











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduccion
En las últimas décadas, las poblaciones de aves en Norte América han disminuido dramáticamente (Rosenberg et al., 2019). El grupo de las aves de pastizal no son la excepción, y la declinación de sus poblaciones es debida a cambios en el paisaje y al uso del suelo (Askins et al., 2007; Pool et al., 2014; NACBI, 2016). Se estima que el 80% de las especies de las aves del pastizal son migratorias y pasan el invierno en los pastizales del Suroeste de los Estados Unidos y Norte de México (CEC, 2013).
En ese sentido, las poblaciones de algunas especies han disminuido diferencialmente, adquiriendo estatus distintos de importancia para su conservación y requiriendo un esfuerzo trinacional entre Canadá, Estados Unidos y México (Berlanga et al., 2010). Por ejemplo, las poblaciones de Centronyx bairdii (gorrión de Baird), Ammodramus savannarum (gorrión chapulín) y Anthus spragueii (bisbita llanera) han enfrentado una disminución anual de -2.1%. -2.5% y -4.2% respectivamente (Sauer et al., 2017). Estas especies, consideradas como aves focales de pastizal, se reproducen en las praderas de pastos cortos en Estados Unidos de América (Askins et al., 2007).
Al respecto, se han realizado estudios en este grupo de aves, en diferentes áreas de pastizal del desierto chihuahuense mexicano que tratan de incrementar el conocimiento acerca de las causas probables en la disminución poblacional con respecto de a) abundancia y distribución (Martínez-Guerrero et al., 2011; CEC, 2013), b) sobrevivencia invernal (Macías-Duarte et al., 2017), c) uso de hábitat (Pool et al., 2012; Strasser et al., 2019), y d) precipitación pluvial y cobertura vegetal (Macías-Duarte et al., 2018).
No obstante, el grupo de aves de pastizal requiere especial atención debido a la estimación del 52% de pérdida en los últimos 48 años (Rosenberg et al., 2029). Lo anterior ha estimulado el interés por el estudio de estas aves y la búsqueda de información básica que permitan diseñar estrategias de conservación exitosas tanto en el presente como hacia el futuro (Comer et al., 2018, Wilsey et al., 2019),
Por otro lado, y de acuerdo con Subirós (2006), los métodos cuantitativos en ecología del paisaje proveen de información numérica sobre la situación de la composición de la cobertura terrestre lo que representa un panorama sobre las características morfológicas de la cobertura vegetal que puede ser clave para interpretar las condiciones y sus procesos ecológicos tanto del punto de vista espacial como temporal (Burel y Baudry, 2002).
El objetivo del presente trabajo fue estudiar la densidad invernal de Centronyx bairdii, Ammodramus savannarum, y Anthus spragueii y su relación con los atributos de la vegetación y cobertura de suelo en el Área Prioritaria para la Conservación de Pastizales (CEC y TNC, 2005) denominada Cuchillas de la Zarca.
Los resultados obtenidos aportan al conocimiento del hábitat invernal de las especies estudiadas a gran escala, o que es necesario para señalar los lugares del área de estudio que deben atenderse con algunas estrategias para conservación.
Materiales y métodos
Área de estudio
El presente estudio se desarrolló en el Área Prioritaria para la Conservación de Pastizales (APCP) denominada Cuchillas de la Zarca, la cual forma parte del Desierto Chihuahuense en México y comprende una porción pequeña del sur del estado de Chihuahua y el centro -norte del estado de Durango; cuenta con una superficie de 11,588 km2 ubicada entre las coordenadas geográficas extremas 25° 20’ 00” y 27° 00’ 00” de latitud Norte y 105° 50’ 00” y 104° 25’ 00” de longitud Oeste (Figura 1).
