










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkREB. Revista de educación bioquímica
versión impresa ISSN 1665-1995
Rev. educ. bioquím vol.32 no.1 Ciudad de México mar. 2013
Artículos
Comparación de los diferentes métodos de análisis cinéticos para determinar el tipo de inhibición de dos compuestos
Elizabeth Lira Silva y Ricardo Jasso Chávez
Departamento de Bioquímica, Instituto Nacional de Cardiología, Distrito Federal, México. Correo E: eli_lira_sil@hotmail.com
Recibido: 5 de noviembre de 2012
Aceptado: 12 de febrero de 2013
Introducción
El estudio de los mecanismos cinéticos de enzimas y transportadores así como el efecto de sus inhibidores y activadores sigue siendo una herramienta útil para entender mejor la naturaleza y regulación de las vías metabólicas involucradas en los procesos bioquímicos de todas las células. El mejor entendimiento de los mecanismos catalíticos ha cobrado mayor relevancia biotecnológica ya que nos acerca a un mejor diseño de fármacos en el caso del tratamiento de enfermedades o para realizar modificaciones puntuales y obtener así una mayor producción de metabolitos. De esta forma, para obtener datos que permitan una mejor entendimiento de la participación de enzimas y transportadores en la fisiología de las células, se siguen diseñando compuestos cada vez más específicos que afecten la actividad (Vmax) o afinidad (Km) por sus substratos. Sin embargo, en muchos trabajos de la literatura donde se realizan estudios cinéticos, en ocasiones el análisis de los resultados es sobre-interpretado, incompleto o en el peor de los casos erróneo.
El objetivo de este trabajo es realizar una revisión de los métodos utilizados para analizar el tipo de inhibición de un compuesto. Estos métodos permiten comparar los datos experimentales con patrones predichos para diferentes mecanismos cinéticos, utilizando datos de velocidad inicial (es decir la velocidad de la reacción durante los primeros minutos o segundos, cuando la formación de producto es lineal con respecto al tiempo y la concentración de sustrato es mucho mayor que la concentración de enzima), experimentalmente el método para determinar los patrones de velocidad inicial consiste en variar la concentración de un sustrato frente a concentraciones fijas de otros sustratos y en ausencia de productos.
Existen dos estrategias para analizar el tipo de inhibición de un compuesto, el ajuste por regresión lineal y el ajuste por regresión no lineal.
Ajuste por regresión lineal
Este análisis es ampliamente utilizado en cinética enzimática, los datos se grafican de tal manera que en el eje x se representa la variable independiente y en el eje y la variable dependiente, la línea trazada por regresión lineal se elige para minimizar la suma de los cuadrados de las distancias de los puntos a partir de esa línea. Un método para tratar con relaciones curvas es transformar los datos en una línea recta y después realizar la regresión lineal. Algunos métodos de linearización utilizados son: Lineweaver-Burk (l/v versus l/S), Eadie-Hofstee (v versus v/S), Hanes (S/v versus S) y Scatchard (v/S versus v) donde v es velocidad inicial, S la concentración de sustrato. Estos métodos se pueden aplicar a la ecuación de Michaelis-Menten v=(Vmax [S])/ (Km+[S]), para determinar la Km y Vmax a partir de la pendiente y el intercepto. Entre ellos, el método de Lineweaver-Burk (dobles recíprocos) es el más utilizado como una herramienta gráfica para calcular los parámetros cinéticos de una enzima, cuyo reciproco es 1/v=Km/(Vmax [S])+1/ Vmax, donde v es la velocidad de reacción, Km es la constante de Michaelis-Menten, Vmax es la velocidad máxima, y [S] es la concentración de sustrato. El gráfico de Lineweaver-Burk permite identificar la Km y Vmax; a partir del punto de intersección con el eje de las ordenadas es (l/Vmax), y el de las abscisas al origen se puede obtener (-1/Km). Realizar el ajuste por regresión lineal sobre datos transformados tiene algunas ventajas ya que es muy sencillo y no requiere un software específico, además resulta fácil de evaluar estadísticamente. Sin embargo, estos métodos son muy dependientes de la calidad de los datos de velocidad y se aplican sólo para mediciones de velocidad inicial (4). Los errores en la determinación de velocidad a bajas concentraciones de sustrato se amplifican en el método Lineweaver-Burk y Eadie-Hofstee y en menor media al utilizar Hanes, debido a que estas distorsiones se incrementan en transformaciones que combinan los valores de x y y.
Otra alternativa para determinar los mecanismos cinéticos es realizar ensayos de inhibición utilizando productos, altas concentraciones de sustratos alternos u otras moléculas análogas a cualquiera de estos. Se busca que los inhibidores utilizados sean reversibles, es decir deben formar complejos dinámicos (asociación-disociación) con propiedades catalíticas diferentes a las que tiene la enzima en ausencia del inhibidor. Los inhibidores reversibles pueden ser totales (lineales), es decir al saturar una enzima con un inhibidor total la actividad enzimática se abate por completo y al realizar los regráficos de la ordenada (1/Vmax versus [I]) y la pendiente (Km/Vmax versus [I]) del gráfico de dobles recíprocos se generan líneas rectas o inhibidores parciales (hiperbólicos) cuando se observa cierta actividad enzimática residual en presencia de una concentración saturante de inhibidor y en los regráficos de dobles recíprocos se obtiene una curva hiperbólica (2). Además existe otra clasificación para estos tipos de inhibición con base en el efecto que el inhibidor tiene sobre la eficiencia catalítica (Vmax/Km) y la velocidad máxima. La inhibición competitiva se caracteriza por disminuir la afinidad de la enzima por su sustrato sin afectar la velocidad máxima aparente (en presencia de inhibidor). La inhibición incompetitiva disminuye la Vmax y no afecta la eficiencia catalítica. Y la inhibición no competitiva disminuye el valor de ambos parámetros (Vmax y Vmax/Km).
