











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El sistema CRISPR (Clustered Regularly Interspersed Short Palindromic Repeats), así como su proteína asociada Cas (CRISPR associated) se encuentran ampliamente distribuidos en el mundo microbiano, de hecho, están presentes en el 40% de las especies bacterianas y en el 90% de las arqueas reportadas (Mojica et al., 2000). La importancia biológica de este sistema radica en que se constituye como un sistema inmune adaptativo microbiano que responde a elementos de DNA invasores, como pueden ser plásmidos o virus. Recientemente se logró adaptar este sistema como una herramienta para la edición genética y su uso se está extendiendo rápidamente debido a su simplicidad y gran precisión (Doudna & Charpentier, 2014).
Desde el descubrimiento de la doble hélice del DNA por James Watson y Francis Crick en 1953, los investigadores han contemplado la posibilidad de realizar modificaciones sitio específicas en esta molécula, primero con la intención de investigar la función de cada gen y posteriormente con el propósito de revertir o prevenir enfermedades producidas por mutaciones puntuales en el genoma, como la distrofia muscular o la fibrosis quística. Sin embargo, introducir mutaciones sitio específicas en el genoma de células y organismos ha sido una tarea sumamente complicada. Las estrategias más exitosas se basan en el empleo de las nucleasas de dedos de zinc (ZFN) y las nucleasas tipo activadores de transcripción (TALEN´s) que utilizan los principios de reconocimiento del DNA-proteína para introducir cortes específicos en la molécula de DNA (Kim & Kim., 2014). No obstante, dificultades con el diseño y síntesis de las proteínas específicas necesarias para que ambos sistemas puedan funcionar han supuesto barreras infranqueables en la adopción generalizada de estos dos métodos de edición genética.
Actualmente, el sistema CRISPR/Cas ha sido empleado para generar diversas mutaciones tanto en células de mamíferos y de plantas, así como de microorganismos, con la finalidad de realizar estudios genéticos (Charpentier & Marraffini, 2014), demostrando ser una herramienta de edición genética más eficiente que sus predecesores (los sistemas ZFN y TALEN´s).
Origen del sistema CRISPR/Cas
El sistema CRISPR/Cas fue descrito por primera vez como una serie de secuencias directas repetidas cortas, e inter-espaciadas por secuencias cortas en el genoma de la bacteria Escherichia coli (Ishino et al., 1987). Debido a que las secuencias espaciadoras son altamente diversas inclusive entre cepas estrechamente relacionadas, inicialmente se utilizaron con motivos de tipificación, es decir, para diferenciar distintos aislados, cepas y especies bacterianas (Groenen et al., 1993). Posteriormente, se encontró que el elemento asociado Cas contiene dos dominios clave para su actividad, un dominio de nucleasa (RuvC) y otro de helicasa (HNH) de los cuales se hablará más adelante (Jansen et al., 2002). Una observación clave para descifrar la función biológica del sistema CRISPR/Cas fue realizada por diversos grupos de investigación, lo cuales reportaron que muchas secuencias espaciadoras eran similares a elementos de DNA exógeno, como plásmidos y virus y por lo tanto podrían ser derivados de los mismos (Bolotin et al., 2005; Mojica et al., 2005; Pourcel et al., 2005).
Se propuso que el sistema CRISPR/Cas actúa como un sistema inmune adaptativo, que emplea el RNA antisentido codificado por las secuencias espaciadoras como guía para que la nucleasa Cas reconozca como blanco a las moléculas de DNA exógeno y lo desactive mediante la introducción de cortes específicos (Makarova et al., 2006). Posteriormente, se describió que en E. coli y Staphylococcus epidermidis los RNA codificados en el locus del sistema CRISPR (crRNA) forman un complejo con la proteína Cas y que dicho complejo es capaz de interferir con la proliferación de fagos (Brouns et al., 2008; Marraffini & Sontheimer, 2008). Adicionalmente, se estableció que el sistema CRISPR/Cas de Streptococcus thermophilus opera como un sistema inmune adaptativo de defensa contra la invasión de fagos líticos (Barrangou et al., 2007).
