











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El chile habanero (Capsicum chinense Jacq.) es una planta anual de tipo arbustivo (Lim, 2013) nativa de las tierras bajas de la cuenca amazónica, donde se dispersó en la época prehispánica a la cuenca del Orinoco y otras regiones de Perú, las Antillas y finalmente arribó a la Península de Yucatán (Trujillo, 2018). Actualmente este es el cultivo hortícola de mayor importancia económica para la península de Yucatán en México (Ruiz-Lau et al.., 2011); desde 2010 posee la denominación de origen en esta región (CICY, 2016). El fruto posee diversas propiedades nutraceúticas y medicinales (Lim, 2013), es usado también en la industria alimentaria, farmacéutica y militar (Ruiz-Lau et al.., 2011).
Debido a esto, ha surgido la necesidad de realizar investigación de su cultivo en ambiente controlado con iluminación LED para analizar el comportamiento como una alternativa de producción, ya que con este sistema se elimina la aparición de plagas y enfermedades, por lo tanto, la obtención de un producto orgánico libre de plaguicidas e inocuo a la salud humana. Otra ventaja sería su producción continua, sin estacionalidad de cultivo, por el control de las variables ambientales y de la fuente de luz, lo cual en su conjunto trae un incremento exponencial de la producción (Kozai y Niu, 2016).
Actualmente no existe investigación de esta especie bajo estas condiciones de crecimiento; sin embargo, se han realizado investigaciones por parte de Gangadhar et al.. (2012) en chile picante cv. Cheonyang y Naznin et al.. (2019) en pimiento dulce enano cv. Redstart, en ambas investigaciones existieron diferencias entre los tratamientos de luz LED azul y roja que evaluaron, respecto a las variables del crecimiento como altura de planta, área foliar, peso fresco de la parte aérea, número de frutos por planta y en los pigmentos fotosintéticos en hoja como clorofila a, b, total a/b y carotenoides.
Con base en la información anterior, el objetivo del presente estudio fue evaluar el efecto de diferentes proporciones de luz LED roja y azul sobre crecimiento, concentración de pigmentos fotosintéticos, fenoles totales y nutrimental en hoja; así como determinar si los tratamientos influyen en la germinación de polen, grosor de hoja, frecuencia estomática, anatomía de ovario y antera de chile habanero.
Materiales y métodos
Cámara y gabinetes de crecimiento
Se realizó una adaptación de una cámara de crecimiento a partir de un cuarto frío (TORREY®, MEX), con medidas interiores de 2.76 m x 2.76 m x 1.90 m (ancho, largo y altura), las modificaciones internas de la cámara de crecimiento se basaron en el diseño de Katagiri et al.. (2015), se reguló la temperatura ambiental y humedad relativa con un equipo de aire acondicionado portátil (LG® modelo LP1017WSR 10200 BTU, KOR) y un deshumidificador (Whirlpool® modelo WAD200 5Q 20L/24 h, USA). En el interior de esta cámara, se diseñaron seis gabinetes de crecimiento a partir de un anaquel metálico de 0.3 m x 0.85 m x 1.85 m (ancho, largo y altura) a los cuales se les añadieron paredes de placas de unicel de 2 cm de grosor forradas con papel aluminio para tener máxima reflexión de la luz en el interior y para aislar los tratamientos uno de otro.
Tratamientos con diferentes proporciones de luz LED azul y roja
Las luminarias que se utilizaron fueron hechas a mano, compuestas por perfiles de aluminio de 5 cm x 85 cm (ancho x largo) en donde se anclaron focos LED azul y rojos (Longniu® modelo GR-DB015 de 4 watts, CHN) y las lámparas fluorescentes (Osram® modelo Octron/F017 de 17 watts, DEU). Las luminarias fueron colocadas debajo de la parte superior de cada uno de los gabinetes de crecimiento. Los tratamientos consistieron en un testigo fluorescente (4 lámparas) y en los tratamientos A0 %:R100 % (A0) con 24 focos LED rojos, A34.5 %:R65.5 % (A34.5) con 18 focos LED rojos y 6 azules, A57.1 %:R42.9 % (A57.1) con 12 focos LED rojos y 12 azules, A76 %:R24 % (A76) con 6 focos LED rojos y 18 azules, A100 %:R0 % (A100) con 24 focos LED azules. Se utilizaron dos perfiles por tratamiento (12 focos LED por perfil) y el testigo con cuatro perfiles (1 lámpara fluorescente por perfil). La densidad de flujo de fotones fotosintéticos (DFFF) (µmol m2 s-1) y las propiedades físicas del espectro electromagnético de los tratamientos se midió con un espectroradiómetro (Apogee instruments® modelo SS-110, EUA) donde se tomaron 330 mediciones sobre el área de cultivo (66 cm x 20 cm= 1320 cm2), que fue la superficie en relación a la parte inferior del gabinete, donde las plantas tuvieron su desarrollo durante el transcurso del experimento, y así verificar que los tratamientos tuvieran la misma densidad de flujo de fotones fotosintéticos (DFFF). Se realizó un análisis estadístico de las variables físicas de los tratamientos (Tabla 1) para asegurar que las intensidades de luz entre tratamientos LED fueran iguales y las proporciones de luz azul y roja fueran diferentes; además de verificar el pico de longitud de onda (nm) e intervalo de la luz LED presente en los tratamientos. Se estableció un fotoperiodo para todos los tratamientos de luz de 15 h por día durante 145 días después del trasplante.
Tabla 1 Propiedades físicas de los tratamientos con diferentes proporciones de luz LED azul y
roja en chile habanero cultivado bajo condiciones controladas.
