











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
Burkitt's lymphoma (BL) is a monoclonal B cell non-Hodgkin's lymphoma with a high proliferative index1. Endemic BL is extranodal and tumors are frequently found in the jaw or abdominal region2. The peak age of onset is 6 years3 indicating that there is a very short time frame between exposure to cancer promoting events and the emergence of malignancy. There are three forms of BL found worldwide: endemic, sporadic, and AIDS-associated and all carry a t8;14 chromosomal translocation resulting in the deregulation of the c-myc oncogene4 5. It is likely however, that different pathogenic mechanisms drive the emergence of these BL subtypes6 7. The focus of this review is on the endemic form of BL (herein referred to simply as BL).
Both molecular and epidemiologic studies have indicated that there is an etiologic link between EBV and the endemic form of BL8 9 10 11. At the molecular level, the viral genome is present in all cells12, exists as a clonal population within the tumors9 and the viral protein EBNA-113 along with viral non-coding BART microRNAs14 15 are consistently expressed within the tumors. In addition, the capacity of EBV to transform B cells also highlights the virus' oncogenic potential. A large-scale prospective study conducted in Uganda in the 1970s provided evidence that EBV infection was a risk factor for BL. In this study, greater than 40,000 children were pre-bled, serum was stored and when tumors appeared, very high antibody titers against the EBV viral capsid antigen (VCA) were found in children who subsequently developed BL. The elevated VCA titers, and the stability of the elevated VCA antibodies over time, led de-Thé et al.16 to suggest that infection of infants with EBV early in life could result in an infection that was poorly controlled by the host and thus increased the risk for BL. Plasmodium falciparum, is also connected to BL both based on the overlapping regions of high malaria transmission with areas of high BL incidence17 18 19 20 as well as case control studies21 22.
2. Epstein-Barr virus transmission
EBV is an enveloped gamma herpes virus that is transmitted primarily through contact with saliva23. In developed countries, there is a biomodal distribution of the age of EBV infection with 30-50% of children infected before 5 years of age and then a later transmission occurring in young adulthood. For example, in a recent study by Condon et al., they evaluated the seroprevalence of EBV infection in a cross-sectional study of children in the U.S. aged 18 months-20 years of age. They found that 31% of the children were EBV seropositive by 5 years of age, whereas 71% were seropositive by 19 years of age24. Infection in childhood is thought to be primarily asymptomatic while the later age of infection can cause infectious mononucleosis (IM), a self-limiting disease25. IM is characterized by expansion of both EBV-specific and non-specific CD8+ T cells. In a study of Gambian infants infected with EBV by 14 months of age, although there was EBV-specific CD8 T cells detected, there was no concurrent over expansion of the CD8+ T cell pool26 perhaps explaining why EBV infection in infants is asymptomatic.
There are a limited number of published longitudinal studies on primary EBV infection in infants27 28 29 30 31 32 and most have utilized only serologic markers as indicators of infection. In a more recent study following an infant cohort born in rural areas in Kenya, significantly higher levels of EBV infection at less than 1 year of age were observed as compared to other studies31. Of note, in the malaria high transmission area, 35% of infants were infected with EBV by 6 months of age suggesting that malaria infection modulated the age of EBV infection31. Early age of EBV infection was also observed in Kenyan infants born to HIV infected mothers32. Both of these studies found that early age of acquisition of EBV was associated with a poor control of EBV infection as indicated by high viral loads that were maintained over time. The neonatal immune system is not as effective as the adult immune system for reasons that include lack of immunologic memory, immaturity and skewing towards a Th2 phenotype33 34 35. Early transmission of EBV could induce tolerance of the viral antigen and consequently limit the specific immune response, as seen for early age of hepatitis B virus infection36. Alternatively, infants lack a fully functional cytotoxic T cell response before 12 months of age34, so infection early in life could lead to ineffective or minimal control of the virus. Early age of infection with subsequent high viral loads could increase the risk for subsequent EBV-associated malignancies.
An alternative source of EBV transmission has been hypothesized to be breast milk37 38 39 but in most studies to date, only viral DNA was measured. More recently, infectious virus, not just viral DNA, was found in breast milk providing support to the hypothesis that breast milk could be a source for EBV transmission40. Pregnant women with malaria have high viral loads41, and the loss of control of EBV latency following P. falciparum infection during pregnancy and subsequent increase in EBV load in circulation possibly contribute to enhanced shedding of EBV in maternal breast milk post-partum40 and drives early age of transmission of EBV. EBV DNA was detected in breast milk of HIV infected mothers38 39 and this could contribute to early age of EBV infection in infants born to HIV infected women. Why there is higher prevalence of EBV in breast milk of mothers in developing40 as compared to developed countries42 could also reflect the higher burden of EBV in the general population.
