










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkTIP. Revista especializada en ciencias químico-biológicas
versión impresa ISSN 1405-888X
TIP vol.18 no.1 Ciudad de México jun. 2015
Artículo de revisión
Estado del conocimiento de la durofagia en el registro fósil: interacción depredador-presa en moluscos marinos (Clases Gastropoda y Bivalvia)
State of the art of durophagy in the fossil record: predator-prey interactions in marine mollusks (classes Gastropoda and Bivalvia)
Catalina Gómez-Espinosa1, Raúl Gío-Argáez2 y Miriam Carreño de la Vega3
1 Instituto de Ciencias Nucleares, UNAM, Circuito Exterior S/N, Ciudad Universitaria, Deleg. Coyoacán, C.P. 04510, México, D.F. E-mail: 1c_gomez@ciencias.unam.mx
2 Unidad Académica Procesos Oceánicos y Costeros, Instituto de Ciencias del Mar y Limnología, UNAM, Circuito Exterior S/N, Ciudad Universitaria, Deleg. Coyoacán, C.P. 04510, México, D.F. E-mail: 2raulgio@cmarl.unam.mx
3 Investigador independiente. 3mircve@hotmail.com
Artículo recibido el 30 de enero de 2015.
Aceptado el 07 de mayo de 2015.
Resumen
La depredación como un mecanismo evolutivo, de diversificación y biomineralización es un tópico de gran interés. Un tipo específico de depredación es la durofagia que se define como el hábito alimenticio de los animales que consumen organismos con un esqueleto duro mineralizado, ya sea concha o exoesqueleto. Los moluscos representan una fuente muy importante para examinar la depredación durófaga en el registro fósil debido a su distribución mundial, abundancia, buena preservación y a que habitan en una gran variedad de ambientes. En este trabajo se revisa el número de artículos publicados en revistas de circulación internacional que se enfocan en el tema de la durofagia y que tienen como potenciales presas a gasterópodos y bivalvos marinos fósiles. Se recuperaron y revisaron 101 artículos sobre este tema, enfocados en seis tipos de evidencia: perforación, reparación y fragmentación de la concha, mordidas, "punctures" y coprolitos. La mayoría de los ejemplos correspondieron a moluscos del Cenozoico. En general hay más registros en la zona tropical. Durante el Paleozoico, la mayoría de las veces, no se identificó el potencial depredador; esta tendencia continuó en el Mesozoico pero se reconoció la depredación de gasterópodos y bivalvos por parte de gasterópodos carnívoros, artrópodos, peces y reptiles. Durante el Cenozoico dominó la depredación por parte de gasterópodos natícidos y murícidos y se reconoció también la depredación por parte de artrópodos, peces y mamíferos.
Palabras Clave: Bivalvos, depredación, durofagia, fósiles, Gasterópodos.
Abstract
Predation as an evolutionary mechanism of diversification is a topic of great interest. Durophagy is the feeding behavior of preying on hard-shelled or exoskeleton-bearing organisms. On this matter, because of their worldwide distribution, abundance, wide range of habitats, and a remarkable preservation frequency, mollusks are of particular interest in the study of predation in the fossil record. In the present work, papers published in international scientific magazines focusing on the issue of durophagy on fossil marine bivalve and gastropods as potential prey were analyzed. A total of 101 papers on this subject were reviewed. Six-kinds of evidence of durophagy were identified: drilling, repaired and fragmented shells, bites, punctures and coprolites. Most items are of Cenozoic age and the majority of the records available correspond to the tropics. Most Paleozoic potential predators were not identified; this situation extends to the Mesozoic, but predation of gastropods and bivalves by carnivorous gastropods, arthropods, fishes and reptiles was recognized. Predation by naticid and muricid gastropods prevails during the Cenozoic, also was recognized predation by arthropods, fishes and mammals.
Key Words: Bivalves, predation, durophagy, fossils, Gastropods.
Introducción
Las interacciones bióticas son aquellas que se establecen entre dos o más seres vivos dentro de un ecosistema, que les confiere un papel específico dentro de su hábitat. En el caso particular de la interacción conocida como depredación, lo que ocurre es el consumo de un organismo (la presa) por otro organismo (el depredador), estando viva la presa cuando es atacada por el depredador1. Una de las variantes de la depredación es la durofagia.