Table 1. Physical properties of treatments with
different blue and red LED light ratios on habanero pepper grown
under controlled conditions.
| Tratamientosx | ||||||
| FL | A0 | A34.5 | A57.1 | A76.0 | A100 | |
| PrA | Testigo | A0 % | A34.5 % | A57.1 % | A76 % | A100 % |
| PrR | Testigo | R100 % | R65.5 % | R42.9 % | R25.8 % | R0 % |
| Dtot | 119.1±0.6b | 241.5±3.1a | 241.4±3.3a | 244.4±2.4a | 241.4±1.8a | 243.8±2.5a |
| LID | 6.4±0.0b | 13.0±0.2a | 13.0±0.2a | 13.2±0.1a | 13.0±0.1a | 13.2±0.1a |
| λA | - | - | 457±0b | 457±0a | 457±0b | 457±0b |
| %DA | - | - | 34.5±0.6d | 57.1±0.3c | 76.0±0.3b | 100.0±0.0a |
| λR | - | 636±0b | 635±0c | 637±0a | 636±0b | - |
| %DR | - | 100.0±0.0a | 65.5±0.6b | 42.9±0.3c | 24.0±0.3d | - |
xMedias con letras iguales en las filas son estadísticamente iguales (Tukey, P ≤ 0.05). FL= fluorescente con un intervalo de 416nm - 715nm (testigo); A= Luz LED azul; R= Luz LED roja; PrA= Proporción luz azul; PrR= Proporción luz roja; Dtot= Densidad de flujo de fotones fotosintéticos total de tratamiento (µmol m-2 s-1); LID= Luz integral diaria (mol m-2 d-1); λA= Pico de longitud de onda (nm) color azul; %DA= Porcentaje de densidad de flujo de fotones fotosintéticos de luz azul (%); λR = Pico de longitud de onda (nm) color rojo; %DR= Porcentaje de densidad de flujo de fotones fotosintéticos de luz roja (%).
Material vegetal
Se utilizaron semillas de chile habanero ‘Mayan Ba’alché’ del CICY (Centro de Investigación Científica de Yucatán), las cuales previamente se sembraron en un semillero de 200 cavidades, se colocó una semilla por cavidad a una profundidad de 1 cm en una mezcla de turba rubia (Pro mix®, CAN) y perlita (Agrolita®, MXN) en proporción 3:1 v/v, los semilleros se cortaron para tener ٨٠ cavidades (4 filas x 20 columnas) y tener una charola por tratamiento. Una vez sembradas, se colocaron en un laboratorio donde tuvieron luz solar incidente a través de una ventana durante la duración del día y fueron regadas con agua potable (pH 7.1 y CE 0.42 dS m-1). La temperatura ambiental (día 25.8±0.2 oC/noche 22.6±0.1 oC) y la humedad relativa (día 51.5±0.6 %/noche 58.6±0.6 %); se registraron con un datalogger (Extech instruments® modelo RH10, USA). A los 17 días en estas condiciones, al inicio del desarrollo, las plántulas contaban con dos cotiledones y dos hojas; en este momento las charolas con las plántulas se trasladaron a la cámara de crecimiento, para iniciar los tratamientos con luz. En estas nuevas condiciones las plántulas crecieron por 35 días a una temperatura ambiental (oC) promedio (día 28.2±0.1/noche 25.3±0.1) y humedad relativa (%) promedio (día 56.2±0.5/noche 77.1±0.5). El riego se hizo diariamente a cada una de las charolas con 300 mL de la solución de Hoagland modificada a 1/5 de su concentración (Bojórquez-Quintal et al.., 2014) con pH de 6.8 y CE de 1.35 dS cm-1, donde se abastecieron los macronutrientes de la forma siguiente: 1.2 mM KNO3, 0.8 mM Ca(NO3)2, 0.2 mM KH2PO4, 0.2 mM MgSO4 y los micronutrientes se abastecieron con la mezcla Tradecorp® AZ (0.065g L-1).
Trasplante
Una vez que transcurrieron 35 días, se obtuvieron plántulas adecuadas para trasplante, se eligieron tres por tratamiento y se trasplantó una por maceta, las cuales conservaron los mismos tratamientos de luz (Tabla 1) una vez trasplantadas, se mantuvo así por 145 días después del trasplante (ddt). Se ocuparon tres macetas de 5 L por tratamiento con una mezcla de turba:perlita (3:1), turba rubia (Pro mix®, CAN) y perlita (Agrolita®, MXN) y se regaron diariamente con 1 L de la solución nutritiva de Hoagland (Hoagland y Arnon, 1950) a 100 % de concentración de macronutrientes con un pH de 5.8 y CE de 1.33 dS cm-1, los micronutrientes se abastecieron con la mezcla Tradecorp® AZ (0.065g L-1). La temperatura ambiental promedio fue (día 26.7±0.1 oC/noche 23.9±0.1 oC) y la humedad relativa promedio (día 66.1±0.2 %/noche 82.3±0.2 %), éstas se registraron cada minuto con el promedio de tres datalogger (Extech instruments® modelo RH10, USA), los cuales se colocaron uno por gabinete a nivel de copa de la planta en los tratamientos A76 %:R24 %, A34.5 %:R65.5 % y el testigo. La temperatura promedio de la solución nutritiva se midió con un termómetro infrarrojo (Extech instruments® modelo 42530, USA) y tuvo un valor de 26.2±1.3 oC. La concentración promedio de CO2 se midió con un medidor portátil de CO2 ambiental (Extech instruments® modelo CO250, USA) y tuvo una media de 404±52 ppm. Durante el crecimiento de la planta se realizaron podas de chupones y ramas para tener una planta con dos tallos, además se realizaron podas de hojas en la parte inferior de la planta con el fin de evitar un sombreo excesivo entre éstas. También se realizaron polinizaciones manuales con un pincel para aumentar el porcentaje de amarre de frutos.
