











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Cystic fibrosis (CF) is a genetic disorder that mainly affects the lungs but also the pancreas, liver, kidney, and intestine. This disease is caused by mutations in the cystic fibrosis transmembrane conductance regulator gene (CFTR)1. Mutations in CFTR alter the transport of chloride and sodium ions, HCO3- and water across the cell membrane in the airways.
Lung epithelia with impaired CFTR gene function produce thick, sticky mucus that clogs the airways and traps opportunistic bacteria, producing infections that cause inflammation, leading to decreased lung function, respiratory distress, and eventually respiratory failure2.
During the first years of life, the airways of children with CF are rapidly colonized by non-typeable Haemophilus influenzae and Staphylococcus aureus3, and progressively by Pseudomonas aeruginosa and Burkholderia cepacia complex, which are the primary opportunistic pathogens associated with chronic infection and decreased pulmonary function3. In Mexico, most children with CF are colonized by P. aeruginosa and S. aureus4. Worldwide, airway colonization by S. aureus ranges between 30-50%5.
S. aureus is a Gram-positive bacterium associated with establishing an inflammatory process in the lower respiratory tract, reducing lung function and contributing significantly to lung tissue damage6. The ability of these bacteria to produce a biofilm is associated with increased resistance to antibiotics in vitro7. In particular, methicillin-resistant S. aureus (MRSA) has been associated with accelerated deterioration of lung function and increased mortality8. Methicillin resistance is attributed to an alternate penicillin-binding protein (PBP2a or PBP2’) encoded by the mecA gene9. S. aureus is a versatile bacterium with an arsenal of virulence factors, including Panton-Valentine leukocidin (PVL), which facilitates tissue adhesion and host cell injury10. PVL is often related to community-associated MRSA (CA-MRSA)11, and its expression has been associated with severe infections, bacteremia, osteomyelitis, and necrotizing pneumonia12. PVL is a bicomponent pore-forming cytotoxin that causes leukocyte lysis13. This study aimed to characterize the genetic relatedness, presence of MRSA, and macrophage cytotoxic activity in clinical isolates of S. aureus from a cohort of children with CF in Mexico.
Methods
We conducted a descriptive study derived from a study published in 20204. This study was approved by the Institutional Review Board of the Faculty of Medicine of the Universidad Nacional Autónoma de México (Protocol FMED/CI/RGG/022/2016).
Bacterial isolates
Bacterial isolates were obtained from sputum samples collected from 50 pediatric patients attending the CF clinic of the Hospital Infantil de México Federico Gómez (HIMFG), Mexico City, Mexico, from August 2016 to January 20184. Parents of children with CF who agreed to participate in this study signed a consent form authorizing the collection of sputum samples to identify bacterial pathogens. Patient samples were collected as part of routine hospital care. Each patient had appointments scheduled every 3 to 6 months, although some patients and their parents do not always attend their scheduled appointments at HIMFG.
Sputum samples or cough swabs were transported to our laboratory for processing. Sputum samples were dissolved weight/volume (1:1) in sputolysin (Merk-Millipore, Darmstadt, Germany) for 30 minutes at 37oC. Dissolved sputum samples and cough swabs were used to inoculate salt and mannitol, chocolate, blood, MacConkey (DIBICO, State of Mexico, Mexico), and cetrimide (Becton Dickinson, New Jersey) agar media plates. Plates were incubated at 37oC for 24 hours. Chocolate agar and blood agar plates were also incubated at 37oC for 24 hours under microaerophilic conditions. Bacterial isolates were identified by standard microbiological methods4. All samples were stored at −70oC until analysis.
Pulsed-field gel electrophoresis
The bacterial relatedness of 50 clinical isolates of S. aureus was determined by pulsed-field gel electrophoresis (PFGE). Bacterial genomic DNAs were purified and prepared as described elsewere14. Genomic DNAs were digested with SmaI (Invitrogen) for 24 h and resolved by PFGE using a Gene Path system (BioRad®USA). Bacterial relatedness among S. aureus clinical isolates was determined according to the Tenover criteria and the use of the Dice coefficient, as previously described4. An isolate was considered a member of the same pulsotype when it had a > 85% correlation.