Actualmente, el sistema CRISPR/Cas ha cobrado una gran relevancia en diversos campos de investigación, como la medicina y la biotecnología, ya que gracias a la capacidad del sistema para introducir cortes específicos en la molécula de DNA ha sido posible adaptarlo como una poderosa herramienta para la edición genética. A continuación se describen los componentes del sistema y su mecanismo de acción como determinante en la inmunidad bacteriana y como herramienta para la manipulación genética.
Componentes y mecanismo del sistema CRISPR/Cas
Hasta la fecha se han identificado seis tipos distintos de sistemas CRISPR/Cas (I-VI) basados en el mecanismo molecular que emplean para el reconocimiento del DNA y la forma en que se unen al mismo (Makarova et al., 2011). Sin embargo, aquí nos enfocaremos únicamente en describir al sistema tipo II, debido a que es el más utilizado para realizar ediciones genéticas (Jiang & Marraffini, 2015).
El locus donde se encuentran codificados los elementos del sistema CRISPR/Cas consta de una región promotora, que se encarga de modular la transcripción de todos los elementos. Posteriormente se encuentra la región CRISPR, que se compone de los elementos repetidos y de los espaciadores, estos últimos codifican moléculas de RNA cortas (30-40 pares de bases) a las cuales se les denomina RNA CRISPR (crRNA) y son las encargadas de guiar a la proteína Cas a su sitio blanco. Finalmente se encuentran los genes que codifican a las proteínas Cas (Figura 1) (Jiang et al., 2013).
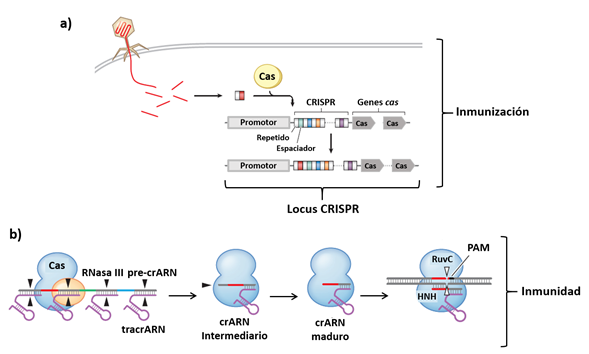
Figura 1 Fases del sistema inmune adaptativo bacteriano CRISPR/Cas. (a) Fase de inmunización. Fragmentos de material genético exógeno adquirido a través de virus o plásmidos es incorporado en el locus CRISPR, donde se utilizará posteriormente para proteger a la bacteria de una reinfección. (b) Fase de inmunidad. La secuencia espaciadora es transcrita desde la región promotora y el transcrito resultante, crRNA intermediario, es procesado en un crRNA maduro. El crRNA es utilizado para guiar a la nucleasa específica Cas, que cuenta con los dominios RuvC y HNH, hasta su sitio blanco. Las puntas de flechas, negras y blancas, indican los sitios de unión del RNA y del DNA respectivamente. Además se muestra el sitio PAM, necesario para el reconocimiento del sitio blanco. Modificado de Jiang & Marraffini, 2015. Ver la versión a color en línea.
El mecanismo mediante el cual el sistema CRISPR/Cas codifica un sistema inmune adaptativo preciso contra elementos de DNA invasores consta de dos fases: I) Fase de inmunización: después de la incorporación del DNA exógeno proveniente de virus o de plásmidos, el elemento Cas reconoce a la molécula extraña e integra un fragmento en el locus CRISPR donde se convertirá en un nuevo elemento espaciador (Figura 1) (Horvath & Barrangou, 2010). II) Fase de inmunidad: el elemento espaciador incorporado se transcribe como un crRNA, que posteriormente sirve de guía para que la proteína Cas alcance su blanco, que es el elemento genético invasor (Figura 1) (Horvath & Barrangou, 2010). Los dominios conservados de la proteína Cas son cruciales para su actividad, el dominio de helicasa (HNH) se encarga de abrir la doble hebra del DNA, y el dominio de nucleasa (RuvC) le confiere la capacidad para cortar el DNA en la región definida por los crRNA (Sapranauskas et al., 2011).