Variables de crecimiento de la planta
A los 145 (ddt), las plantas se cosecharon y se determinó la altura de planta (cm), el peso fresco de tallo y de hoja (g planta-1), con una balanza de precisión (Ohaus® Scout Pro, USA), el peso fresco de la parte aérea (g planta-1) se estimó con la suma del peso fresco del tallo y de las hojas. El diámetro de tallo (mm) se midió con un vernier digital (Truper®, MEX); el área foliar (cm2 planta-1) se determinó con un integrador de área foliar LI-3100 (LICOR®, USA) y se calculó el índice de área foliar con base en la fórmula [área foliar (cm2)/área de cultivo (cm2)]. La parte aérea de la planta se secó en una estufa de aire forzado (BINDER® modelo FED 115, DEU) por 48 horas a 65 oC y se registró el peso seco de tallo y hoja (g planta-1), ambos por separado, con una balanza de precisión (Ohaus® Golden series, USA), el peso seco de la parte aérea (g planta-1) se determinó mediante la suma del peso seco del tallo y de las hojas. Los frutos se cosecharon a los 109 y 145 ddt y se contabilizó el total por tratamiento. La frecuencia estomática (número de estomas por mm2) se determinó mediante la aplicación de barniz transparente sobre el envés de la hoja; posteriormente, se despegó la calca que se formó en la hoja y se colocó en un portaobjetos para la contabilización de los estomas con un microscopio (Carl Zeiss® Axiostar, DEU), con aumento de 40 x. En cada una de las variables medidas se utilizaron tres repeticiones por tratamiento, excepto para la frecuencia estomática donde se tuvieron 10 repeticiones.
Concentración de pigmentos fotosintéticos en hoja
Las concentraciones de clorofila a, b y total (mg g-1 de peso fresco), proporción de clorofila a/b y concentración de carotenoides (mg g-1 de peso fresco) se determinaron mediante la metodología descrita por la AOAC (1980). Estas variables se determinaron en hojas fuente (de reciente maduración), completamente desarrolladas, recién cortadas y utilizando tres hojas por planta por tratamiento.
Compuestos fenólicos totales en hoja
Se tomaron 2 g de hojas fuente completamente desarrolladas recién cortadas, las cuales se cortaron finamente y se les adicionó 10 ml de MeOH al 80 % (v/v), posteriormente se homogeneizó mediante sonicación por 15 minutos a temperatura ambiente y se dejó reposar durante 24 h en refrigeración y oscuridad. Pasado este tiempo, se tomó 0.1 mL del extracto metanólico previamente preparado, se agregó 0.1 mL del reactivo Folin-Ciocalteu 0.2 N, 4.5 ml agua destilada y 0.3 ml de Na2CO3 2 % (p/v), esta mezcla se incubó a temperatura ambiente y en oscuridad por 2 h. Posteriormente se tomaron lecturas de absorbancia del sobrenadante en un espectrofotómetro Genesys 10s (Thermo Fisher Scientific, USA) a una longitud de onda de 765 nm. La concentración de compuestos fenólicos se calculó a partir de una curva estándar preparada con ácido gálico y se expresó en mg equivalentes de ácido gálico por 100 g de peso fresco (mg EAG 100 g-1 PF) de acuerdo con el método de Singleton y Rossi (1965) con algunas modificaciones. Se utilizaron tres repeticiones de cada tratamiento.
Concentración nutrimental en hojas
Se determinaron P, K, Ca, Mg, Fe, Mn, Zn, B y Cu en un espectrofotómetro de Emisión Atómica de Plasma por Inducción Acoplada (ICP-AES) (VARIAN modelo Liberty II, USA); la concentración de N se determinó con el método micro-Kjeldahl (Alcántar y Sandoval, 1999). Se utilizaron para las determinaciones 0.1 g de hojas fuente completamente desarrolladas previamente secas en una estufa de aire forzado (BINDER® modelo FED 115, DEU) por 48 horas a 65 oC. Para la determinación de los elementos se utilizaron tres repeticiones por tratamiento.
Variables anatómicas y germinación de polen
A los 106 ddt, se realizaron cortes anatómicos longitudinales de flores en antesis y cortes transversales de hojas completamente desarrolladas. Por tratamiento se utilizaron cinco flores y cinco hojas. En el caso de las flores, se eliminaron los pétalos y posteriormente se seccionó el ovario a la mitad de manera longitudinal con un bisturí. Para el caso de las hojas se tomó una sección de 5 x 5 mm por hoja, con ayuda de un bisturí, de la parte media de la hoja que se ubicó entre la vena central y el borde. Tanto para el caso de los ovarios como de hojas se fijaron en FAA (50 % etanol al 96 % + 5 % ácido acético glacial + 10 % formaldehído al 37 % + 35 % de agua). Posteriormente se realizó un lavado en agua y etanol al 70 % y a continuación se conservaron en GAA (25 % glicerol + 50 % 2-propanol + 0.002 % eosina + 25 % agua) hasta el momento de incluirlos en parafina para realizar los cortes anatómicos que tuvieron un grosor de 10 µm, para lo cual se utilizó un micrótomo rotatorio. La observación se realizó en un microscopio (Carl Zeiss® Axiostar, DEU) con aumentos de 10X y 40X para ovario más anteras y de 40X para hoja. El grosor de la hoja se midió usando micrómetros objeto y de ocular. Se evaluó la anatomía de los óvulos en el ovario en base a la presencia o ausencia de óvulos, pared del ovario, núcleo del óvulo y tegumentos. Por otra parte, en la anatomía de polen en anteras, se determinó la viabilidad del polen en base al grado de coloración por parte de la tinción sobre el citoplasma de éste, en donde una tinción completa y de color rojo intenso es indicativo de un polen viable, por otra parte, un citoplasma no coloreado, o de una tonalidad baja es indicativo de un polen no viable.
Para la viabilidad de polen se usó la técnica modificada de Mercado et al.. (1994) , se preparó un medio sólido con 6 g L-1 de agar-agar, 10% de sacarosa, 0.1 mM de ácido bórico y 1 mM de cloruro de calcio para dos cajas Petri. Se recolectaron cinco flores frescas en estado inicial de apertura a antesis a los 135 ddt, excepto el tratamiento A100 %:R0 % que ya no contaba con flores, el polen fresco de las anteras se depositó sobre la superficie del medio de agar. Las cajas Petri se colocaron dentro de sus respectivos tratamientos de luz por dos horas, se observaron en un microscopio (Carl Zeiss® Axiostar, DEU) con un aumento de 40X y se calculó el porcentaje de germinación de polen en cuatro campos por caja Petri.
Diseño experimental y análisis estadístico
El experimento se estableció en un diseño completamente al azar, una repetición estuvo constituida por una plántula. Los datos se sometieron a un análisis de varianza (ANOVA), a la prueba de comparación de medias de Tukey (P < 0.05) utilizando el programa SAS V9.0 (SAS Institute, 2002).