3. EBV persistence
A model for EBV persistence proposed by Thorley-Lawson, the Germinal Center (GC) Model, is an elegant model based on years of study of healthy U.S. adults43. In this model, EBV exploits the differentiation of B cells in the lymphoid tissue to set up a lifelong persistent latent infection in memory B cells in equilibrium with the host. In healthy, EBV-seropositive adults, there is a stable, low frequency of latently infected cells estimated at 1-200 EBV positive B cells in 105 total B cells44. Without enrichment for B cells from peripheral blood, it is difficult to detect EBV infected cells in healthy EBV seropositive adults by PCR amplification using real-time quantitative PCR45 46 or quantitative competitive PCR47. However, unaccounted for in this model is how other systemic infections can influence EBV persistence.
3.1. Malaria and EBV persistence
In the last decade, several studies have pointed to a profound dysregulation of EBV persistence and immunity in children due to malaria31 46 48 49 50 51 52 53 54 55 56 57. Repeated malaria infections during infancy expand the viral load54 and an expansion of EBV infected cells during acute malaria has also been found49 58. The cysteine rich interdomain 1α (CIDR1α) of the P. falciparum erythrocyte membrane protein, which is expressed in infected red blood cells, binds to non-immune immunoglobulins, and acts as a T-cell independent B-cell activator59 and induces EBV reactivation from B cells60. EBV is more frequently reactivated in children living in a malaria endemic region48 and more directly, elevated viral DNA in plasma is found during acute malaria51 52. A decline in EBV DNA in the plasma following anti-malaria treatment53 is also indicative of viral reactivation driven by Plasmodium infection.
In children living in a malaria endemic region, EBV viral loads are more readily detected as compared to western controls46 49 suggestive of a long-term consequence of repeated malaria infections. More recently, in regions of the Gambia where malaria transmission has been reduced, the dysregulation of EBV immunity by malaria appears to have also been minimized61.
4. Activation induced cytidine deaminase (AID) and BL
While it is likely that elevated EBV load increases the risk for BL, the mechanistic link is unknown. In transplant patients, EBV viral load is monitored closely and elevated viral load is associated with increased risk for post-transplant lymphoproliferative disease62 yet these patients do not go on to develop BL. So, elevated viral load alone is not sufficient for the emergence of a malignant clone. Recent clues point to a model where chronic antigenic activation of EBV-infected B cells within the context of repeated P. falciparum infections may lead to cytogenetic abnormalities induced by the enzyme activation induced cytidine deaminase (AID)63 64 65.
AID is required for class switch recombination and somatic hypermutation in germinal center B cells66. But AID over-expression in AID deficient B cells was sufficient to induce IgH-c-myc translocations67 characteristic of BL. AID also induces lesions on the c-myc gene68. In addition, when Eμ-c-myc transgenic mice were crossed with AID deficient mice, only AID+/+ mice developed predominantly mature B cell lymphomas69. The strongest evidence for a role of aberrant AID activation by Plasmodium infection comes from a recent study by Robbiani et al.70 In this study, the authors found that repeated infections of p53-deficient mice with Plasmodium chabaudi resulted in development of B-cell lymphomas that had the characteristic c-myc translocation. Moreover, development of the lymphomas was dependent on AID. Although these studies were done in mouse models, these data argue for a critical role of AID in the c-myc:IgH translocation characteristic of BL and suggest that aberrant AID expression could be a risk factor for lymphomagenesis. In support of this hypothesis, a strong correlation between AID expression in peripheral blood lymphocytes and increased risk for non-Hodgkin's lymphoma was observed71 72 and children who were EBV viral load positive from a malaria endemic region were found to have elevated AID expression63. Cell culture studies showed that P. falciparum could induce AID activation and class switch recombination in B cells64 73. If overexpression of c-myc occurred following an AID-mediated translocation, normal B cells would die by apoptosis. However, EBV latent proteins are anti-apoptotic and could thus allow a B cell to tolerate the c-myc translocation74.