Para el caso específico de sistemas con inhibidor se puede utilizar el método de Dixon, donde se grafica (1/v versus [I]) y si la cinética se puede describir por la ecuación v=(Vmax [S])/(KS (1+[I]/Ki)+[S](1+[I]/Ki ) ), donde, Vmax es la velocidad máxima, [S] es la concentración del sustrato, KS es la afinidad de la enzima por el sustrato (asumiendo equilibrio rápido, ver referencia 2), [I] es la concentración del inhibidor, Ki es la constante de inhibición (2), el patrón de líneas rectas para diferentes concentraciones de sustrato se intersectan en [I]=-Ki y l/v=[l-(Ki/Ki')/Vmax para determinar la Ki, pero tiene la desventaja de que solo se aplica para mediciones de velocidad inicial y no siempre se puede distinguir entre inhibición mixta e inhibición competitiva (4).
Ajuste por regresión no lineal
Este método permite ajustar los datos experimentales a cualquier ecuación, como en la regresión lineal el procedimiento por regresión no lineal permite determinar los valores de los parámetros que minimizan la suma de los cuadrados de las distancias de los puntos de la curva, con el objetivo de minimizar el residual de la suma de cuadrados. Este método presenta varias ventajas: se pueden determinar simultáneamente varios parámetros cinéticos a diferencia de los métodos lineales, menor restricción en la elección de las variables controladas comparado con los métodos lineales donde los datos se limitan a un intervalo lineal y por lo tanto disminuyen las desviaciones estándar y aumenta la precisión al estimar los parámetros cinéticos, debido a que no se requiere realizar transformaciones lineales a las variables. Sin embargo el ajuste por regresión no lineal no puede resolverse en un solo paso, este debe resolverse de forma iterativa. Realizando iteraciones matemáticas de la ecuación de velocidad mediante software específico. Existen algunos programas comerciales de computadora que realizan regresiones no lineales como Origin, Sigma Plot, Graf fit y Curve fit. Se debe proporcionar una estimación inicial del valor de cada parámetro, después se ajustan estos valores para mejorar el ajuste de la curva a los datos, a continuación se ajustan los valores nuevos para mejorar nuevamente el ajuste y de esta forma estas iteraciones matemáticas continúan hasta que la mejora es mínima. En este trabajo, el análisis no lineal se realizó mediante el "software" Origin 5.0, el cual, da un valor estadístico de Chi cuadrada (Chi2, distribución de probabilidad continua) que ayuda a discernir sobre el mejor ajuste de los datos experimentales a las ecuaciones de los diferentes tipos de inhibición. Se puede aplicar a mediciones de velocidad inicial y a curvas de progreso. Sin embargo, en ocasiones es necesario realizar la estimación de los parámetros cinéticos por métodos lineales antes de realizar las iteraciones, se necesita un "software" especifico y las consideraciones no tomadas en cuenta en el modelo pueden causar ajustes poco satisfactorios, por ejemplo para analizar diferentes tipos de inhibición los métodos lineales son la mejor opción. Además se debe tener cuidado al analizar y realizar un ajuste no lineal, debido a que se pueden producir resultados engañosos si el programa se usa de manera inapropiada.
La selección de un método por regresión lineal o por regresión no lineal para analizar el mecanismo cinético de un compuesto, debe ser cuidadosa y tomar en cuenta tanto sus ventajas y desventajas, el objetivo del análisis; así como sus excepciones para poder predecir de manera precisa el tipo de inhibición esperado, se ha descrito que el tipo de inhibición se puede modificar si se consideran concentraciones saturantes o no saturantes de los sustratos (2).
En este trabajo se hace una revisión de los diferentes métodos de análisis cinético para el estudio del efecto de inhibidores sobre el transportador mitocondrial de citrato publicado en la Revista Molecular Pharmacology titulado Inhibitors of the Mitochondrial Citrate Transport Protein: Validation of the Role of Substrate Binding Residues and Discovery of the First Purely Competitive Inhibitor, con esto se pretende exhortar al lector y en especial al estudiante de posgrado a revisar críticamente la literatura consultada relevante para su proyecto.
Planteamiento del problema
El transportador mitocondrial de citrato (CTP) es crítico en el metabolismo energético y biosintético de células eucariontes, debido a que el citrato es un intermediario esencial en el metabolismo de carbohidratos y lípidos. Es un antiportador que media el intercambio de la forma dibásica de los ácidos dicarboxílicos (malato, succinato) o fosfoenolpiruvato en eucariontes superiores, con los ácidos tricarboxílicos (citrato o isocitrato) en levadura (1).
Los estudios de estructura y función de transportadores de membrana se facilitan en gran medida por la disponibilidad de potentes inhibidores específicos. Sin embargo, para el CTP se conocen pocos inhibidores que además son de bajo potencial. El mejor inhibidor disponible es el 1,2,3- benzenetricarboxylate (BTC), el cual muestra una inhibición de tipo competitivo (1). Con la finalidad de encontrar otros inhibidores más específicos se analizó el 4-chloro-3-[(3-nitrophenyl)sulfamoyl] benzoic acid (compuesto 792949); cuyas fórmulas estructurales se muestra en la figura 1.
Para determinar la naturaleza de inhibición de un CTP-Cys (transportador mitocondrial reconstituido de levadura) por BTC y el compuesto 792949, el ensayo se realizó en proteoliposomas incubados por 10 min a 21°C, midiendo el transporte durante 13-50 segundos (parte lineal) e iniciando la reacción adicionando citrato marcado radioactivamente a concentraciones de 0.05-14 mM, (radioactividad específica de 43 - 430 x 102 cpm/nmol citrato) más concentraciones variables de BTC (0-1.5 mM) o del compuesto 792949 (0-2 mM). La reacción se detuvo por la adición de un exceso de BTC (9 mM). Después del transporte, el citrato intraliposomal marcado radioactivamente fue separado del externo por cromatografía y cuantificado en un contador de centelleo. La velocidad de transporte sensible a BTC se calculó restando el valor control del valor experimental (1). Los datos de velocidad inicial se muestran en las Tablas 1 y 2.



