 and 2014 displayed no significant differences among batches, which is in agreement to similar studies by Gómez and co-workers (2002) that determined no significant differences between total soluble sugar of green (38.6 mg sugars/g), intermediate (43.8 mg sugars/g) and ripe (48.6 mg sugars/g) papaya. Therefore, the commercially disregarded fruits owing to sensory perceptions or low quality standards would be still of use in our LAF system. The papaya concentration and fermentation time had a significant effect on LA as shown in Figure 3. The combination of papaya level and time yielded the largest acid production. According to the results, papaya level increased LA production when time increased. The LAF carried out with 30 or 50 (wt/wt%) of fruit contained enough soluble sugars, however, at low papaya concentrations (10 and 20 wt/wt%), the TTA decreased with time, thus reaching a peak at 24 h that gradually fell toward the end of the fermentation. By contrast, high TTA (>0.6 mmol of LA/g LAF) values were attained in LAF at increased fruit content (Figure 3). The treatment with 50 wt/wt% of papaya displayed a steep decrease in pH and high TTA, although the latter decreased significantly after 96 h, therefore, according to the experimental data, 30 wt/wt% of papaya supplemented with 10 wt/wt% of molasses as carbon source (519.3 ± 64.61mg sugars/g molasses) ensured a stable acid production throughout fermentation. As shown in Figure 4, treatments with these favoring levels kept the pH near 4.6 with increase in the LA as a consequence of the lactobacilli growth which in turn, might inhibit spoilage such as hydrogen peroxide and bacteriocins as reported by Todorov et al. (2011). In the present work relatively high sucrose, fructose and glucose consumptions were observed in the first 24 h until reaching a stationary phase (Figure 4). In spite of using a homofermentative starter, acetic acid was also found to a small extent, which might be ascribed to heterofermentative endogenous microflora. Both organic acids displayed inhibitory effect on enterobacteria as it was determined by microbiological analyses, as expected, owing to the thermal pretreatment and the acidic conditions afterwards. The viable cell count of lactobacilli increases throughout the time course of the LAF (Figure 4b) concomitantly with the sugar consumption and LA production (Figure 4a), and additionally, glucose and fructose decreased to low concentrations toward the stationary phase.
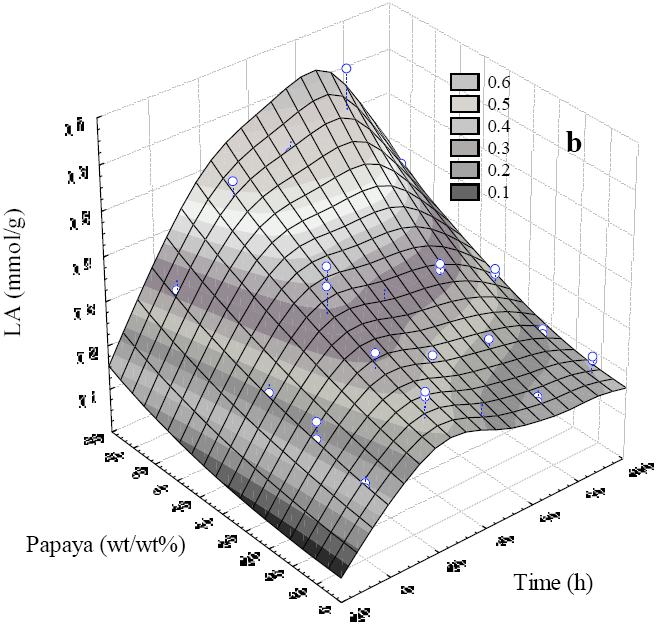
Fig. 3 Graphical 3D representation of time course of LA production of fermentations inoculated with 5 (wt/wt%) of L. plantarum and variations on papaya levels.