Para el proceso de inmunidad es necesario un sitio adicional de reconocimiento compuesto por una secuencia corta de DNA (de 3 a 5 pares de bases, que se encuentra inmediatamente río arriba de la secuencia blanco y que reconocen tanto a los crRNA como a la proteína Cas (Sashital et al., 2012). Este sitio se conoce como motivo adyacente al protoespaciador (PAM) y se cree que es necesario para evitar la autoinmunidad, ya que sin este elemento de reconocimiento adicional, los crRNA reconocerían su propio gen codificante y la nucleasa podría introducir cortes en su propio genoma (Wang et al., 2015A). Además, las secuencias del elemento PAM difieren entre cada organismo, por ejemplo, en Streptococcus pyogenes SF370 la secuencia es NGG (Mojica et al., 2009), por lo que estas secuencias se empleaban para la tipificación bacteriana.
Como se ha descrito, el mecanismo por el que el sistema CRISPR/Cas confiere inmunidad parece bastante simple; sin embargo, para que los crRNAs estén listos para cumplir con su función es necesario un proceso previo de maduración, donde se forma una molécula híbrida (RNA-proteína) entre el crRNA y la proteína Cas. Primero se produce un pequeño crRNA que cuenta con una región complementaria a la secuencia repetida del locus CRISPR, que se denomina crRNA transcodificado (tracrRNA). La proteína Cas se une al tracrRNA y este complejo se alinea con las secuencias repetidas que son precursoras de los crRNA. Este RNA de doble hebra (dsRNA) va a ser cortado por la enzima RNasa III y se genera una nueva molécula que contiene a la proteína Cas cargada con los tracrRNA y el crRNA guía (Figura 1) (Jiang & Marraffini, 2015).
Los elementos del sistema CRISPR se expresan de manera constitutiva, por lo que siempre están listos para el momento en el que ocurra una infección. Cuando la célula entra en contacto con una molécula de DNA invasora, el complejo tracrRNA/crRNA/Cas escanea a la molécula exógena en busca de los elementos de reconocimiento. Una vez localizada una secuencia de unión, el elemento Cas corta a la molécula de DNA y de esta manera desactiva a la potencial amenaza (Gasiunas et al., 2012).
Mecanismo de edición genética basado en el sistema CRISPR/Cas
El primer paso para convertir el sistema inmune bacteriano CRISPR/Cas en una tecnología eficaz para la edición de genes fue convertir a los tracRNA y crRNA en un único RNA denominado sgRNA (Jinek et al., 2012), de esta manera se puede evitar el paso de maduración y por lo tanto no es necesaria la acción de la RNasa III. El sgRNA conserva dos características críticas para su función: la región de reconocimiento del extremo 5´que es la responsable de la unión específica con el sitio blanco y el sitio de unión del elemento Cas en el extremo 3´ (Jinek et al., 2012). La implementación de un único sgRNA creó un sistema simple y eficaz que necesita únicamente dos componentes (sgRNA y Cas) para cumplir con su acción. Además, la secuencia guía del sgRNA se puede modificar para programar el sitio de corte de manera específica, por lo que potencialmente es posible modificar cualquier molécula de DNA (Doudna & Charpentier, 2014).
La introducción de cortes precisos en la molécula de DNA, aunque es un evento crucial, es sólo en el primer paso para realizar una edición genética exitosa, debido a que dependiendo del tipo de modificación que se desee realizar es necesario, por ejemplo, introducir nuevas pares de bases, o eliminar algunas otras (mutación por inserción o por deleción respectivamente). Una vez que el complejo sgRNA-Cas9 del sistema CRISPR/Cas ha introducido el corte en la doble hélice de DNA y si se requiere reparar el daño por ejemplo en células eucariotas existen dos sistemas de reparación de daño, que serán los encargados de insertar las modificaciones y son: la recombinación no homóloga (RNH) y la recombinación homóloga (RH), por otro lado, muchas bacterias, incluyendo a E. coli, únicamente cuentan con el sistema RH (Shuman & Glickman., 2007). Para completar el proceso de edición genética, durante la reparación con el sistema RNH se insertan o se escinden regiones de varias pares de bases cercanos al sitio del corte, este evento altera el marco de lectura abierto del probable gen blanco y por lo tanto lo desactiva (mutación knockout) (Figura 2) (Cong et al., 2013; DiCarlo et al., 2013; Li et al., 2013). Por otro lado, el sistema RH necesita de una molécula de DNA adicional que actúe como templado para la reparación del corte, estos templados pueden estar presentes en el cromosoma (genes duplicados) o pueden ser suministrados de manera exógena; por ejemplo, a través de plásmidos. Gracias a que el templado adicional, necesario para el sistema RH, se puede suministrar de manera exógena, esta vía ha sido empleada para introducir secuencias específicas dentro de las regiones cortadas con el sistema CRISPR/Cas (Figura 2). Hasta el momento, el sistema RH ha sido empleado de manera eficiente para realizar modificaciones específicas, como inserciones, deleciones (Gratz et al., 2013) y sustituciones (Shalem et al., 2015; Wang et al., 2013).