Con estos valores se determinaron mediante diferentes métodos, el tipo de inhibición que ejerce el BTC y el compuesto 792949 sobre el CTP-Cys, los correspondientes parámetros cinéticos (Vmax, KS, Ki), así como las diferentes constantes que afectan la Vmax (factor β) y la afinidad por el substrato (factor α).
Se debe aclarar que la cinética del transporte facilitado puede analizarse mediante las ecuaciones utilizadas para una enzima, debido a que este tipo de transportadores presentan un sitio especifico expuesto en la cara externa de la membrana al cual se enlaza la molécula (y posee un valor KS o Kd) y un valor de Vmax (3).
Inhibición para el compuesto BTC
Para determinar el tipo de inhibición del BTC se debe realizar como primera aproximación un gráfico de Dixon, el cual consiste en graficar el inverso de las velocidades iníciales a diferentes concentraciones de citrato versus la concentración del inhibidor BTC (Fig. 2). Se pueden observar dos comportamientos: a concentraciones bajas se observan hipérbolas lo que sugiere una inhibición parcial, es decir que el complejo transportador-sustrato-inhibidor es catalíticamente activo; mientras que a concentraciones altas se observan líneas rectas lo que sugiere una inhibición simple. Por lo que el análisis de estos dos comportamientos se debe realizar de manera independiente: a) concentraciones de sustrato < 2 mM y b) concentraciones de sustrato > 2 mM.