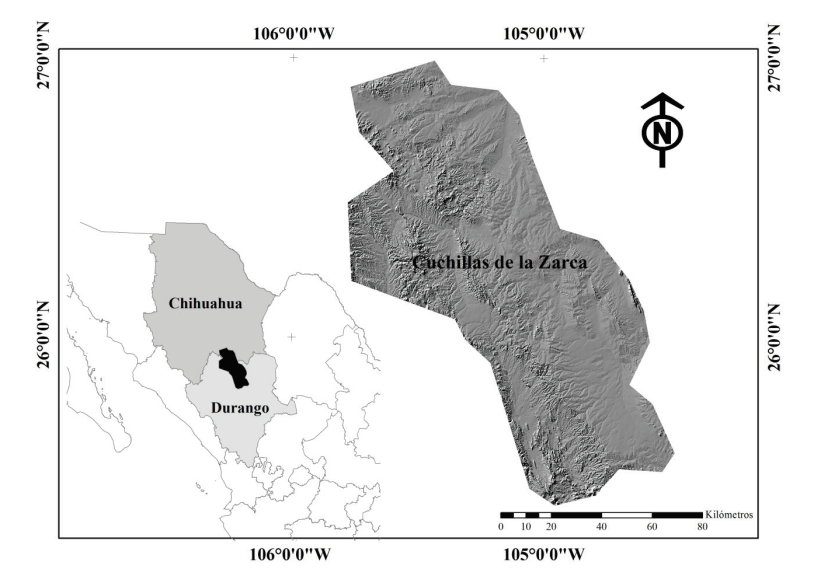
Figura 1 Localización del área de estudio: Área Prioritaria para la
Conservación de Pastizales (APCP) Cuchillas de la Zarca
Figure 1. Location of the study area: Priority
Grassland Priority Conservation Area of Cuchillas de la
Zarca.
En la zona de estudio se presentan tres tipos de clima, el semiárido-templado (BS1kw), en la parte central de la región, con temperatura media anual entre 12°C y 18°C, temperatura del mes más frío entre -3°C y 18° C, temperatura del mes más caliente menor de 22°C; lluvias de verano del 5% al 10.2% anual, en lugares con altitudes mayores a los 1,800 m.s.n.m. El árido-templado (BSokw) en las laderas de la Sierra madre Occidental, se caracteriza por temperaturas entre 12°C y 18°C, temperatura del mes más frío entre -3°C y 18° C, temperatura del mes más caliente menor de 22°C; lluvias de verano del 5% al 10.2% anual. En el clima muy árido -semicálido (BWhw) en la parte oriental de la zona, donde las temperaturas fluctúan entre 18°C y 22°C, la temperatura del mes más frío menor de 18° C, temperatura del mes más caliente mayor de 22°C y lluvias de verano del 5% al 10.2% anual (Vidal-Zepeda, 2005).
La vegetación en el área de estudio se compone de matorral xerófilo y pastizal natural (Rzedowski, 1981). El estrato arbustivo está conformado por especies de los géneros Larrea, Prosopis, Acacia, Ephedra, Nolina, Opuntía, Juniperus y Quercus; el estrato herbáceo por los géneros Bouteloa,Aristida, Buchloe, Andropogon, Melinis, Muhlenbergia, Sporobolus, Heteropogon y Pleuraphis.
Esta región prioritaria para la conservación incluye el extremo sur de los pastizales asociados a la vertiente oriental de la Sierra Madre Occidental en los estados de Chihuahua y Durango. La región se delimita hacia el oeste y el norte por subcuencas donde el parteaguas llega hasta la cota de los 2,000 msnm que coincide con bosques bajos-abiertos de encinos, y hacia el este y sur con pastizales naturales. La región alberga los pastizales naturales, matorrales xerófilos y las zonas ecotonales de la Sierra Madre Occidental (piedemonte) como son los chaparrales y los bosques bajos-abiertos (Martínez et al., 2011). Los diferentes tipos de suelo desarrollan y mantienen pastizales con diferente composición de especies, los cuales dan origen a una riqueza de aves alta.
Estimación de densidades de especies de aves de pastizal y variables de vegetación
El monitoreo de aves y vegetación lo realizamos usando la metodología descrita por Panjabi et al. (2010); se elaboró una malla de cuadrantes de 18 km2 que cubrieron la totalidad de la superficie del APCP (CEC y TNC 2005). Del total de cuadrantes (n =36) se seleccionaron aquellos donde ocurre pastizal nativo (n=16). Además, verificamos que estos cuadrantes contaran con caminos disponibles (más de 5 km), en los cuales se ubicaron cada 500 m sitios de muestreo que fueron seleccionados de forma aleatoria (Figura 2).