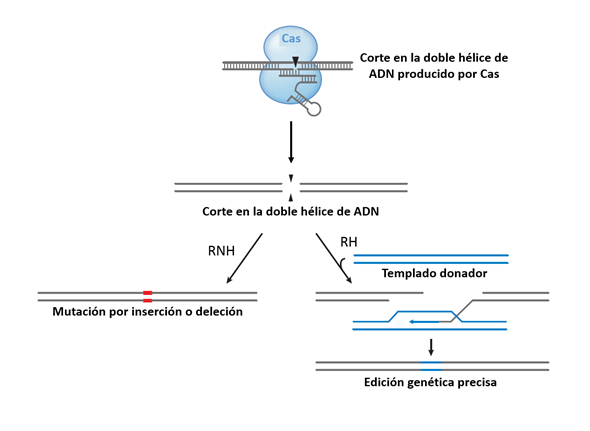
Figura 2 Edición genética mediada por el sistema CRISPR/Cas. La nucleasa Cas introduce cortes precisos en la doble hélice del DNA. Las lesiones genómicas pueden repararse a través de dos vías: El sistemas de reparación homólogo (RH) utiliza un templado adicional como donador para la recombinación, reemplazando la secuencia existente con la secuencia modificada de interés. La vía de reparación no homóloga (RNH) une los extremos cortados en un proceso donde pueden insertarse o removerse secuencias de DNA. Las puntas de flecha indican los sitios de corte del DNA. Modificado de Jiang & Marraffini, 2015. Ver la versión a color en línea.
En contraste con los sistemas de edición que han sido empleados a la fecha como el sistema ZFN y TALEN´s, que requieren una cantidad significativa de elementos para modificar el DNA (Kim & Kim., 2014), en el sistema CRISPR/Cas únicamente se requiere modificar los RNA guías para desactivar un gen y una molécula de DNA adicional, que sirva como templado, para introducir modificaciones específicas (Doudna & Charpentier, 2014). Por esta razón, el sistema CRISPR/Cas ha sido adoptado rápida y ampliamente por la comunidad científica para editar genomas en un gran número de células y de organismos como el caso del maíz donde fue mutado en el gen Wx1, generando una mutante que presenta un fenotipo con alto contenido de amilopectina (Waltz, 2016) o en cerdos la mutación del gen MSTN, que codifica un regulador negativo del crecimiento muscular (Wang et al., 2015B). Aunque el uso del sistema CRISPR/Cas se está extendiendo rápida y ampliamente, esta revisión se centrará en describir una aplicación novedosa, como es el uso del sistema en el desarrollo de un antimicrobiano secuencia específica.
Uso del sistema CRISPR/Cas como antimicrobiano secuencia específica
La resistencia a los antimicrobianos representa uno de los más grandes retos de nuestro tiempo debido a que el uso indiscriminado de estos agentes ha permitido la selección de bacterias multirresistentes por lo que las opciones para un tratamiento efectivo han disminuido y en algunos casos se han agotado (Spizek & Havlicek, 2015). Aunado a esto, en la actualidad hay escases en el desarrollo de nuevos antimicrobianos, ya que desde hace varias décadas no ha salido al mercado una nueva familia de agentes antimicrobianos (Aminov, 2017). Predicciones recientes estiman que de continuar la misma tendencia, para el año 2050 el número de muertes producidas por bacterias multirresistentes a los antimicrobianos alcanzarán los 10 millones anuales, cifra que superaría a las muertes producidas por cáncer que rondarían los 8.2 millones anuales (O´neill, 2016). Por lo anterior se han buscado nuevas estrategias para combatir a las infecciones bacterianas y una adaptación en el uso del sistema CRISRP/Cas podría ofrecer una nueva opción eficiente en el tratamiento de infecciones bacterianas.