Resultados y discusión
Variables de crecimiento de la planta
Los resultados de las variables de crecimiento se muestran en la Tabla 2. Se observó que las plantas que recibieron tratamientos con luz LED la altura de planta no fue significativamente diferente entre estos, sin embargo, el tratamiento con 100% de luz azul fue mayor al testigo. Las plantas del tratamiento A100, en donde no se recibió ninguna cantidad de luz roja, presentaron la menor frecuencia estomática respecto a las plantas que si recibieron cierto porcentaje de luz roja. Bajo la presencia de luz LED roja en los tratamientos evaluados, se obtuvieron frutos, excepto en las plantas que recibieron 34.5 % de luz azul, mientras que en las plantas del tratamiento monocromático rojo se obtuvo la mayor cantidad de frutos. Las variables que resultaron estadísticamente no significativos pudieron deberse a las podas de hojas a las cuales fueron sometidas las plantas durante el ciclo de crecimiento.
Tabla 2 Variables de crecimiento evaluadas en chile habanero cultivado bajo condiciones
controladas con diferentes proporciones de luz LED azul y roja.
Table 2. Growth variables evaluated in habanero pepper
grown under controlled conditions with different ratios of blue and
red LED light.
| Var | Tratamientosx | |||||
| FL | A0 | A34.5 | A57.1 | A76.0 | A100 | |
| AP | 85.50±10.28b | 102.90±35.52b | 94.63±11.66b | 97.37±17.41b | 109.70±17.97b | 176.77±4.57a |
| GT | 8.03±0.28ns | 10.91±1.71ns | 11.30±1.40ns | 11.95±0.73ns | 9.05±0.85ns | 9.80±0.33ns |
| AF | 2008.0±714.2ns | 1991.0±1592.3ns | 2703.0±1037.3ns | 2661.0±828.0ns | 2222.0±1220.5n | 1989.0±565.1ns |
| IAF | 4.56±1.62ns | 4.56±3.62ns | 6.16±2.36ns | 6.06±1.88ns | 5.03±2.77ns | 4.53±1.28ns |
| PFT | 47.00±12.45ns | 74.73±48.50ns | 81.27±26.48ns | 61.03±9.80ns | 80.00±48.66ns | 182.63±45.76ns |
| PFH | 34.27±4.64ns | 36.50±11.11ns | 49.90±7.47ns | 56.23±7.41ns | 44.83±10.10ns | 44.97±6.74ns |
| PFPA | 81.27±23.82ns | 111.23±76.79ns | 131.17±45.77ns | 117.27±14.82ns | 124.83±74.08ns | 227.60±62.48ns |
| PST | 6.49±2.36ns | 13.68±9.50ns | 12.52±5.96ns | 12.87±3.95ns | 12.41±8.82ns | 22.43±6.67ns |
| PSH | 3.31±1.55ns | 4.17±3.66ns | 5.04±2.58ns | 5.78±2.30ns | 5.62±3.76ns | 5.65±2.51ns |
| PSPA | 9.80±1.51ns | 17.85±5.10ns | 17.55±3.31ns | 18.65±2.38ns | 18.03±4.86ns | 28.07±3.52ns |
| FE | 127.00±5.12b | 144.70±7.32ab | 128.30±6.30ab | 154.00±11.57ab | 159.30±7.14a | 88.30±6.76c |
| NF | 0 | 24 | 0 | 5 | 4 | 0 |
xMedias con igual letra en las filas son estadísticamente iguales (Tukey, P ≤ 0.05). ns= No significativo; Var= Variables; FL= fluorescente (testigo); AP= Altura de planta (cm); GT= Grosor de tallo (mm); AF= Área foliar (cm2 planta-1); IAF= Índice de área foliar; PFT= Peso fresco de tallo (g planta-1); PFH= Peso fresco de hoja (g planta-1); PFPA= Peso fresco de la parte aérea (g planta-1); PST= Peso seco de tallo (g planta-1); PSH= Peso seco de hoja (g planta-1); PSPA= Peso seco de la parte aérea (g planta-1); FE= Frecuencia estomática (mm2); NF= Número de frutos por tratamiento.
Verma et al.. (2018) encontraron la mayor altura de planta en cultivo de digitales con un proporción de luz LED azul y roja A100 %:R0 % respecto a un tratamiento testigo fluorescente. Por otra parte, Naznin et al.. (2019) no encontraron diferencias estadísticas en altura de planta de espinaca y albahaca entre cuatro diferentes tratamientos de luz roja y azul (A17 %:R83 %, A9 %:R91 %, A5 %:R95 %, A0 %:R100 %). Las plantas del testigo, a pesar de que recibieron la mitad de la intensidad luminosa en comparación con los tratamientos LED (Tabla 1), no tuvieron diferencia en altura de planta con los tratamientos LED que tuvieron presencia de luz roja, sin embargo, con las plantas que recibieron 100 % de luz azul si fue menor.
La luz roja a través de la acción de los fitocromos, tiende a elongar la altura de las plantas, mientras que la luz azul se le ha asociado con la formación más compacta y de menor altura en plantas (Berkovich et al.., 2017; Pocock, 2015). Sin embargo, en habanero, al no existir diferencias significativas entre tratamientos LED, la luz roja o azul no tuvieron un efecto evidente sobre la altura de planta, salvo en el caso de las plantas del testigo en comparación con las que recibieron 100 % de luz azul, la diferencia de altura pudo deberse principalmente a la menor intensidad luminosa que presentó.
Wollanger y Runkle (2015) encontraron un comportamiento similar en el área foliar en plántula de tomate. El índice de área foliar debe encontrarse entre 3.0 a 4.0 para que se aproveche de un 90 a 100 % de luz incidente, si es mayor el valor como en los resultados obtenidos, supone una pérdida de fotosintatos asimilados debido a la existencia de una mayor respiración por causa del sombreo en hojas inferiores de la planta Yamori (2016) .