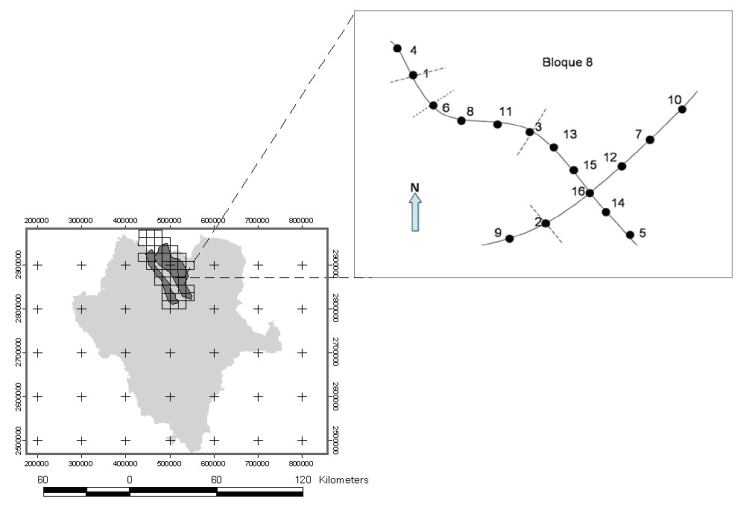
Figura 2 Esquema del monitoreo de aves y vegetación (Panjabi
et al., 2010)
Figure 2. Monitoring scheme of birds and vegetation
(Panjabi et
al., 2010).
En los primeros tres sitios seleccionados en cada cuadrante de manera aleatoria (1-3) ubicamos transectos para monitoreo de aves (Panjabi et al., 2007) realizados por dos observadores capacitados para reconocer aves de pastizal. Con apoyo de un navegador GPS Garmin Etrex Vista®, cada observador realizó recorridos a pie en transectos en cada sitio de muestreo de un kilómetro de longitud. Los transectos fueron perpendiculares al camino y para registrar a las aves observadas y determinar su distancia al transecto, se usaron binoculares Eagle Optics® 8x42 mm y distanciómetros láser marca BUSHNELL®.
El monitoreo lo realizamos entre el 20 de enero y el 21 de febrero de los años 2009, 2010 y 2011 entre las 08:00 y 12:00 horas; no se trabajaron los días con mucho viento, lluvia o nieve. La distancia perpendicular al transecto la utilizamos para calcular la densidad de aves por km2, para las especies Centronyx bairdii (BAIS), Ammodramus savannarum (GRSP) y Anthus spragueii (SPPI) mediante el uso del programa DISTANCE® 6.0 release 1 (Buckland et al., 2001), con las opciones de muestreo convencional (CDS) por transecto lineal simple, bajo el modelo medio-normal para la función de detección.
Inmediatamente después de terminar el monitoreo de aves efectuamos el muestreo de vegetación cada 100 m, en sentido inverso a la dirección al punto de origen del camino sobre el mismo transecto de aves, utilizando círculos de 5 m de radio para estimar visualmente cobertura de suelo, registrando la cobertura (%) de pasto, hierba, suelo desnudo y otro tipo de cobertura (vegetación muerta), y altura (cm) de pasto, hierba y arbustos (m) (Macías-Duarte et al., 2017).
Estimación del Índice de Vegetación Diferencial Normalizado
Mediante el programa computacional ArcGis 10.1 calculamos el Índice de Vegetación Diferencial Normalizado, derivado del procesamiento e interpretación de imágenes de seis escenas del sensor Landsat-5/TM (path/row31-41 y 31-42) con fechas de captura del 2 al 24 febrero, correspondiendo al periodo de adquisición del 2009, 2010 y 2011.