Se considera que la depredación ha tenido un papel muy importante en la evolución, diversificación y biomineralización de los metazoarios2-5, por lo que se han estudiado las características morfológicas y etológicas de algunos grupos de depredadores para identificar los posibles patrones que influyeron en su evolución6.
La durofagia es el hábito alimenticio de los animales que consumen organismos con un esqueleto duro mineralizado ya sean concha o exoesqueleto7, por lo que aparece después del "evento de biomineralización" asociado a la radiación cámbrica8 y la "revolución del sustrato"9. El primer registro fósil de durofagia corresponde a perforaciones encontradas en Cloudina uno de los géneros más antiguos que posee un esqueleto mineralizado10.
Vermeij11, 12 utilizó el término durofagia para denominar los procesos de trituración de las conchas y perforaciones ocasionadas por carnívoros depredadores.
La depredación durófaga ha sido tema de investigación en gran variedad de invertebrados fósiles y recientes, dentro de los que se encuentran los cnidarios13, conuláridos14, serpúlidos15, braquiópodos16, equinodermos17, moluscos18 y artrópodos19; sin embargo, la mayoría de la literatura sobre durofagia se ha centrado en el estudio de la depredación de corales recientes por parte de peces20.
Los moluscos representan una de las fuentes de información más importantes para estudiar el registro fósil de la depredación, debido a que tienen una distribución mundial, son muy abundantes, están bien preservados y se conservan en una gran variedad de ambientes21. La principal evidencia de depredación de organismos malacófagos en el registro fósil son las huellas (perforaciones, cicatrices de reparación, marcas de mordidas y fracturas)22.
Los organismos durófagos que se alimentan de moluscos tienen cuatro métodos de depredación: la fragmentación pre-ingestiva o trituración de la concha, la ingestión completa del organismo, la inserción y extracción y la perforación2, 23.
La ingesta completa del molusco y la extracción e ingestión no dejan una evidencia reconocible y/o preservable en el registro fósil24; la trituración pre-ingestiva y la perforación son los dos métodos por los que puede reconocerse la depredación en gasterópodos y bivalvos, permitiendo diferenciar incluso los daños letales de los no letales24.
El daño no letal en las conchas de moluscos se reconoce a través de las cicatrices dejadas por la sanación de lesiones25, ya que los moluscos pueden reparar su concha, mientras que el manto permanezca intacto26.
La trituración de la concha de los moluscos por depredadores es considerada como una de las principales causas de selección que favoreció la evolución de la resistencia de la concha, la cual a través del tiempo se ha vuelto más eficiente en la protección de los organismos contra sus depredadores y otros agentes letales2,11,27.
El estudio de la frecuencia de reparación en conchas de moluscos marinos ha sido fundamental para sustentar la Hipótesis de la Escalación propuesta por Vermeij11 y que propone a la depredación como fuerza macroevolutiva, donde las defensas cada vez son más sofisticadas porque el ambiente es más agresivo28.
Durante el Paleozoico Inferior, y particularmente de la fauna cámbrica de Burgess Shale, se ha propuesto a los anomalocáridos y a Sidneyia como los probables representantes de depredadores malacófagos29. Para el Paleozoico Medio se propone la diversificación de los durófagos30. De este modo, entre los invertebrados depredadores de moluscos, se encontraban los crustáceos malacostráceos16, probablemente los amonites (su dieta es incierta) y los nautiloideos, considerados por algunos autores como malacófagos31, mientras que otros los excluyen por carecer de mandíbulas calcificadas30. Dentro de los vertebrados malacófagos paleozoicos se consideran a los peces placodermos y condrictios30.
Como depredadores de moluscos durante el Mesozoico se proponen, entre los invertebrados, a los palinúridos32, braquiuros y pagúridos24 y entre los vertebrados depredadores se incluyen a los peces (picnodontes y condrictios)24 y a los reptiles (placodontes24, mosasáuridos33 y quelonios34).