Hogewoning et al.. (2010) encontraron en pepino resultados similares en la frecuencia estomática, con tratamientos de 0 %, 15 %, 30 %, 50 % de luz LED azul respecto a la luz LED roja donde no tuvieron diferencias estadísticas entre éstos. De manera contraria Wang et al.. (2016) encontraron que el tratamiento monocromático rojo obtuvo la menor frecuencia estomática en lechuga en comparación con diferentes proporciones de luz LED azul y roja. El desarrollo de los estomas se ha asociado a los fitocromos, los cuales son activados por luz roja (Demotes-Mainard et al.., 2016; Pocock, 2015), con lo cual, se confirma que en plantas de habanero que recibieron diferentes proporciones de luz LED roja junto con el testigo, que posee un espectro completo de luz (416-715 nm), existió una mayor frecuencia estomática que las plantas que recibieron 100 % de luz azul.
Gangadhar et al.. (2012) encontraron en chile picante cv. Cheonyang el mayor número de frutos por planta con una proporción de luz azul y roja de A50 %:R50 % (3.8), seguido del testigo fluorescente (3.3) y el tratamiento A0 %:R100 % (3.0), el menor número se obtuvo con el tratamiento A100 %:R0 % (2.1). En el caso de Naznin et al.. (2019) registraron en pimiento dulce enano cv. Redstart, el mayor número de frutos por planta con una proporción de luz azul y roja de A5 %:R95 % (5), A9 %:R91 % (5) y los menores con los tratamientos A17 %:R83 % (1) y A0 %:R100 % (1).
El rendimiento de frutos del testigo pudo haber sido afectado debido a que tuvo la mitad de la luz integral diaria (LID) (6.4 mol m-2 d-1 de fotones) en comparación con los tratamientos con luz LED (13.0 a 13.2 mol m-2 d-1 de fotones) (Tabla1). Además, fue 82 % menos de lo necesario para el testigo y 63 % para los tratamientos LED respecto a la LID que requiere una planta de tomate en etapa de producción (35 mol m-2 d-1 de fotones) (Kubota, 2016). Lo anterior pudo generar una baja tasa en la fotosíntesis y una baja concentración de fotosintatos en la planta para llevar a cabo un adecuado amarre de frutos. A pesar de la falta de LID, en las plantas que recibieron solo luz roja existió una mayor cantidad de frutos. Esto pudo deberse a un posible efecto de la luz roja sobre los fitocromos en el amarre de frutos, ya que están relacionados en la regulación de la floración (Berkovich et al.., 2017; Demotes-Mainard et al.., 2016).
Concentración de pigmentos fotosintéticos y compuestos fenólicos en hoja
Los resultados obtenidos en la concentración de pigmentos fotosintéticos en hoja se muestran en la Tabla 3. De manera general se observó que las plantas que recibieron 0 % de luz azul tuvieron mayor concentración de clorofila a, b y total que el resto de tratamientos con luz LED, pero sin diferencias estadísticas en comparación al testigo y una disminución de concentración de carotenoides conforme la proporción de luz azul aumentó a partir de las plantas que recibieron 76 % y 100 % de luz azul en comparación con las plantas que solo recibieron luz roja.
Tabla 3 Pigmentos fotosintéticos y compuestos fenólicos en hojas de chile habanero cultivado
bajo condiciones controladas con diferentes proporciones de luz LED
azul y roja.
Table 3. Photosynthetic pigments
and phenolic compounds in habanero pepper leaves grown under
controlled conditions with different blue and red LED light
proportions.
| Variables | Tratamientosx | |||||
| FL | A0 | A34.5 | A57.1 | A76.0 | A100 | |
| Cla | 0.90±0.08ab | 1.03±0.03a | 0.68±0.04b | 0.74±0.06ab | 0.65±0.12b | 0.65±0.06b |
| Clb | 0.50±0.02ab | 0.58±0.02a | 0.40±0.02bc | 0.39±0.02bc | 0.35±0.05c | 0.33±0.03c |
| Cltot | 1.40±0.10ab | 1.61±0.04a | 1.09±0.06b | 1.14±0.08b | 1.00±0.17b | 0.98±0.08b |
| Cla/b | 1.81±0.09ns | 1.77±0.03ns | 1.68±0.02ns | 1.87±0.04ns | 1.81±0.08ns | 1.93±0.03ns |
| Car | 4.20±0.40ab | 4.53±0.04a | 3.16±0.18ab | 3.43±0.22ab | 2.93±0.47b | 2.93±0.24b |
| CF | 29.14±1.97b | 39.27±1.14ab | 29.86±3.58b | 31.23±2.52b | 40.93±3.04ab | 53.36±4.75a |
xMedias con igual letra en las filas son estadísticamente iguales (Tukey, P ≤ 0.05). ns= No significativo; FL= fluorescente (testigo); Cla= Clorofila a (mg g-1 de peso fresco); Clb= Clorofila b (mg g-1 de peso fresco); Cltot= Clorofila total (mg g-1 de peso fresco); Cla/b= Clorofila a/b; Car= Carotenoides (mg g-1 de peso fresco); CF= Fenoles totales (mg equivalentes de ácido gálico 100 g de peso fresco).
Liu et al.. (2018) encontraron en plántulas de tomate un comportamiento similar de clorofila a, b y total, en plantas que recibieron 100% de luz azul, éstas obtuvieron la menor concentración en comparación con las que tuvieron 75 %, 60 %, 50 % y 25 % de luz respecto a luz roja. De manera contraria, Wang et al.. (2016) observaron en lechuga un comportamiento contrario, en donde plantas que recibieron solo luz roja monocromática obtuvieron la menor concentración de clorofila total, en comparación con las que recibieron 100 %, 50 %, 20 %, 11 % de luz azul respecto a la luz roja. Agarwal et al.. (2018) encontraron en espinaca un comportamiento similar a nuestros resultados respecto a la concentración de carotenoides.