Análisis estadístico
La variable de densidad de las aves no cumplió el supuesto de normalidad por lo seleccionamos estadística no paramétrica, con el fin de comprobar la existencia de diferencias observadas de las densidades mediante una prueba de permutación (10,000) basada en reasignaciones elegidas al azar (Manly, 1997). Para calcular los promedios y el error estándar de las variables de vegetación por año y por cuadrante, usamos estadística univariada mediante el paquete estadístico SAS/STAT® (SAS Institute Inc. 2004), con los procedimientos PROC UNIVARITE y PROCFORMAT.
Dado que no se halló linealidad o una distribución normal de los datos, la relación entre la densidad relativa de aves por bloque y las variables de estructura vegetal por bloque, la estimamos por la prueba de covariación (C) descrita por Gregorius et al. (2007). Por definición, las dos variables ordinales X yY muestran toda covariación si una variable aumenta constantemente o disminuye a medida que las demás variables aumentan constantemente. Por tanto, existe una relación estrictamente monótona (pero no necesariamente lineal) entre las dos variables. La covariación C oscila entre 1 y -1, donde C = 1 corresponde a una covariación totalmente positiva y C = -1 a una covariación estrictamente negativa. Si su denominador es cero, C es indefinida (Gregorius et al., 2007). Formalmente:
donde:
C = covariación entre cada variable X y especie Y.
(Xi-Xj) = Diferencia en los valores de las variables de estructura vegetal X, entre el i-ésimo y las variablesj-ésimo.
(Yi-Yj) = Diferencia en valor de la variable densidad de aves Y, entre el i-ésimo y las variables j-ésimo.
El denominador de la ecuación, es la suma de los valores absolutos de los productos indicados. Con el fin de probar la probabilidad de que los grados observados de covariación C se produzcan sólo por sucesos aleatorios en lugar de fuerzas dirigidas, se realizó una prueba de permutación de dos caras, realizando reasignaciones elegidas al azar, de acuerdo con Manly (1997). Los porcentajes de C imitado mayor que o igual a los respectivos C (valores de p) observados se calcularon para un número satisfactorio de permutaciones.
A su vez, también se aplicó la prueba de Kruskal-Wallis ya que representa una excelente alternativa no paramétrica al ANOVA de un factor completamente aleatorizado, para comparar las variables de vegetación entre especies por año de estudio y para comparar las densidades de cada especie por año.
Resultados
En el Área Prioritaria para la Conservación de Pastizales Cuchillas de la Zarca, durante los inviernos de los años 2009, 2010 y 2011, monitoreamos las especies de aves de pastizal Centronyx bairdii, Ammodramus savannarum y Anthus spragueii en 96 transectos de 1 km de longitud, 6 transectos por cuadrante (n=16). La densidad de Centronyx bairdii fue de 50.8, la de Ammodramus savannarum de 76.7 y la de Anthus spragueii de 12.6 (aves/km2) en promedio (Tabla 1).
Tabla 1 Densidad (ind./km2 promedio ± e.e) de tres especies de aves de pastizal en Cuchillas de
la Zarca años 2009-2011.
Table 1. Density (ind./km2 mean ± e.e) of three grassland
birds species in Cuchillas de la Zarca 2009-2011.
| Año | ||||
|---|---|---|---|---|
| Especie | 2009 | 2010 | 2011 | promedio |
| BAIS | 67.7±1.40 | 39.6±0.70 | 45.1±1.04 | 50.8 |
| GRSP | 90.8±1.77 | 73.6±1.49 | 65.7±1.29 | 76.7 |
| SPPI | 18.1±0.55 | 10.3±0.44 | 6.80±0.27 | 12.6 |
Centronyx bairdii (BAIS), Ammodramus savannarum (GRSP) y Anthus spragueii (SPPI)
*American Ornitologhist Union Bird Codes (Pyle y DeSante, 2003).