El desarrollo de una nueva estrategia basada en el uso del sistema CRISPR/Cas surgió de la observación de que los cortes introducidos en el cromosoma por el elemento Cas resultan letales para algunas bacterias cuando no se les suministra un templado exógeno para realizar la reparación con el sistema RH, ya que no cuentan con el sistema RNH, por lo que es posible programar a los sgRNA del sistema para introducir cortes de manera selectiva sobre bacterias de interés (Figura 3) (Citorik et al., 2014). Además, a diferencia de los antimicrobianos convencionales cuya actividad se considera de amplio espectro, es decir, que son tóxicos para una gran cantidad de bacterias, el sistema CRISPR/Cas puede programarse de manera específica para que únicamente actúe sobre un tipo de población bacteriana, quedando intactas otras poblaciones, como por ejemplo, las que conforman la microbiota intestinal humana (Figura 3) (Bikard et al., 2014; Gomaa et al., 2014).
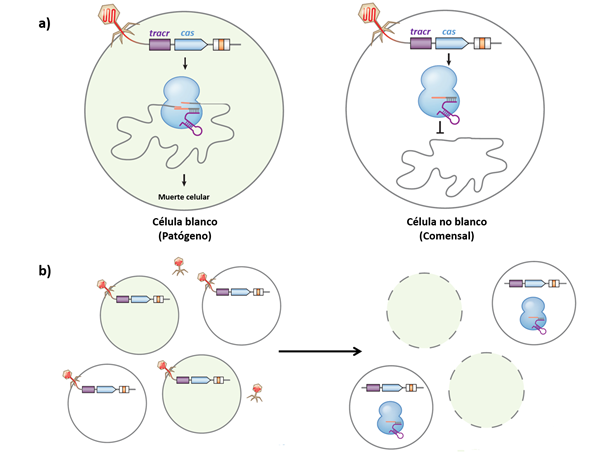
Figura 3 Uso del sistema CRISPR/cas como antimicrobiano secuencia específica. (a) En la mayoría de las bacterias con sistemas de reparación homóloga la acción de la nucleasa Cas con blancos específicos provoca la muerte celular debido a que los cortes en el DNA no pueden ser reparados. Para transportar al sistema es posible usar fagémidos a los cuales se les incorporan los genes que codifican los crRNA y a la proteína Cas. (b) En teoría, los fagémidos son específicos para un gen o secuencia de interés, por lo que al emplearse contra una población compleja las bacterias comensales permanecerán intactas. Modificado de Jiang & Marraffini. 2015. Ver la versión a color en línea.
Uno de los principales retos en el diseño de esta nueva estrategia ha sido idear la forma de llevar el sistema CRISPR/Cas hasta las células blanco. Una opción que está dando buenos resultados, es el uso de fagémidos, plásmidos empaquetados dentro de una cápside vírica, a los que se les han incorporado los genes que codifican la nucleasa Cas y el sgRNA modificado para atacar a los genes de interés (Citorik et al., 2014). Un ejemplo del uso exitoso de esta estrategia es el uso del fago filamentoso M13, donde de manera selectiva fue posible eliminar a una cepa de E. coli resistente a antimicrobianos. Además, se observó que el tratamiento con los fagémidos incrementó la supervivencia de la polilla de la cera, Galleria mellonella, que es un modelo para estudiar a E. coli EHEC 0157:H7 capaz de producir infecciones intestinales (Citorik et al., 2014). Otro caso exitoso es el uso del fago ΦNM1, donde los fagémidos fueron empleados para tratar una infección cutánea producida por Staphylococcus aureus en ratones (Yosef et al., 2015). Después del tratamiento algunas bacterias lograron sobrevivir, sin embargo, no contenían los genes de resistencia a antimicrobianos contra los que estaba dirigida la nucleasa Cas, por lo que el uso del sistema CRISPR/Cas permitió la eliminación de las bacterias que poseían el determinante de resistencia contra el que estaba dirigido (Yosef et al., 2015).