El comportamiento de los pigmentos fotosintéticos en hoja, podrían explicarse en base a que la presencia de luz azul tiende a disminuir la clorofila total (Huché-Thélier et al.., 2016), y además los fitocromos, asociados a la luz roja, regulan las etapas tempranas de biosíntesis de clorofila y carotenoides (Berkovich et al.., 2017). Las plantas presentan un alto grado de plasticidad a las condiciones de luz que son expuestas, ya que al crecer bajo luz LED azul y roja, nunca recibieron longitudes de onda superiores a 680 nm, con lo cual el fotosistema II (680 nm) estará, preferentemente excitado en relación al fotosistema I (700 nm) durante todo el ciclo de crecimiento, esto no sucede en la naturaleza, pero aun así las plantas crecen y posen mecanismos de aclimatación a largo plazo que aún no se han determinado (Pocock, 2015).
La concentración de compuestos fenólicos en hojas favoreció su acumulación en plantas que tuvieron un tratamiento monocromático azul en comparación con las que recibieron 34.5% y 57.1 % de luz azul y con el testigo (Tabla 3). Jeon et al.. (2017) encontraron en Oenanthe stolonifera un comportamiento similar en la concentración de compuestos fenólicos en donde utilizaron tratamientos de luz monocromática azul y roja, al igual que diferentes proporciones de éstos.
La luz azul tiende a incrementar la síntesis y acumulación de compuestos fenólicos (Berkovich et al.., 2017), debido a que es un mecanismo de tolerancia al estrés foto-oxidativo y puede asumirse que está regulado por la producción de éste a través del fotorreceptor criptocromo, el cual está asociado con la foto activación a la luz azul (400 a 500 nm) y UV-A (320 a 400 nm) (Huché-Thélier et al.., 2016).
La presencia de luz azul y roja pueden ser responsables del aumento de la producción de metabolitos secundarios de la planta, debido a un aumento en la actividad de la enzima fenilalanina amonio liasa (PAL) la cual está involucrada en el primer paso de la vía de síntesis de fenilpropanoides (Hasan et al.., 2017). La enzima PAL se asocia con la producción de compuestos fenólicos en plantas, la cual es mayormente estimulada bajo la combinación de luz azul y roja (Heo et al.., 2012). Sin embargo, el comportamiento de los resultados puede variar debido a que no solo depende del tipo del espectro de luz que inciden en las plantas, si no, también depende de la especie, la variedad y etapa de desarrollo (Samuolienė et al.., 2012).
Concentración nutrimental en hoja
En la Tabla 4, se muestran los resultados que se obtuvieron de nutrientes en hoja chile habanero. En los macronutrientes, los diferentes tratamientos de luz no ejercieron efecto en la concentración de P en hoja, sin embargo, el tratamiento de luz monocromática roja presentó la menor concentración. Entre tratamientos con luz LED no existió diferencias estadísticas en la concentración de K, sin embargo, la concentración de K en las plantas del testigo fue mayor en comparación con las plantas que recibieron 0 % y 76 % de luz azul. La concentración de Mg en hoja fue mayor en las plantas que recibieron 0 % de luz roja en comparación con el resto de tratamientos LED y el testigo.
Tabla 4 Concentración nutrimental en hojas de chile habanero cultivado bajo condiciones
controladas con diferentes proporciones de luz LED azul y roja.
Table 4. Nutrient concentration in habanero pepper
leaves grown under controlled conditions with different blue and red
LED light ratios.
| Ele | Tratamientosx | |||||
| FL | A0 | A34.5 | A57.1 | A76.0 | A100 | |
| N | 4.69±0.07ns | 4.08±0.10ns | 4.81±0.42ns | 4.83±0.25ns | 5.30±0.30ns | 4.33±0.59ns |
| P | 0.47±0.00a | 0.33±0.00b | 0.52±0.03a | 0.49±0.03a | 0.45±0.00a | 0.53±0.02a |
| K | 5.20±0.06a | 3.62±0.10b | 3.92±0.41ab | 4.14±0.42ab | 3.75±0.19b | 4.00±0.28ab |
| Ca | 1.12±0.05ns | 1.41±0.07ns | 1.34±0.10ns | 1.32±0.05ns | 1.25±0.08ns | 1.40±0.10ns |
| Mg | 0.74±0.01b | 1.30±0.07a | 0.92±0.12b | 0.68±0.02b | 0.89±0.06b | 0.68±0.03b |
| B | 49.56±0.62ns | 52.37±1.95ns | 51.47±0.96ns | 46.47±1.46ns | 50.54±3.72ns | 59.40±5.54ns |
| Fe | 223.33±26.42ns | 166.25±13.94ns | 255.30±57.70ns | 198.93±8.19ns | 193.44±3.03ns | 244.12±22.94ns |
| Cu | 11.23±0.62a | 5.24±0.18c | 9.85±0.93ab | 10.00±0.61ab | 7.91±0.37bc | 10.78±0.59a |
| Mn | 96.850±14.41bc | 171.20±8.61a | 136.51±8.40ab | 89.02±7.00bc | 118.11±18.96abc | 77.98±1.04c |
| Zn | 136.69±7.51ns | 119.93±16.43ns | 188.35±34.12ns | 147.58±18.61ns | 119.88±12.48ns | 148.14±1.58ns |
xMedias con letras iguales en las filas son estadísticamente iguales (Tukey, P ≤ 0.05). Ele= Elementos; ns= No significativo; FL= fluorescente (testigo); N= Nitrógeno (% de peso seco); K= Potasio (% de peso seco); P= Fósforo (% de peso seco); Ca= Calcio (% de peso seco); Mg= Magnesio (% de peso seco); Fe= Hierro (mg kg-1 de peso seco); Mn= Manganeso (mg kg-1 de peso seco); Zn= Zinc (mg kg-1 de peso seco); B= Boro (mg kg-1 de peso seco); Cu= Cobre (mg kg-1 de peso seco).
En los micronutrientes, la concentración de Cu en hojas de plantas del testigo y que recibieron 100 % de azul, fue mayor que las que recibieron solo luz monocromática roja. La concentración de Mn, fue mayor las hojas de las plantas que recibieron 0 % de luz azul que las que recibieron 100 % y 57.1 % de luz azul como además que las del testigo.