Los resultados del análisis de variables de la vegetación invernal estimada y analizada mediante la prueba no paramétrica de Kruskal-Wallis, para comparar entre especies por cada año, observamos diferencias significativas para los años 2010 y 2011 entre los gorriones (BAIS, GRSP) que se encontraron con una cobertura de pasto mayor que la bisbita llanera (SPPI), la cual tolera mayor cobertura de hierba (Tabla 2). Derivado de la prueba de comparación múltiple de medias entre las densidades relativas de las tres especies de aves focales de pastizal, se encontró que solo existe diferencia significativa (P ≤ 0.05) entre los años de 2009 y 2011 para la especie Anthus spragueii, especie para la que su densidad relativa disminuye entre años (Tabla 3).
Tabla 2 Valores promedio de variables de la vegetación invernal (hábitat) para tres especies de
aves focales de pastizal durante tres años consecutivos (2009-2011) en
la APCP de Cuchillas de la Zarca, comparación entre especies por año
(Kruskall - Wallis).
Table 2. Average of winter vegetation variables (habitat)
for three grassland focal bird species through three consecutive years
(2009-2011) in the GPCA of Cuchillas de la Zarca, comparison between
species per year (Kruskall – Wallis).
| Especie | Altura arbustos (m) | Altura pastos (cm) | Altura hierba (cm) | Suelo desnudo (%) | Cobertura arbusto (%) | Cobertura pasto (%) | Cobertura hierba (%) | Otra cobertura (%) |
|---|---|---|---|---|---|---|---|---|
| (1) BAIS | 1.24a | 22.00a | 14.34a | 12.82a | 4.76b | 63.72a | 10.27b | 13.24a |
| (1) GRSP | 1.23a | 20.96a | 14.65a | 14.26a | 6.38a | 60.25b | 11.28a | 14.35a |
| (1) SPPI | 1.15a | 21.68a | 14.81a | 13.77a | 7.03a | 60.40b | 10.98ab | 14.90a |
| (2) BAIS | 1.29a | 19.97a | 13.25b | 16.74a | 3.85a | 52.85a | 6.75b | 18.60a |
| (2) GRSP | 1.36a | 19.55a | 12.78b | 17.49a | 3.95a | 53.01a | 6.22b | 18.56a |
| (2) SPPI | 1.28a | 19.74a | 15.10a | 17.95a | 3.98a | 48.57b | 8.71a | 18.99a |
| (3) BAIS | 1.63a | 20.22a | 15.83b | 16.71b | 4.51b | 55.37a | 7.00b | 16.13a |
| (3) GRSP | 1.57a | 19.16ª | 15.10b | 16.78b | 4.30b | 55.30a | 6.64b | 16.44a |
| (3) SPPI | 1.50a | 20.16a | 17.57a | 19.08a | 6.45a | 49.15b | 9.09a | 15.39a |
(1)= Datos del año 2009, (2) = Datos del año 2010, (3) = Datos del año 2011 Literales distintas entre columnas por especie son estadísticamente diferentes (Z≥1.96)
Centronyx bairdii (BAIS), Ammodramus savannarum (GRSP) y Anthus spragueii (SPPI)
*American Ornitologhist Union Bird Codes (Pyle y DeSante, 2003).
(1)= Data for the year 2009, (2) = Data for the year 2010, (3) = Data for the year 2011
Different letter between columns by species are statistically different (Z≥1.96)
Tabla 3 Comparación múltiple de medias de la densidad relativa de las tres especies de aves
focales de pastizal en Cuchillas de la Zarca años 2009-2011.
Table 3. Multiple comparison of means of the relative
density of the three species of grassland focal birds in Cuchillas de la
Zarca, years 2009-2011.
| Año | BAIS | GRSP | SPPI |
|---|---|---|---|
| 2009-2010 | 0.0650 | 0.2410 | 0.0699 |
| 2009-2011 | 0.1289 | 0.1341 | 0.0215* |
| 2010-2011 | 0.3382 | 0.3362 | 0.1896 |
*Asociación con valores de P ≤ 0.05 son significativas.
Centronyx bairdii (BAIS), Ammodramus savannarum (GRSP) y Anthus spragueii (SPPI)*American Ornitologhist Union Bird Codes (Pyle y DeSante 2003).