Fig. 4 LAF with papaya (30 wt/wt%) and molasses as carbon sources, Time course determinations of: a) sugars and organic acids determined by HPLC (♦) pH , (•) sucrose, (o) glucose, (▼) fructose, (blacksquare) LA and () acetic acid and b) (•) acidic, (o) neutral proteases, (■) DH and (×) lactic acid bacteria enumeration.
3.3 Determination of DH during fermentation
The DH determined during time course of LAF (Figure 4b) reached 89% at 120 h whereas reports of enzymatic protein hydrolysis in fish show lower values (4.7 to 30%), which might be explained by proteolytic activities observed during our LAF (Figure 4b). Acidic protease activity was high when the fermentation started but it decreased to null activity at 96 h. However, neutral protease activity remained throughout the fermentation, which resulted in an increase of DH (Figure 4b). Keeping in mind that protease activities in fish are mostly active from neutral to mild acidic conditions, this observed activity might be ascribed to trypsin-like and chymotrypsin-like digestive enzymes (Sriket, 2014). In spite of the acidic conditions during LAF, the activity of acidic proteases decreased at long reaction times that might be attributed to the cleavage by other proteases such as fish digestion enzymes, papain or microbial proteases. In this regard, pepsins found in tuna viscera might present activities under our LAF conditions in agreement to other studies (Simpson, 2010). The pepsins from marine animals were reported to have relatively higher optimum pH and stability range (pH 2.0-4.0) for hydrolysis than mammalian pepsins with a wide optimal temperature range (37-55 °C) and additionally, the papaya source presented neutral protease activity values of 0.767 (0.08 U/mg. According to Skelton (1968), papain from Carica papaya displays a temperature range from 25 °C (about 65% of maximal activity) to 70 °C (about 60% of maximal activity) with and optimum at 40 °C. In the present study, the pH ranges from 5 (70% maximal activity) to 8 (85% maximal activity) and the optimum varied from 6 to 6.5. The proteolytic activity from exogenous proteases with L. plantarum is reported at pH above 4.0, similar to that in LAF, with viable cell populations from 1.2 × 106 to 2.0 × 106 CFU/mL (Khalid and Marth, 1990), which is lower than the 108 CFU/mL determined in our LAF. This data provides further evidence of the relatively high activity of proteases from papaya and fish by-products. Finally, Figure 5 shows the electrophoretic profiles of initial time and LAF samples after 72 h, where high molecular weight proteins (> ca 260 kDa) were not detected but an increase of those with low molecular weights. The SDS-Tricine gels display protein bands below 8.7 kDa for the 96 and 120 h of LAF.

3.4 RSA of protein hydrolysates
The group with the highest RSA was that from 72 and 96 h with IC50 of 5.75 ± 1.04 μg/g and 7.6 ±1.7 μg/g, respectively, however, samples from 24 and 48 h displayed IC50 of 10.82 ± 1.9 and 9.06 ± 1.06 μg/g, respectively (See Supporting information 2 for EPR spectra in the range of concentrations tested for determination of IC50 by fitting RSA data to Probit model). In this regard, inherent proteins, melanoidins and polyphenols in fruit well might explain the observed RSA (IC50 of 23.24 ± 1.04 μg/g) at the beginning of LAF (Fig. 6b). However, RSA was retrieved by the release of hydrolysates from protein (Fig. 3c). Je et al. (2007) and Bougatef et al. (2010) also reported relatively high activities with low molecular weight peptides from fish, included in a cryptic form in the sequence of the precursor protein molecules and released after the hydrolysis therefore the contribution to RSA of other components in the LAF mixture in addition to the formed protein hydrolysates cannot be ruled out (Wang et al., 2008; Kumar et al., 2010; Zhang et al., 2012). Generally though, RSA in fish wastes has been attributed to proteins and peptides with increased hydrophobicity that displayed antioxidant properties as an important factor for increasing interaction with free radicals (Chalamaiah et al., 2012; Chi et al., 2015).

Conclusions
This work describes a detailed study on the important variables involved in the biotechnological process for exploitation of tuna wastes which can be used profitably toward production of added value protein hydrolysates, which besides nutritional value, could also present potential human and animal health promotion. The highest buffer capacity was estimated in the mixture of 27.9:33.3:39.3 of DM:V:HBF, which also contained high protein content, 84.24 ± 2.96 % slykes. In addition papaya represents a useful carbon source in tropical countries and therefore, the Lactobacillus plantarum fermentation of tuna using papaya fruit and molasses proved successful for this purpose. The potential scaling up of this biotechnological process is envisaged.