Pennisi et al.. (2019) encontraron en albahaca una tendencia de mayor acumulación de nutrientes (N, P, K, Ca, Mg, Fe) con una proporción de luz azul y roja A23 %:R70 %. De manera contraria, Verma et al.. (2018) registraron en digitales que una proporción de luz azul y roja A80 %:R20 % obtuvo la máxima concentración de nutrientes (P, S, Ca, Mg, Fe, Mn, Zn, Zn, B, Cu).
La regulación de nutrientes en plantas está asociado a factores como la especie, luz, temperatura, CO2 ambiental, estrés hídrico, fitohormonas y la presencia de otros nutrientes, debido a un posible mecanismo de optimización de crecimiento al estar en diferentes condiciones de crecimiento (Sakuraba y Yanagisawa, 2017).
Sakuraba et al.. (2018) mencionan que la luz roja promueve la absorción y acumulación de fósforo en Arabidopsis debido a que el fitocromo B media el mecanismo regulatorio de absorción y uso de fósforo en la planta, a través de dos vías de señalización y regulación asociadas a la luz roja: vía-fitocromo B-HY5 y vía-fitocromo B-PIFs (PHYTOCHROME INTERACTING FACTOR family), los cuales en conjunto inducen la expresión del gen transportador de fósforo inorgánico ubicado en raíz PHT1;1 (INORGANIC PHOSPHATE TRANSPORTER) y el gen PHL1 (HOMEODOMAIN-LIKE SUPERFAMILY PROTEIN) el cual induce también la expresión del gen PHT1;1.
La menor concentración de fósforo en hoja se registró en plantas que recibieron solo luz roja (Tabla 4), una posibilidad de esto pudo deberse a la alta demanda de nutrientes y fotosintatos a la cual la planta estaba sometida, debido a que tuvo una mayor cantidad de frutos respecto al resto de tratamientos.
García-Caparrós et al.. (2019) mencionan que la concentración de potasio en plántulas de Arabidopsis se incrementó con luz azul debido a cambios en la actividad de transporte de la membrana en células guarda en estomas. Este comportamiento no se observó en ningún tratamiento con luz LED en plantas de habanero, sin embargo, se puede asumir que otras longitudes de onda pudieran estar involucradas en la absorción de potasio, dado que en las plantas que recibieron luz fluorescente, el cual tiene un espectro completo que abarca desde los 416 - 715 nm, se obtuvo mayor concentración en comparación que las plantas que recibieron 0 % y 76 % de luz azul.
El mecanismo que modula el contenido de cobre en cloroplastos se asocia con el factor de transcripción HY5, el cual es activado por la señalización generada de diversos fotoreceptores que son activados por el tipo de luz asociada a éstos, posteriormente el factor de transcripción HY5 interactúa físicamente con el factor de transcripción SPL7 (SQUAMOSA PROMOTER-BINDING PROTEIN-LIKE 7), los cuales coactivan el gen MIR408 (microRNA408) que es requerido para regular la expresión de los genes PETE1 (PLASOCYANIN 1) y PETE2 (PLASTOCYANIN 2), encargados de la síntesis de la proteína plastocianina que contiene cobre en su estructura y se ubica en cloroplastos (Gangappa y Botto, 2016; Sakuraba y Yanagisawa, 2017). Por alusión a lo anterior, en el tratamiento monocromático azul se promovió la síntesis de plastocianina asociada a cobre en plantas de habanero en comparación con las plantas que recibieron luz roja monocromática.
Miao et al.. (2019) mencionan que la luz roja promueve la acumulación de manganeso, lo cual coincidió con lo obtenido en planta de habanero en el tratamiento monocromático rojo en comparación con los resultados del tratamiento monocromático azul.
La concentración de nutrientes en tejidos de plantas puede ser modificada por factores específicos como la transpiración, apertura estomática, tasa de crecimiento y acumulación de biomasa en diferentes etapas de desarrollo (Pennisi et al.., 2019). Se ha demostrado que la luz azul está involucrada en el proceso de acumulación de nutrientes en planta a través de mecanismos que incrementan la activación de las bombas de protones y la actividad de canales iónicos en membranas celulares, con lo cual se incrementa la actividad de transporte de iones en las membranas celulares en los tejidos de plantas (Gerovac et al.., 2016). La luz azul tiene además la capacidad de regular la apertura estomática, lo cual, junto con la capacidad que tiene de incrementar el transporte de iones a través de las membranas celulares, puede afectar directamente la absorción de nutrientes mediante un aumento en la transpiración de la planta (Craver et al.., 2018). Existe poca información sobre como ciertas longitudes de onda pueden afectar la dinámica de absorción de nutrientes en plantas, por lo cual se requiere mayor investigación para descifrar estos mecanismos (Amoozgar et al.., 2016).
Variables anatómicas y viabilidad de polen
Los cortes anatómicos mostraron la existencia de óvulos bien desarrollados dentro de los ovarios, en base a la presencia de óvulos dentro del ovario, pared de ovario, núcleo del ovulo y tegumentos en todos los tratamientos evaluados. Por otra parte, hubo la presencia de polen maduro viable en anteras, en base a la existencia de una tinción completa del citoplasma del polen y de color rojo intenso en casi todos los granos de polen que se observaron en los cortes anatómicos a lo largo de todos los tratamientos evaluados (Figura 1). Estos resultados coinciden con la descripción descrita por Pérez-Pastrana et al.. (2018) en cortes anatómicos de flores en antesis para la misma variedad de chile habanero.