Tabla 4 Covariación (C) de Gregorius entre la densidad de tres especies de aves y atributos de la vegetación por año (2009-2011) en el Área Prioritaria para la Conservación de Pastizales de Cuchillas de la Zarca.
| Atributos de vegetación | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Especies | Altura arbustos (m) | Altura pastos (cm) | Altura Hierba (cm) | Suelo desnudo (%) | Cobertura arbusto (%) | Cobertura pasto (%) | Cobertura hierba (%) | Otra cobertura (%) | NDVI |
| (1) BAIS | -0.49 | 0.68 | 0.64 | -0.60 | -0.59 | 0.68 | -0.33 | -0.64 | 0.44 |
| (2) BAIS | -0.61 | 0.77* | 0.34 | -0.88* | -0.73* | 0.69 | -0.06 | 0.04 | 0.32 |
| (3) BAIS | -0.37 | 0.92* | 0.29 | -0.89* | -0.60 | 0.80* | 0.92 | -0.23 | 0.37 |
| (1) GRSP | -0.36 | 0.85* | 0.85* | -0.64 | -0.37 | 0.54 | 0.27 | -0.62 | 0.29 |
| (2) GRSP | -0.53 | 0.95* | 0.16 | -0.76* | -0.76* | 0.73* | -0.07 | -0.26 | 0.38 |
| (3) GRSP | -0.21 | 0.79* | -0.08 | -0.86* | -0.66 | 0.85* | -0.55 | -0.24 | 0.35 |
| (1) SPPI | -0.31 | 0.09 | 0.47 | 0.19 | 0.69 | -0.30 | 0.47 | 0.14 | 0.10 |
| (2) SPPI | -0.49 | -0.18 | 0.78* | 0.45 | 0.00 | -0.85* | 0.96* | 0.52 | 0.50 |
| (3) SPPI | -0.33 | -0.07 | 0.58 | 0.46 | 0.62 | -0.57 | 0.53 | -0.39 | 0.10 |
(1) = Datos del año 2009, (2) = Datos del año 2010, (3) = Datos del año 2011, NDVI = Índice de Vegetación de Diferencia Normalizada. Centronyx bairdii (BAIS), Ammodramus savannarum (GRSP) y Anthus spragueii (SPPI)
*American Ornitologhist Union Bird Codes (Pyle y DeSante 2003).
* En negrita son los valores de C que fueron significativos con P ≤0.025.
(1)= Data for the year 2009, (2) = Data for the year 2010, (3) = Data for the year 2011, NDVI = Normalized Difference Vegetation Index. Centronyx bairdii (BAIS), Ammodramus savannarum (GRSP) and Anthus spragueii (SPPI)
* American Ornitologhist Union Bird Codes (Pyle and DeSante 2003).
* Bold are the values of C that were significant with P ≤ 0.025.
Los valores de covariación de Gregorius (C) muestran que, de las especies estudiadas, tienen una correlación significativa (P <0.025) con al menos seis variables de atributos de vegetación invernal (Cuadro 4). Tanto BAIS, como GRSP presentaron relaciones de valor alto, positivas y significativas, simultáneamente con las variables: altura de pastos (cm) y cobertura de pastos, mientras que para la variable suelo desnudo, su relación fue significativa, igualmente alta pero negativa. La especie SPPI sólo tuvo relación significativa negativa con la variable cobertura de pastos en el año 2010 y relaciones significativas positivas con altura de hierba y cobertura de hierba. La densidad de las especies en los años de estudio no presentó ninguna relación con el NDVI.
Discusión
Los resultados de densidad (individuos /km2) para Centronyx bairdii (BAIS) (Cuadro 1), son similares a los reportes previos de Martínez et al. (2011) quienes estiman un promedio de 59.8 ind./km2, al igual que los resultados de Macías-Duarte et al. (2011) quienes calculan 47.04 ind./km2 para los años 2007-2011 ,y al dato reportado por la Comisión para la Cooperación Medio ambiental (CEC 2013) con 47.27 aves/km2 para los años 2007-2013; esta fuente menciona además la estimación de 335,00 aves de esta especie, en el área de estudio durante el invierno.