a) [Citrato ] < 2 mM
Se grafica el inverso de las velocidades iníciales contra el inverso de la concentración de sustrato a diferentes concentraciones de inhibidor (gráfico Lineweaver-Burk) para el BTC (Fig. 3a). En la figura 3a se observa que el BTC modifica tanto el valor de la ordenada al origen (inverso de la velocidad máxima) como las pendientes (KS /Vmax). En este gráfico el punto de intersección de todas las rectas esta en el 2° cuadrante, lo que indica que es una inhibición de tipo mixto y en este caso, mixto parcial (Fig. 4).

Un regráfico m1/S versus [I] o b1/S versus [I] (a partir del gráfico Lineweaver-Burk) ayuda a diferenciar un inhibidor simple de un inhibidor parcial, debido a que con éste último se obtiene una hipérbola (2), lo cual se aplica para el inhibidor BTC (Fig. 5).

El esquema cinético para un inhibidor mixto parcial, se muestra a continuación:

Donde: E es el transportador CTP-Cys, I es el inhibidor, S es el sustrato, P es el producto, KS es la afinidad de la enzima por el sustrato (asumiendo equilibrio rápido), Ki es la constante de inhibición, kp es la constante catalítica, α es el factor que indica cuantas veces se modifica la KS cuando la enzima o transportador está saturada (o) con inhibidor, mientras β expresa el cambio en la constante catalítica en presencia de inhibidor saturante. En este caso, para el inhibidor BTC la intersección en el segundo cuadrante indica que α > 1 y β entre 0 y 1. La ecuación de velocidad para un inhibidor mixto parcial es:

El inverso de la ecuación general 1 es:

La ecuación 2 describe cada recta de la figura 3a (gráfico de Lineweaver-Burk). La Vmax corresponde al inverso de la ordenada de la recta sin inhibidor, que en este caso fue 1/0.00097 = 1031 nmol•min-1•mg proteína-1. La KS es el valor negativo del inverso de la abscisa de la misma recta sin inhibidor, en este caso KS = -1/-6.46 = 0.15 mM.
Debido a que el regráfico de m1/S ó b1/S versus [I] son hiperbólicos (Fig. 5), para calcular los valores de Ki , β, α es necesario obtener Δb (cambio en las ordenadas) y Δm (cambio en las pendientes) de las rectas con inhibidor con respecto a la recta sin inhibidor. Es decir, a cada una de las ordenadas de las rectas con BTC se le resta la ordenada de la recta sin BTC y el mismo procedimiento se aplica a las pendientes y se grafica 1/Δb o 1/m versus 1/ [BTC] (Fig. 6). A partir de la ecuación 2 se obtienen las siguientes ecuaciones:



A partir de las ecuaciones 3 y 4 se obtienen los inversos:

Las ecuaciones 5 y 6 definen las rectas obtenidas al graficar los inversos de los cambios en la ordenada y en la pendiente contra el inverso de la concentración de inhibidor, como se muestra en la figura 6. A partir de los valores obtenidos en estos regráficos, y conociendo los valores de Vmax y KS determinados en la gráfica de dobles recíprocos, es posible determinar el resto de los parámetros cinéticos.
El factor β se despeja de la ordenada obtenida de la recta 1/Δb versus 1/BTC y se obtiene:

Lo que significa que la velocidad con la que el complejo ESI efectúa la catálisis disminuye un 50 % respecto al complejo ES.
El factor α se puede calcular utilizando el valor de la ordenada de la recta 1/Δm versus 1/BTC, la cual despejando y sustituyendo se obtiene lo siguiente:

Lo que significa que la afinidad del transportador por el sustrato (KS) disminuye aproximadamente 6.5 veces cuando el transportador esta interactuando con el inhibidor (EI) para formar el complejo (ESI). Finalmente se obtiene el valor de la Ki a partir de la definición de la abscisa para cualquiera de las dos rectas utilizando los valores ya determinados de α y β, recordando que debe ser igual en ambos casos.

Con base en el análisis realizado se puede decir que el BTC a concentraciones de citrato < 2 mM, es un inhibidor de tipo mixto parcial cuya unión al complejo transportador -sustrato (ES) aumentó aprox. 6.5 veces la Ki, es decir disminuye la afinidad con la que el inhibidor se une al complejo (ES) para formar el complejo productivo (ESI) cuya kp se alteró y disminuyó un 50% (β = 0.51).
b) [Citrato ] > 2 mM
En la figura 3b se grafica el inverso de las velocidades iníciales contra el inverso de la concentración de sustrato a diferentes concentraciones de inhibidor (gráfico Lineweaver-Burk). Se observa que el BTC modifica las pendientes (KS /Vmax) conforme aumenta la concentración de inhibidor, mientras que el valor de la ordenada al origen (inverso de la velocidad máxima) permanece constante. También se observa que conforme se incrementa la concentración de inhibidor, el intercepto sobre el eje 1/S se acerca al origen, esto es la Km aumenta. Este patrón sugiere que el BTC a concentraciones de citrato > 2 mM presenta una inhibición de tipo competitivo simple. El regráfico de las pendientes del gráfico de Dixon (Fig. 7), el cual parte del origen, confirmó este tipo de inhibición.

El esquema cinético para un inhibidor competitivo simple es:

Donde: E es el transportador CTP-Cys, I es el inhibidor, S es el sustrato, P es el producto, KS es la afinidad de la enzima por el sustrato (asumiendo equilibrio rápido), Ki es la constante de inhibición y kp es la constante catalítica.
La ecuación de velocidad para un inhibidor competitivo es:

El inverso de la ecuación general es:

En el gráfico de dobles recíprocos (Fig. 3b), se obtuvo una Vmax = 1369 nmol•min-1•mg proteína-1, KS = 0.72 mM para el BTC a [citrato] > 2 mM, sin embargo para obtener la Ki es necesario realizar el regráfico de m1/S o Km versus [I]. A partir de la ecuación 11, se obtienen las siguientes ecuaciones:


El regráfico m1/S versus [I] o b1/S versus [I] (obtenido a partir del gráfico Lineweaver-Burk) fue lineal, lo que confirma que el BTC a altas concentraciones de citrato actúa como un inhibidor competitivo simple (Fig. 8a y 8b). Considerando las ecuaciones 13 y 14, así como los regráficos se obtuvo Ki = 0.11- 0.15 mM.
Como se menciona al inicio el gráfico de Dixon ayuda a identificar el tipo de inhibición según el patrón de rectas obtenido, pero también es posible obtener la Ki.
La ecuación para el gráfico de Dixon (Fig. 2) para un inhibidor competitivo es:

A partir de esta gráfico, se puede determinar el tipo de inhibición, si se sabe que la inhibición es competitiva y se conoce la Vmax, se puede trazar una línea a la altura de 1/Vmax y el valor de -[I] = Ki en la intersección de esas dos líneas, las ecuaciones que describen este comportamiento son:

Le pendiente de la grafica de Dixon es:

También se realiza el análisis por regresión no lineal utilizando las ecuaciones generales para un inhibidor de tipo mixto parcial y para un inhibidor competitivo, este método es una forma confiable para obtener los valores de KS, Ki y Vmax; a partir del análisis global de datos, sin embargo en este caso el mejor ajuste no lineal se realizó utilizando la ecuación para un inhibidor competitivo simple, considerando que el valor de Chi2 fue menor (Fig. 9).

Inhibición para el compuesto 792949
A continuación se analiza el tipo de inhibición para el compuesto 792949 sobre el transporte de citrato (Fig. 10). Se observa que el gráfico de Dixon para el compuesto 792949, muestra un comportamiento lineal con pendiente positiva, lo que sugiere que se trata de un inhibidor simple.
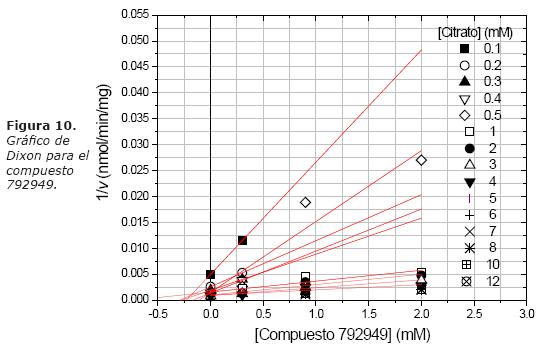
Sin embargo, para conocer el tipo de inhibición se puede usar el inverso de las velocidades iníciales contra el inverso de la concentración de sustrato (gráfico Lineweaver-Burk) a diferentes concentraciones del compuesto 792949 (Fig. 11). Todas las rectas a diferentes concentraciones de inhibidor junto con la curva control se intersectaron en un punto sobre el eje 1/v, el intercepto 1/Vmax. Además, conforme la concentración de inhibidor se incrementa el intercepto sobre el eje 1/S se acerca al origen, esto es la Km aumenta; este patrón sugirió que es una inhibición competitiva simple y que el compuesto 792949 es un inhibidor de tipo competitivo.

En el gráfico de dobles recíprocos (Fig. 11), se obtuvo una Vmax= 1545 nmol•min-1•mg proteína-1, KS = 0.66 mM para el compuesto 792949; sin embargo, para obtener la Ki es necesario realizar el regráfico de m1/S o Km versus [I]. A partir de la figura 12 y considerando las ecuaciones 11, 13 y 14, se obtuvo que la Ki = 0.064-0.080 mM.
Con base en el patrón de líneas rectas obtenidas a partir del gráfico de Dixon y con el regráfico de las pendientes, el cual parte del origen se sugiere que el compuesto 792949 es un inhibidor de tipo competitivo simple. Y utilizando las ecuaciones 15 y 16, se obtuvo la Ki = 0.080 mM.
También se realizó el ajuste no lineal para el inhibidor competitivo 792949 (Fig. 13) y se obtuvieron valores de Vmax, KS y Ki muy similares a los obtenidos por los métodos lineales descritos.