5. BL and EBV association
Although EBV can be readily detected in the endemic pediatric form of BL found in sub-Saharan Africa, there is a variable association of EBV with the sporadic form of BL. Sporadic BL that occurs in adults in developed countries rarely has EBV detected in the tumor. However, in South America, there is an intermediate level of EBV detected in sporadic BL tumors75. A larger series of cases of BL from Brazil found a predominance of EBV detection in the pediatric form of BL as compared to adults and a higher prevalence of pediatric EBV+ BL in the northern part of Brazil where there is also a higher prevalence of malaria and other infectious diseases76. The intermediate incidence of EBV association with sporadic BL in South America raises the question of whether malaria infection in other parts of the world outside sub-Saharan Africa could also drive lymphomagenesis. A key difference of malaria transmission in Africa is the perennial and intense nature vs. the epidemic and seasonal transmission found in South and Central America. However, it is possible that there could exist a concurrence of early age of EBV infection and malaria infection that could increase the risk for lymphomagenesis in other parts of the world. Studies on the age of EBV infection in Latin America remain few. A recent report from a pediatric hospital case series in Mexico found symptomatic EBV infection occurring in children characteristic of infectious mononucleosis with a mean age of 5 years,77 much younger than seen in U.S.-based series23. However, a limitation of the Mexico study is that it was based on hospitalized cases rather than a population-based study so true incidence cannot be determined. Nonetheless, this study does suggest that EBV infection is occurring in very young children in Mexico.
Summary
Cumulative evidence suggests that EBV persistence is in dysequilibrium when primary infection occurs in very young infants and co-infection with P. falciparum malaria is common. The prevailing model for EBV persistence, the Germinal center model, was based on studies of healthy U.S. adults and as such does not take into account the disruption to B cell homeostasis78 79 and germinal center architecture that occurs during acute P. falciparum malaria infection80 or the ability of Plasmodium to directly interact with B cells through either TLR9 ligation81 or binding of the P. falciparum-infected red blood cell to B cells60. This dysequilibrium of EBV persistence results in high numbers of latently infected cells, which are at risk for AID activation induced by P. falciparum and subsequent c-myc translocation (Fig. 1). Not discussed in this review is a potential role for EBV to suppress apoptotic pathways in latently infected cells,82 which would allow B cells with a c-myc translocation to emerge. It remains to be seen whether other pathogens can also lead to dysequilibrium of EBV infection and higher viral loads and account for some of the diverse burden of EBV-associated malignancies worldwide.
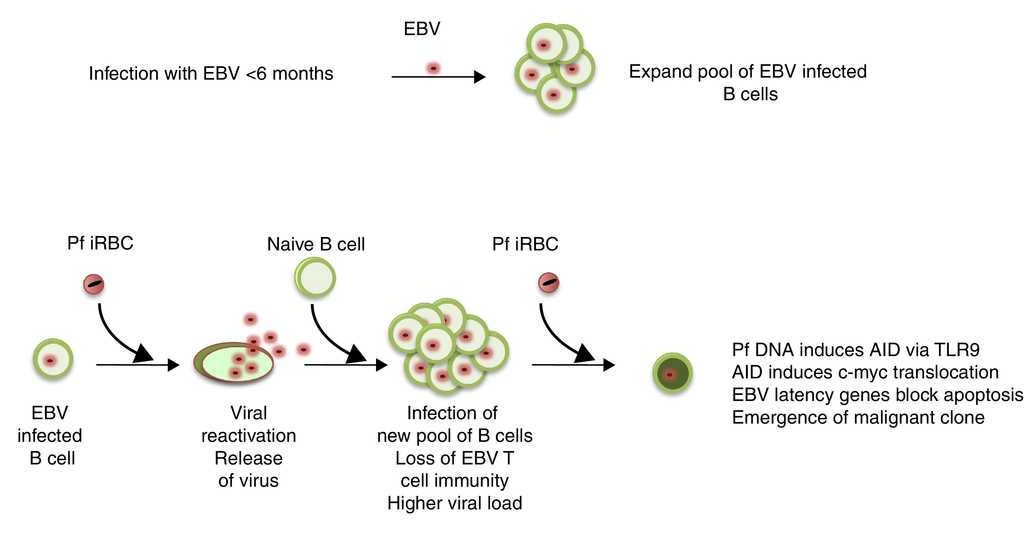
Figure 1 Model for interaction between EBV and P. falciparum malaria. P. falciparum (Pf) malaria results in EBV transmission in infants <6 months of age. These infants have poor control of EBV infection and a higher number of latently infected cells. Repeated infection with P. falciparum throughout infancy and early childhood expands the number of infected cells by inducing EBV reactivation and reinfection of new pool of B cells as well as suppression of EBV T cell immunity. P. falciparum can also act on the EBV infected B cells to induce AID via TLR9 ligation and subsequent c-myc translocation. Because EBV induces epigenetic changes in the infected B cells, the c-myc translocation is tolerated and there is emergence of a malignant B cells.