Hasta el momento, el uso de esta biotecnología para el tratamiento de infecciones bacterianas ha probado ser muy eficaz y tiene el potencial para el diseño de tratamientos personalizados, ya que puede utilizarse únicamente en las bacterias responsables de la infección o adaptarse para atacar genes de interés que codifican factores de resistencia a antimicrobianos, como las β-lactamasas de espectro extendido, o factores de virulencia, como los determinantes en la formación de biofilm.
Limitaciones del sistema CRISPR/Cas
Un requisito fundamental de cualquier sistema de edición genética es la ausencia, o al menos una baja frecuencia, de mutaciones secundarias accidentales. Para el caso del sistema CRISPR/Cas la especificidad está directamente relacionada con la capacidad de los sgRNA para reconocer su blanco, ya que son los responsables de guiar a la nucleasa Cas hasta el sitio donde serán introducidos los cortes en la molécula de DNA y posteriormente los sistemas de reparación se encargan de introducir o remover fragmentos de DNA (Figura 2) (Jiang & Marraffini, 2015). Debido a que se necesita un alineamiento perfecto entre el sgRNA y el DNA blanco para que el elemento Cas se una, la probabilidad de que se modifique una región distinta a la deseada es extremadamente baja. Los sgRNA necesitan de 20 nucleótidos para alcanzar su blanco (Jinek et al., 2012), por lo que la probabilidad para alcanzar un sitio equivocado ha sido predicho que es de 4-20 X 2 (debido a que la secuencia de 20 nucleótidos puede alinear en ambas hebras del DNA) (Jiang & Marraffini, 2015). Sin embargo, estudios en células bacterianas (Jackson et al., 2014) y la caracterización bioquímica del elemento Cas (Gasiunas et al., 2012) mostraron que no se necesita un alineamiento perfecto entre los primeros 5-10 nucleótidos del extremo 5´, por lo que la probabilidad de alcanzar una secuencia no deseada se incrementa.
Además de la generación de mutaciones indeseadas, una de las limitaciones más importantes del sistemas CRISPR/Cas es la ausencia de métodos eficaces para transportar al sistema dentro de las células de interés (Maggio & Gonçalves, 2015). La primera estrategia para transportar al sistema dentro de células bacterianas es la electroporación (Maggio & Gonçalves, 2015), donde se introduce un plásmido que contiene los genes que codifican para el sistema CRISPR/Cas exponiendo las células a pulsos eléctricos, desde luego que este sistema únicamente podría emplearse in vitro. Para un modelo más complejo, como lo es el de una infección bacteriana, donde está involucrado un huésped y bacterias comensales a los cuales se desea mantener intactos, el uso de fagémidos para transportar el sistema se asocia con una gran eficiencia y una baja citotoxicidad comparada con el uso de bacteriófagos (Citorik et al., 2014). No obstante, este sistema está sustancialmente limitado debido a que los fagémidos conservan el mismo blanco de infección que los bacteriófagos del cual derivan, por lo tanto, únicamente pueden infectar a las bacterias que expresen los receptores apropiados (Bikard et al., 2014). Además, los fagémidos son inmunogénicos, por lo que podrían desencadenar efectos no deseables en el huésped. Por otro lado, debido a que el sistema se distribuye en forma de DNA plasmídico, este podría ser neutralizado por la acción de enzimas de restricción (Bikard et al., 2014). Finalmente, una de las limitaciones más serias es la necesidad de realizar una caracterización detallada de la microbiota del paciente a tratar, con el objetivo de que los sgRNA diseñados sean altamente específicos.
Conclusiones
El desarrollo de un método de edición genética basado en el sistema CRISPR/Cas resalta la importancia de la investigación básica, ya que sus principios son los mecanismos de replicación y reparación del DNA, además del mecanismo de defensa contra la incorporación de DNA exógeno. Una vez que se descifró el mecanismo mediante el cual el sistema CRISPR/Cas confiere inmunidad contra moléculas de DNA invasoras, fue posible incorporar este sistema como una prometedora herramienta para la edición genética. Actualmente, la posibilidad para modificar genomas tanto de organismos procariotas como eucariotas mediante este sistema parece ser ilimitada, lo que a su vez, abre una gran diversidad de posibles aplicaciones médicas y biotecnológicas, mismas que ya se están explorando, como es el caso del diseño de terapias contra una gran cantidad de enfermedades, incluyendo las infecciones bacterianas.