Antibiotic susceptibility testing
S. aureus isolates were tested for antimicrobial susceptibility to ciprofloxacin, levofloxacin, moxifloxacin, gentamicin, tigecycline, trimethoprim/sulfamethoxazole, oxacillin, erythromycin, clindamycin, linezolid, vancomycin, tetracycline, and rifampicin. The minimum inhibitory concentrations (MICs) were determined using the VITEK®2 system (bioMérieux®SA). Methicillin resistance was determined using the cefoxitin test by the disk diffusion method (Kirby-Bauer). Quality control and interpretation of results were performed according to Clinical and Laboratory Standards Institute (2019) guidelines15. S. aureus ATCC 43300, 25923, and USA300 were used as standard quality controls. The multidrug resistance (MDR) phenotype is defined as non-susceptibility to ≥ 1 agent from ≥ 3 antimicrobial categories16.
DNA extraction and identification of mecA and lukPV genes by polymerase chain reaction (PCR)
S. aureus isolates were grown on salt and mannitol agar plates for 24 hours at 37oC. An isolated colony was resuspended in 100 mL of MilliQ water and boiled for 10 minutes. The bacterial suspension was centrifuged at 10,000 rpm at 4oC for 10 minutes. Supernatants containing genomic DNA were used for PCR reactions. Primers mecA(F): 5’-TGGCTATCGTGTCACAATCG-3’ and MecA(R): 5’-CTGGAACTTGTTGAGCAGAG-3’ were used for mecA amplification, whose amplification product is 310pb17. The lukPV gene was amplified with primer LukPV(F): 5’-ATCATTAGGTAAAATGTCTGGACATGATCCA-3’ and LukPV(R): 5’-GCATCAACTGTATTGGATAGCAAAAGC-3’, with an amplification product of 433pb18.
PCR was performed as follows: we mixed 10 mM of each primer, 1X GoTaq® Green Master Mix, and 3 mL of DNA template extracted by boiling. The conditions for gene amplification were one cycle at 94oC for 5 min, 30 cycles at 94oC for 30 s, 55oC for 30 s, and 72oC for 90 s with a final extension step at 72oC for 5 min. PCR products were resolved on a 1% (w/v) agarose gel for one hour at 100 volts. The gel was stained with ethidium bromide (1 mg/mL) for 5 min and washed twice in deionized water. The gel was analyzed on a UV light transilluminator using Quantity One software (BioRad® USA).
Reagents, cells, and growth conditions
Fetal bovine serum (FBS) and RPMI-1640 cell culture media were obtained from Invitrogen, PBS and Luria-Bertani (LB) broth from Sigma-Aldrich, and salt and mannitol agar from DIBICO (State of Mexico, Mexico). Human THP1 cells were obtained from ATCC® TIB-202; 3x105 human THP1 cells were differentiated into macrophages (THP1m) using PMA 100 ng/mL (Sigma-Aldrich) for 24 hours19. Prior to infection, the medium was changed to RPMI medium with no antibiotics. THP1m cells monolayers were infected with S. aureus at an MOI of 50. To synchronize the infection, we centrifuged the cells at 1,200 rpm for 1 min and then incubated the plates for one hour. After infection, the cells were washed three times with PBS to remove extracellular bacteria; infected cells were returned to the incubator for an additional 24 h in RPMI medium supplemented with gentamicin (100 mg/mL).