Para el caso del gorrión chapulín (GRSP), el promedio estimado para los tres años fue de 76.7 ind./km2. Macías-Duarte et al. (2011), reportaron una densidad de 84.14 ind/ km2 para la misma área de este estudio y CEC (2013) reporta 74.19 ind./ km2 para todas las áreas de estudio en el Desierto Chihuahuense.
La bisbita llanera (SPPI), mostró un decremento en su detección y por consecuencia en la densidad estimada anualmente (Cuadro1); sin embargo, el promedio de aves estimado para los tres años fue de 12.6 ind./km2, superior al reporte de Macías-Duarte et al. (2011) de 4.27 ind./km en la misma APCP y a los 6.36 ind./km2, reportado por la Comisión para la Cooperación Ambiental (2013) para todas las áreas prioritarias para la conservación de pastizales ubicadas en el Desierto Chihuahuense.
Las tres especies las encontramos en más del 50% de los cuadrantes del estudio en el APCP de Cuchillas de la Zarca, y en general los valores de densidad de aves que estimamos tuvieron una reducción entre años (2009-2011), probablemente debido al efecto de la precipitación pluvial (variable no estudiada) durante el verano que tiene efecto demostrado sobre la abundancia de las aves de pastizal en el Desierto Chihuahuense (Macías-Duarte et al., 2018).
El gorrión de Baird se localizó en sitios donde la cobertura de pasto (52% en promedio), la cobertura de hierba (8 % en promedio) y la cobertura de arbustos (4.3% en promedio) fueron mayores a los reportes de Macías-Duarte et al. (2011) de 31%, 1.0% y 1.0% respectivamente en todas las APCP del desierto Chihuahuense durante los inviernos 2013-2014. Esos datos son similares al reporte de Martínez et al. (2011); sin embargo, el promedio de altura del pasto de nuestro estudio (20.73 cm) fue inferior a los 24.6 cm, observado por Macías-Duarte et al. (2018), donde tal vez esta especie focal de pastizal compense un poco la falta de cobertura por la altura del pasto en algunos sitios de invernada, del desierto Chihuahuense.
Para el gorrión chapulín observamos que se encontró en lugares donde la cobertura de pasto fue superior a 56% con una altura promedio de 19.89 cm., y de igual manera mantiene la misma tendencia que en el gorrión de Baird, efecto provocado por la escala del presente estudio, en comparación con los datos de 45% de cobertura y 24 cm de altura de pasto estimados por Macías-Duarte et al. (2017), para el APCP de Janos en Chihuahua.
Se conoce que para ambas especies de gorriones, la cobertura y altura de pasto tiene relación con la sobrevivencia invernal, al proveer refugio contra el frío y depredación y además tiene influencia en el tamaño del territorio (Sierra et al., 2019).
La bisbita llanera (SPPI) se caracterizó por encontrarse en lugares con una menor cobertura de pasto con respecto a los gorriones de Baird y chapulín; sin embargo, es una especie que se relaciona positivamente con esta variable como lo demostró Pool et al. (2012), bajo las mismas condiciones de estudio que las de nuestro reporte. No obstante, en nuestro trabajo (macro hábitat), los valores reportados para esta variable (49%) son superiores a los estimados en un trabajo a nivel micro hábitat (Strasser et al., 2019, en prensa) con ayuda de telemetría estimaron que esta especie utiliza sitios con una cobertura de pasto promedio de 35.7%; además, encontraron que la variable de mayor peso en la selección de hábitat, fue la preferencia por lugares con mayor proporción de suelo desnudo (55%), valor muy superior al de nuestro reporte.
La bisbita llanera usa lugares donde pueda observar la presencia de depredadores, ya que huye en vez de cubrirse (esas características de vegetación le permiten incrementar la posibilidad de sobrevivencia).
Con respecto a la relación estimada entre los atributos de la vegetación invernal y las diferentes especies de aves, durante los años del estudio mediante la prueba de covariación de Gregorius (C), Martínez et al. (2011 y 2014) encontraron relaciones positivas para BAIS con altura y cobertura de pastos y hierba, así como relaciones negativas con suelo desnudo y otra cobertura, que concuerdan con el reporte de Pool et al. (2012), quienes señalan que la densidad de aves de esta especie en el desierto chihuahuense está relacionada positivamente a la cobertura de pasto.