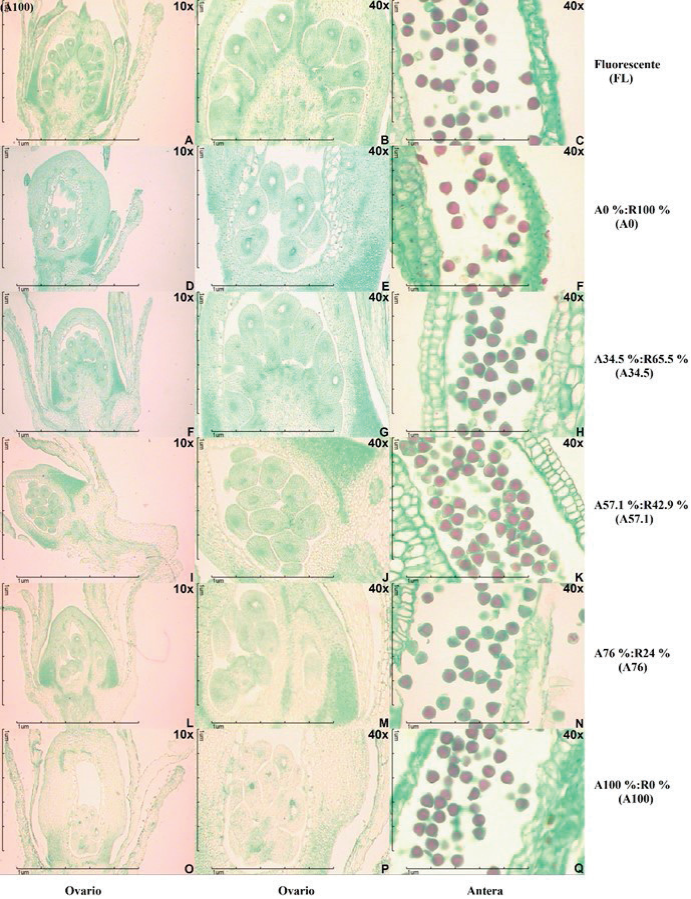
Figura 1 Cortes anatómicos longitudinales de ovario y polen de anteras de flores de chile
habanero cultivado bajo condiciones controladas con diferentes
proporciones de luz LED azul y roja.
Figure 1. Anatomical longitudinal sections of ovary and
flower anther pollen of habanero pepper grown under controlled
conditions with different blue and red LED light ratios.
El mayor porcentaje de germinación de granos de polen se presentó en las plantas del testigo, sin embargo, no existió diferencias entre los diferentes tratamientos con luz LED (Tabla 5). Dhawan y Malik (1981) estudiaron la germinación de polen in vitro en Pinus roxburghii con diferentes tipos de luz y encontraron el menor porcentaje con luz fluorescente (40 %) < luz rojo + rojo lejano (40 %) <luz azul (60 %) <oscuridad (60 %) <luz roja (95 %) y luz roja con el máximo de germinación. Hoyo et al.. (2014) investigaron la germinación de polen in vitro en Cyrtanthus mackenii con cinco picos de longitud de onda, 405 nm (violeta), 465 nm (azul), 630 nm (rojo naranja), 660 nm (rojo), 735 nm (rojo lejano) y oscuridad, concluyendo que la luz no inhibió o promovió la germinación de polen ni tampoco las longitudes de onda afectaron la tasa de germinación.
Tabla 5 Germinación de polen y grosor de hoja en chile habanero cultivado bajo condiciones
controladas con diferentes proporciones de luz LED azul y roja.
Table 5. Pollen germination and leaf thickness on
habanero pepper grown under controlled conditions with different
blue and red LED light ratios.
| Variables | Tratamientosx | |||||
| FL | A0 | A34.5 | A57.1 | A76.0 | A100 | |
| GP | 44.83±5.65a | 17.95±4.11b | 4.30±1.76b | 0.28±0.29b | 12.17±6.57b | --- |
| GH | 115.21±5.1a | 74.17±3.2d | 93.89±3.66bc | 76.67±2.43cd | 94.72±2.90b | 115.00±6.5a |
xMedias con letras iguales en las filas son estadísticamente iguales (Tukey, P ≤ 0.05). ns= No significativo; FL= fluorescente (testigo); Var= Variables; GP= Germinación de polen (%); GH= Grosor de hoja (µm).
Lo anterior sugiere que el efecto del tipo de luz sobre la germinación de polen depende de la especie; además, en chile habanero, los resultados en los tratamientos LED no presentaron diferencias estadísticas entre éstos. Por otra parte, la luz fluorescente al ser una fuente de luz de espectro completo (416 - 715 nm), promovió el porcentaje de germinación, probablemente a que posee una mayor diversidad de longitudes de onda en comparación con el resto de los tratamientos, e incluso a pesar de que la intensidad de la luz fluorescente fue la mitad en relación a estos mismos tratamientos de luz LED.
El grosor de hoja fue mayor tanto en las plantas del testigo como las que recibieron 100 % de luz azul en comparación con las que recibieron luz monocromática roja y las que tuvieron 76% de luz azul (Tabla 5). Wollanger y Runkle (2015) encontraron un comportamiento similar en salvia, ya que el grosor de hoja fue mayor en un tratamiento monocromático azul respecto a uno de rojo.
En chile habanero, el diferente grosor de hoja entre tratamientos, pudo deberse a que la luz azul tiende a incrementar el grosor de hoja respecto a la luz roja (Huché-Thélier et al.., 2016). Evert y Eichhorn (2012) mencionan que, a mayor intensidad de luz, las hojas tienden a tener un mayor grosor y viceversa; sin embargo, en el testigo a pesar de tener la mitad de intensidad luminosa respecto a los tratamientos LED tuvo el mayor grosor de hoja junto con las plantas que recibieron luz azul monocromática.
Conclusiones
Los tratamientos evaluados no indujeron un efecto en las variables de crecimiento. La presencia de luz LED roja indujo el amarre y desarrollo de los frutos, además, favoreció la frecuencia estomática.
La luz roja monocromática favoreció la acumulación de pigmentos fotosintéticos, mientras que la luz azul monocromática indujo mayor concentración de compuestos fenólicos en hoja en la mayoría de los tratamientos evaluados.
La luz roja monocromática indujo menor concentración P, y favoreció la concentración de Mg. La luz LED no generó efecto en la concentración de K, sin embargo, el testigo indujo mayor concentración en comparación con algunos tratamientos. La luz monocromática roja indujo menor concentración de Cu en hoja e indujo en Mn, mayor concentración en comparación con otros tratamientos evaluados.
El testigo incrementó la germinación de los granos de polen y el grosor de hoja, igual que la luz azul en esta última variable. La anatomía de los óvulos en ovarios y el polen en anteras no presentaron ningún efecto de los tratamientos.