Results
Presence and genotyping of S. aureus isolates from CF pediatric patients
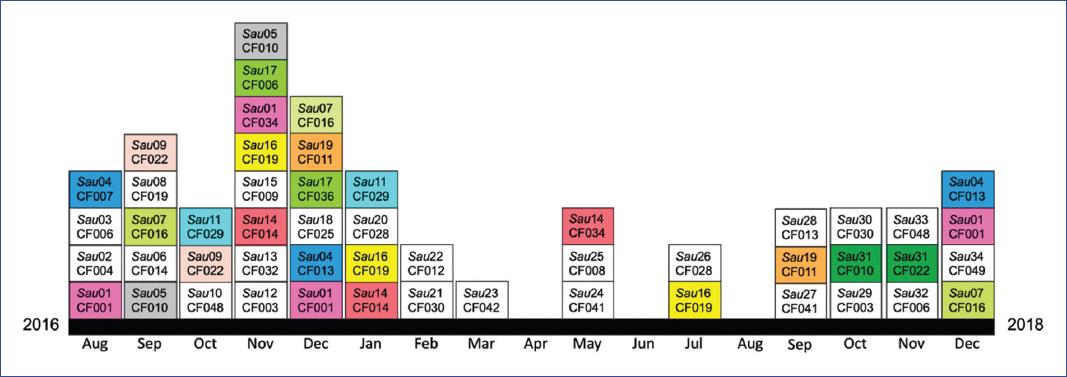
Of the 50 pediatric patients studied, the most frequently isolated bacterial pathogen was P. aeruginosa, followed by S. aureus4. S. aureus was isolated from 26/50 patients, from whom we obtained 51 isolates (Figure 1). Chromosomal analysis by PFGE yielded 34 patterns (pulsotypes) (Table 1). With the 34 patterns, we were able to identify nine clusters (designed as I-IX), which differed by approximately 85% in PFGE band similarity (Figure 1). The cluster with more members (thirteen) was cluster II. The results showed that nine patients (CF001, CF010, CF011, CF013, CF014, CF016, CF019, CF022, and CF029) were chronically infected (the same pulsotype was identified in two or more samples for two or more months) (Figure 2). Twelve patients were colonized with different pulsotypes: eleven with two different pulsotypes and one with three different pulsotypes (Table 1). Patients CF008 (colonized with Sau25) and CF014 (colonized with Sau06 and Sau14) died during the study period.
Table 1 S. aureus pulsotypes isolated from Mexican children with cystic fibrosis
| Patient | Sample | |||
|---|---|---|---|---|
| 1st | 2nd | 3rd | 4th | |
| CF001 | Sau01 | Sau01 | Sau01 | — |
| CF003 | x | Sau12 | Sau29 | — |
| CF004 | x | Sau02 | — | — |
| CF006 | Sau03 | Sau17 | Sau32 | — |
| CF007 | Sau04 | x | — | — |
| CF008a | x | Sau25 | — | — |
| CF009 | x | Sau15 | x | x |
| CF010 | Sau05 | Sau05 | Sau31 | — |
| CF011 | x | Sau19 | Sau19 | — |
| CF012 | x | x | Sau22 | — |
| CF013 | x | Sau04 | Sau04 | Sau28 |
| CF014a | Sau06 | Sau14 | Sau14 | — |
| CF016 | Sau07 | Sau07 | Sau07 | — |
| CF019 | Sau08 | Sau16 | Sau16 | Sau16 |
| CF022 | Sau09 | Sau09 | Sau31 | — |
| CF025 | x | Sau18 | — | — |
| CF028 | x | Sau20 | Sau26 | — |
| CF029 | Sau11 | Sau11 | — | — |
| CF030 | x | Sau21 | Sau30 | — |
| CF032 | Sau13 | — | — | — |
| CF034 | Sau01 | Sau14 | — | — |
| CF036 | Sau17 | — | — | — |
| CF041 | x | Sau24 | Sau27 | — |
| CF042 | Sau23 | — | — | — |
| CF048 | Sau10 | Sau33 | — | — |
| CF049 | x | Sau34 | — | — |
aPatient died.
X, sample with no S. aureus isolate; —, no sample available.