Para el gorrión chapulín (GRSP), nuestros resultados demuestran que la especie está positivamente asociada con la altura de y cobertura de pastos y de manera negativa con la cobertura de arbustos y al porcentaje de suelo desnudo, aspecto igualmente reportado por Macías-Duarte et al., (2009) quienes observaron una relación positiva con las mismas variables en zonas del Sueco y Tinaja Verde en Chihuahua.
La bisbita llanera (SPPI) en nuestro reporte, está asociada negativamente con la altura del pasto y cobertura de pasto; sin embargo, Macías-Duarte et al. (2009) encontraron que se relaciona de manera positiva con la cobertura de pasto (concordando con los hallazgos de Pool et al., (2012)) considerando la obstrucción visual del pasto alto como una variable importante en la selección del hábitat invernal de esta especie. Sin embargo, un reciente estudio a nivel de microhabitat indica que SSPI tiene una fuerte preferencia con lugares con una alta proporción de suelo desnudo (Strasser et al., 2019).
En el estudio reciente de sobrevivencia invernal en estas especies de aves, Macías-Duarte et al. (2017), encontraron que algunas características del hábitat (cobertura y estructura) están asociadas con la posibilidad de sobrevivencia invernal, principalmente y de manera negativa con la altura de los arbustos (≥ 50 cm); ellos explican que estas estructuras son usadas como sitios de percha y caza de depredadores.
El análisis del NDVI con imágenes satelitales y su posible relación con las especies de aves no arrojó ningún resultado significativo y no fue posible establecer diferencias a la escala utilizada por nosotros, además de que es importante señalar que los valores de reflectancia de la vegetación en época invernal son mínimos por la baja actividad fotosintética.
Sin embargo, se puede observar que los valores de NDVI para el año 2010, fueron ligeramente mayores que los otros años de estudio, y pensamos que hipotéticamente, esta sea una consecuencia de la mayor precipitación pluvial en 2009, año en el que la densidad de aves fue la mayor en las tres especies.
Conclusiones
Estimamos las densidades invernales de tres especies de aves de pastizal en el APCP de Cuchillas de la Zarca, con resultados similares a otras investigaciones en la misma zona, pero mayores a otras APCP en el desierto Chihuahuense; el área de estudio destaca de forma importante como sitio de invernada de estas especies en el norte de México.
El análisis de la relación entre los atributos de la vegetación invernal y la densidad de las tres especies de aves de pastizal, utilizando un análisis no convencional como Covariación de Gregorius, y sus resultados son concordantes con otros trabajos que se han publicado al respecto, por lo que este método puede ser útil para estudios futuros a esta escala. La característica cobertura de pastos es muy importante para las especies Centronyx bairdii y Ammodramus savannarum; en Cuchillas de la Zarca se estimó que presentan valores mayores que en otras áreas del desierto Chihuahuense, quizá porque la estructura del paisaje (parches de pastizal de tamaño variable) representa un hábitat más apropiado para estas especies y el cual es seleccionado, teniendo a su vez tiene influencia en la densidad de estas especies.
Las especies de gorriones del presente estudio, que son reconocidas como aves focales de pastizal, mostraron preferencia por sitios con mayor altura y cobertura de pastos, y menor cobertura de suelo desnudo y arbustos; mientras que Anthus spragueii, sólo mostró una relación positiva con la altura y cobertura de hierbas y negativa con la cobertura de pasto, debido a que prefiere áreas más abiertas que le permiten vigilar de manera constante la presencia de depredadores.
Los resultados obtenidos indican que estas especies de aves dependen de una buena cobertura y altura de pastos y sugieren una relación negativa con la cobertura de arbustos: estos datos pueden ayudar a la implementación de estrategias de conservación del hábitat de las aves de pastizal orientadas en ese sentido, que permitan mitigar su declive a nivel continental.

