











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkDeterminación de la actividad de ATPasa de la bomba cerca en homogenados de tejido muscular
Estudios en ratas con alimentación restringida han mostrado que la desnutrición induce cambios en el metabolismo y en el tipo de fibras que componen al músculo rápido EDL (extensor digitorum longus); lo cual se evidencia también por cambios en la expresión de diversas proteínas de la maquinaria contráctil, mitocondriales y del metabolismo glicolítico 1.
Por otro lado, en el síndrome metabólico también se presentan alteraciones en el músculo cardiaco que están relacionadas con la expresión y/o actividad de canales iónicos y bombas de Ca2+ como el receptor de rianodina 2 y la bomba ATPasa del retículo sarco/endoplámico (o bomba SERCA) 3. Ésta última es una ATPasa que bombea Ca2+ del citoplasma al lúmen del retículo sarcoplásmico a expensas de la hidrólisis de ATP, favoreciendo la relajación muscular y recuperando los niveles de Ca2+ de los depósitos intracelulares para la próxima contracción.
En mamíferos la bomba SERCA esta codificada por 3 genes (SERCA 1, 2 y 3) que generan por lo menos 9 isoformas 4.
Un método apropiado para determinar si la actividad enzimática de cualquier isoforma de la bomba SERCA esta alterada, es por medio de la medición directa de su actividad de ATPasa, cuyo ejercicio se muestra a continuación.
Planteamiento experimental
Para determinar la actividad de ATPasa de la bomba SERCA, se utiliza un método espectrofotométrico basado en la formación de un complejo coordinado con el ion fosfato que se libera como producto de la hidrólisis de ATP, generando una coloración azul característica del complejo antimonio-fosfomolibdato conocido como azul de molibdato cuya absorbancia (Abs.) se puede medir en un espectrofotómetro a 850 nm, de forma que la tasa de liberación de fosfato sirve como reportero directo de la actividad enzimática de la bomba 5. Este método permite determinar la actividad de diferentes ATPasas además de la SERCA, mediante la utilización de inhibidores específicos; como en el caso de la ATPasa de Na+-K+ que es inhibida por ouabaína. Para el caso de la bomba SERCA, se utiliza tapsigargina (Tgn), lo que nos permite determinar la actividad específica de esta bomba al realizar la sustracción de los valores de Abs. con Tgn a los valores de Abs. de actividad total (sin inhibidor).
Se utilizan homogenados de músculo esquelético de ratas controles (Ctl) y con síndrome metabólico (SM), a los cuales se les determina la concentración de proteína mediante el método de Lowry. Para el tubo de actividad total de ATPasa, 50 ug de proteína del homogenado se incuban a 37°C durante 5 min con amortiguador que contiene (en mM): KCl 80, MOPS 25, MgCl2 3, Azida de Na+ 5 y EGTA 0.2 (pH 7.0 con TRIS base), también se adiciona Ca2+ 0.2, y el ionóforo de Ca2+ A23187 2 uM. La reacción se inicia con 1 mM de ATP. Para los tubos de actividad inespecífica (es decir, la actividad de ATPasa no dependiente de SERCA) se adicionan los sustratos anteriores además de 1 µM de Tgn.
Después de iniciar la reacción, se toman alícuotas de 100 µL después de 0, 5, 10, 15 y 30 minutos, que se añaden a tubos con 0.9 mL de solución de coloración, la cual contiene (en mM): H2SO4 125, ácido ascórbico 10, tartrato antimónico de potasio (III) 0.4 y heptamolibdato de amonio 0.5. La Abs. a 850 nm se mide 10 min después en un espectrofotómetro.
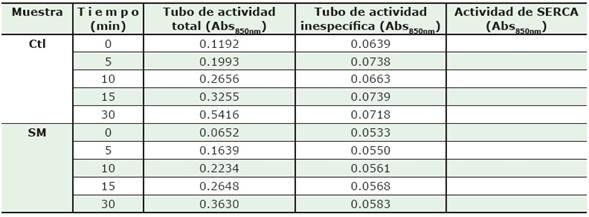
En la Tabla 1 se muestran valores de Abs850nm con y sin Tgn, a diferentes tiempos (0, 5, 10, 15, y 30 min) de homogenados control y SM, respectivamente.
Tabla 1. Absorbancias obtenidas a 850 nm de homogenados de músculo esquelético de ratas control y con síndrome metabólico. La actividad total corresponde a todas las ATPasas en el homogenado, la actividad inespecífica, (en presencia de Tgn) representa la actividad de todas las ATPasa insensibles a este compuesto; y la diferencia (actividad de SERCA) corresponde a la actividad específica atribuida a la bomba SERCA.
Una vez que se tienen las Abs. que corresponden exclusivamente a la actividad de SERCA, estos valores se interpolan en una curva patrón de PO4 -4 (Gráfica 1), para determinar los nmoles de PO4 -4 liberado como se muestra con la Ecuación No. 1.

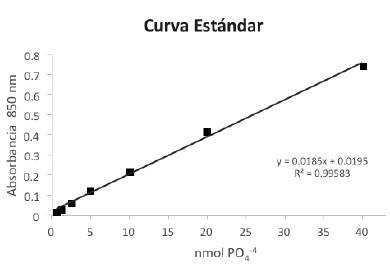
Grafica 1. Curva estándar de de fosfatos. Se presentan los datos de Abs. a 850 nm vs. los nmoles de fosfato. Las mediciones se realizaron por duplicado, por lo tanto los valores de Abs.son el resultado del promedio. Se inserta el resultado de la regresión lineal y el valor del coeficiente de correlación (R2).
Actividades y preguntas
1. Completa los valores de la columna de actividad de SERCA en la Tabla 1, realizando la sustracción correspondiente.
2. Utiliza la Ecuación No. 1 para determinar los nanomoles de PO4 -4 a partir de la absorbancia de SERCA, para cada tiempo y normalizalo con respecto a la cantidad de proteína utilizada.
3. Grafica la actividad de ATPasa de SERCA (nmol de PO4 -4 liberado/mg Pt) con respecto al tiempo (min) para muestras control y con síndrome metabólico. Trata de mantener la misma escala de valores en X y Y para ambas muestras.
4. Determina las velocidades de reacción para cada caso expresadas como nmol de PO4-4/mg de Pt/min.
5. Comparar y discutir los resultados de actividad de ATPasa de muestras control y con síndrome metabólico.