Figure 1 Dendrogram generated from PFGE analysis of 51 S. aureus isolates from pediatric CF patients. A representative PFGE profile of each pulsotype was used to construct the dendrogram. Each pulsotype frequency (number of isolates) is indicated, and each row indicates the patient in which the pulsotype was identified. Patients colonized by more than one pulsotype are indicated (*). A superscript indicates the frequency of each pulsotype identified per patient. The dotted line indicating 85% similarity was used to determine the cluster designation (I-IX).
Antimicrobial susceptibility of S. aureus pulsotypes isolated from CF children
One of the 34 S. aureus pulsotypes identified was used to determine the antimicrobial susceptibility pattern to ten different classes of antibiotics: glycylcyclines, oxazolidinones, glycopeptides, folate pathway antagonists, fluoroquinolones, tetracyclines, macrolides, ansamycins, penicillins, lincosamides, and aminoglycosides (Table 2). We detected 3/34 (8.8%) MDR pulsotypes, and a low rate of resistance to vancomycin (8.8%), moderate resistance to tetracycline and erythromycin (20.6%), and clindamycin (17.7%). All the pulsotypes were susceptible to tigecycline, linezolid, and trimethoprim/sulfamethoxazole (Table 2). We found only two MRSA pulsotypes (5.9%): Sau08 and Sau16, both isolated from patient CF019. Sau16 was isolated three times at different times during sample collection (Figure 1 and Table 1). We identified both pulsotypes (Sau08 and Sau16) carrying the mecA gene by PCR (data not shown). These results also revealed that the two pulsotypes were MRSA with an MDR phenotype.
Table 2 Antibiotic susceptibility for 34 Staphylococcus aureus pulsotypes of pediatric patients with cystic fibrosis
| Antibiotic family | Antibiotic | Breakpoints (µg/mL) | Susceptible n (%) | Intermediate n (%) | Resistant n (%) | ||
|---|---|---|---|---|---|---|---|
| S | I | R | |||||
| Aminoglycosides
Lincosamides |
Gentamicin | ≤ 4 | 8 | ≥ 16 | 30 (88.2) | 2 (5.9) | 2 (5.9) |
| Clindamycin | ≤ 0.5 | 1-2 | ≥ 4 | 27 (79.4) | 1 (2.9) | 6 (17.7) | |
| Penicillins Ansamycins |
Oxacillin (MRSA) | ≤ 2 | — | ≥ 4 | 30 (88.2) | 0 (0.0) | 4 (11.8) |
| Rifampicin | ≤ 1 | 2 | ≥ 4 | 31 (91.2) | 0 (0.0) | 3 (8.8) | |
| Macrolides | Erythromycin | ≤ 0.5 | 1-4 | ≥ 8 | 27 (79.4) | 0 (0.0) | 7 (20.6) |
| Tetracyclines | Tetracycline | ≤ 4 | 8 | ≥ 16 | 27 (79.4) | 0 (0.0) | 7 (20.6) |
| Fluoroquinolones | Ciprofloxacin | ≤ 1 | 2 | ≥ 4 | 29 (85.3) | 2 (5.9) | 3 (8.8) |
| Levofloxacin | ≤ 1 | 2 | ≥ 4 | 31 (91.2) | 2 (5.9) | 1 (2.9) | |
| Moxifloxacin | ≤ 0.5 | 1 | ≥ 2 | 33 (97.1) | 0 (0.0) | 1 (2.9) | |
| Folate pathway inhibitors | Trimethoprim/sulfamethoxazole | ≤ 2/38 | — | ≥ 4/76 | 34 (100.0) | 0 (0.0) | 0 (0.0) |
| Glycopeptides Oxazolidones |
Vancomycin | ≤ 2 | 4-8 | ≥ 16 | 31 (91.2) | 0 (0.0) | 3 (8.8) |
| Linezolid | ≤ 4 | — | ≥ 8 | 34 (100.0) | 0 (0.0) | 0 (0.0) | |
| Glycylcyclines | Tigecycline | ≤ 0.25 | — | — | 34 (100.0) | 0 (0.0) | 0 (0.0) |
MRSA, methicillin-resistant Staphylococcus aureus S, susceptible; I, intermediate; R, resistant.