Se puede decir que el compuesto 792949 es un inhibidor de tipo competitivo simple cuya unión al transportador no permite la unión del sustrato debido a que son mutuamente excluyentes (Tabla 1). Es posible que este nuevo compuesto pueda ser utilizado como otra alternativa para inhibir el CTP-Cys además del inhibidor clásico BTC, debido a que su KS y Ki son similares. Sin embargo estos parámetros cinéticos fueron determinados en el CTP-Cys reconstituido en liposomas, es decir in vitro (, por lo que antes de utilizar estos compuestos in vivo se deben evaluar la permeabilidad y la toxicidad de este tipo de compuestos.
Análisis
En esta revisión se determinó el mecanismo cinético de inhibición del transportador CTP-Cys por BTC y por el compuesto 792949. Con base en el análisis realizado se puede decir que el BTC a concentraciones de citrato < 2 mM, es un inhibidor de tipo mixto parcial, mientras que el BTC a altas concentraciones de citrato actúa como un inhibidor competitivo simple. La diferencia en mecanismo de inhibición encontrado a bajas y a altas concentraciones de BTC, así como el compuesto 792949 fue un inhibidor competitivo simple habla de la importancia fisiológica que puede tener el transportador CTP en la regulación del metabolismo celular, específicamente en la reducción del transporte de un metabolito ya que permite la regulación de vías metabólicas, evitando la utilización de sustratos y formación de subproductos. Además la búsqueda y el análisis de los compuestos que inhiben una enzima o transportador y los que no, pueden proporcionar información sobre las propiedades de unión del transportador. Estos inhibidores también pueden proporcionar información sobre la especificidad de sustratos que podría ser útil en el estudio de vías metabólicas completas. Para esta evaluación se utilizaron tanto métodos por regresión lineal como el ajuste por regresión no lineal, debido a que el uso de uno solo de ellos puede arrojar un análisis poco confiable. De los métodos utilizados, el gráfico de Dixon permite predecir el tipo de inhibición, pero no es suficiente para distinguir entre inhibición mixta y competitiva, mientras que a partir de la linearización de la ecuación para un inhibidor de tipo competitivo mediante el método de Lineweaver-Burk fue posible obtener los parámetros cinéticos como Vmax, Km. Sin embargo, estas manipulaciones matemáticas pueden resultar poco confiables debido a la distribución de los puntos experimentales a lo largo de las rectas como consecuencia de un ajuste poco satisfactorio debido a los rearreglos realizados en la ecuación y del error presente en los datos originales (2). Por lo que para determinar con mayor precisión el mecanismo de inhibición mixto y competitivo para el BTC, así como la inhibición competitiva para el compuesto 792949 se realizaron los regráficos a partir de los gráficos de dobles recíprocos. Los regráficos lineales son útiles porque también permiten determinar el valor real de las constantes cinéticas como Ki, Vmax/Km y Vmax, entre otras (2). Los datos aquí presentados también se analizaron por ajuste no lineal mediante el software Origin 5.0 a la ecuación que describe un mecanismo de inhibición mixto parcial y competitivo para el BTC con la finalidad de poder comparar entre estos mecanismos. Debido a que la Chi2 es más pequeña en el ajuste para la ecuación de inhibición competitiva y a que los errores experimentales y los valores de las constantes cinéticas determinadas a partir del ajuste (Vmax, Km y Ki) son mucho más reales que los obtenidos para el mecanismo mixto parcial. Se consideró como mejor ajuste el que corresponde a la ecuación del mecanismo de inhibición competitiva. Mientras que el ajuste para un inhibidor competitivo por regresión no lineal para el compuesto 792949, proporcionó parámetros cinéticos similares a los obtenidos por los métodos lineales.
Conclusión
Esta revisión y el análisis realizado confirma que estos métodos no son excluyentes, por lo que la mejor forma para realizar el análisis cinético de una enzima y en este caso específico del transportador CTP-Cys fue utilizar de manera complementaria los métodos por regresión lineal y el ajuste no lineal; sin embargo, es necesario utilizar otras estrategias experimentales para confirmar el mecanismo cinético y el tipo de inhibición de los compuestos analizados.
Referencias
1. Aluvila S, Sun J, Harrison DH, Walters DE, Kaplan RS.(2010) Inhibitors of the mitochondrial citrate transport protein: validation of the role of substrate binding residues and discovery of the first purely competitive inhibitor. Mol Pharmacol 77 (1): 26-34. [ Links ]
2. Segel I H (1975). Enzyme Kinetics, John Wiley and Sons, New York. pp. 957. [ Links ]
3. Keith Wilson and John Walker Principles & Techniques of Practical Biochemistry. 5th edition. Cambridge University Press 2000. [ Links ]
4. Marcus M. y B. HESS. 1976. The analysis of kinetic data in biochemistry a critical evaluation of methods. FEBS Lett. 63 (2): 225-230. [ Links ]