*Breakpoints were obtained from the Clinical and Laboratory Standards Institute (2019).
Pulsotypes of S. aureus induced cytotoxicity in human THP1-derived macrophages
The ability of S. aureus to induce cytotoxicity in human THP1 monocytes differentiated into macrophages (THP1m) was evaluated. The results showed that 30/34 (88.2%) pulsotypes were cytotoxic (> 10% of cytotoxicity) (Figure 3), 8/34 (23.5%) were highly cytotoxic (> 50% of cytotoxicity), and 14/34 (41.2%) were able to disrupt the THP1m monolayers (data not shown). To determine whether the cytotoxic effect was associated with pulsotypes carrying the PVL gene, we amplified the lukPV gene by PCR. The results showed that none of the analyzed pulsotypes carried the lukPV gene (data not shown).

Figure 3 Induction of cytotoxicity by S. aureus isolates in THP1macrophages: 5 x 105 THP1 monocytes were differentiated into macrophages with 100 ng/mL of PMA for 24 h. Cells were infected with S. aureus isolates at MOI of 100 for 30 min. Once infected, cells were washed and incubated for 24 h. Supernatants were used to quantify macrophage cell death (cytotoxicity). Results were obtained from three independent experiments, each in duplicate (n = 6). Data were plotted as the mean ± SD and analyzed by one-way ANOVA and Dunnett’s multiple comparisons in relation to S. aureus -USA300. **p < 0.01; ***p < 0.001; ****p< 0.0001; ns: non-significant.
Discussion
The lower airways of children with CF are rapidly colonized by S. aureus and non-typeable H. influenzae5. However, the pulmonary bacterial microbiome gradually changes with the emergence and persistence of P. aeruginosa during adolescence and adulthood5. The eradication of S. aureus in the lower respiratory tract has been compromised by the emergence of MRSA20. The World Health Organization (WHO) has declared priority level 2 in identifying new antibiotics to combat MRSA. To date, MRSA is usually associated with the community- and hospital-acquired infections. In CF, the occurrence of MRSA has been associated with a more rapid decline in lung function21. In a previous study, 44% of children with CF (22/50) were colonized by P. aeruginosa and S. aureus4. In the present study, we identified 34 unrelated pulsotypes, of which 2/34 were MRSA. In particular, the Sau16 pulsotype was consistently isolated three times in patient CF019 during one year. We also isolated the genetically related Sau08 pulsotype (group V) in this patient, suggesting a genetic evolution. Analysis of clinical isolates of S. aureus by whole-genome sequencing suggests possible genetic evolution and spread22. In this study, we identified that pulsotypes Sau08 and Sau16 are MRSA with a consistent MDR phenotype. We also identified the persistence of S. aureus in eight patients. Persistence and long-term carriage of S. aureus are often associated with specific phenotypes, including small colony variants, increased antimicrobial resistance, and biofilm-formation23. Furthermore, MRSA colonization is more frequently associated with individuals carrying the DF508 mutation24. The patient CF019 from whom we isolated both MRSA pulsotypes carried the DF508 mutation4.
S. aureus contains several virulence factors that promote host tissue damage25-27. Toxic shock syndrome toxin (TSST-1) and PVL are two important secreted virulence factors12. Our results showed that none of the S. aureus pulsotypes tested carried the lukPV gene. It has been demonstrated that PVL expression is not sufficient to induce cell death28, suggesting the presence of additional virulence factors that contribute to pathogenesis27.
We determined that 88.2% of the isolates induced cell death in THP1-derived macrophages and that 14/34 (41.2%) of the tested pulsotypes could alter the integrity of the THP1m monolayers. S. aureus α-toxin is a secreted virulence factor involved in the disruption of epithelial cell monolayers29, suggesting that this virulence factor could alter THP1m monolayers, a hypothesis that we will address in the future.
In conclusion, in this study, we demonstrated a low presence of MRSA and MDR and a high frequency of S. aureus pulsotypes with the ability to induce cytotoxicity in THP1-derived macrophages.