Durante el Cenozoico se considera que los invertebrados malacófagos estuvieron conformados por neogasterópodos24, algunos cefalópodos35, braquiuros24 y estomatópodos36. Dentro de los vertebrados se incluye a peces teleósteos37, aves marino-costeras2 y buceadoras y algunos mamíferos marinos24.
Este artículo consiste en la revisión y análisis de la literatura existente respecto al tópico de durofagia en gasterópodos y bivalvos fósiles, para estudiar el estado del conocimiento en este tema, considerando: la tendencia en el número de artículos publicados por año, el tipo de evidencia a través del cual se identifica la durofagia, la proporción de artículos publicados por era y periodo geológico, los potenciales depredadores a través del Fanerozoico, la zona latitudinal donde es más frecuente el registro de durofagia y los países con mayor número de registros. La revisión de la literatura existente sobre la depredación durófaga en el registro fósil se destaca por la relevancia evolutiva que representa para los moluscos, además de ser un tema que hasta el momento no ha sido objeto de estudio por parte de los paleontólogos en México.
Materiales y Métodos
Se analizó la producción de artículos publicados en revistas de circulación internacional cuyos objetivos estaban enfocados en la durofagia, teniendo como presas potenciales a gasterópodos y/o bivalvos marinos fósiles.
La búsqueda detallada de publicaciones se llevó a cabo a través de Web of Science, Google Scholar, GeoRef Information Services, Geomex, ASFA, GeoBase, Scopus, Librunam.
Las palabras clave que se utilizaron para la búsqueda fueron "durophagy" "drilling predation" "shell crushing predation" "shell repair" "trace predation" y "Oichnus"; incluyendo en todas el término "fossil" y variando el uso de los términos "mollusk" "mollusc" "bivalves" "pelecypods" y/o "gastropods".
No se tomaron en cuenta los artículos sobre tafonomía actualística que estudia la durofagia en moluscos recientes y propone modelos experimentales aplicables al registro fósil.
Los datos de los artículos localizados se ordenaron de acuerdo con el periodo geológico, del más antiguo al más reciente y dentro de éstos, se organizó de manera alfabética de acuerdo con el autor.
A partir de los datos de las citas recuperadas se realizaron diferentes gráficas para: comparar el número de artículos por año de publicación, por tipo de evidencia fósil (proxies) de malacofagia, entre periodos y eras geológicas y, por los países y/o localidades donde fueron reportados.
Resultados
Se recuperaron y revisaron 101 artículos, cuyo tema de investigación es la depredación durófaga en gasterópodos y bivalvos marinos a lo largo del Fanerozoico.
Seis de las publicaciones se catalogaron como revisiones, al tratarse de trabajos que consideran, en términos generales, la durofagia en bivalvos38 y en gasterópodos y bivalvos24; a los decápodos como depredadores de moluscos39; a los gasterópodos como depredadores durófagos40 y los patrones de depredación marina post-paleozoica2,41; en el caso de los artículos de gasterópodos durófagos40 y de patrones de depredación2,41 se consideran varios taxones como presas potenciales dentro de los que se incluye, además de los moluscos, a los braquiópodos y a los equinodermos.
El primer estudio sobre durofagia, publicado en 1938, corresponde al registro de perforaciones en la concha de un gasterópodo cretácico en una localidad de Estados Unidos42; mientras que la publicación más reciente, del 2014, estudia la fragmentación por depredación en bivalvos plio-pleistocénicos también de una localidad de Estados Unidos43.
Desde la primera publicación y durante un lapso de 32 años, de 1938 a 1970, sólo se publicaron 4 artículos sobre depredación durófaga en gasterópodos y bivalvos marinos fósiles. A partir de 1970, las publicaciones se volvieron más constantes; sin embargo, el número de artículos publicados se mantuvo bajo, pues se cuenta con una o dos publicaciones por año. Este número se elevó a siete publicaciones, únicamente para los años de 1997, 2008 y 2010 (Fig. 1).
Las publicaciones de durofagia en gasterópodos y bivalvos marinos fósiles se basan en seis tipos de evidencia o proxies de depredación: la perforación, la reparación, la fragmentación, las marcas de mordidas, las trazas de tipo "puncture" y en coprolitos (Fig. 2).
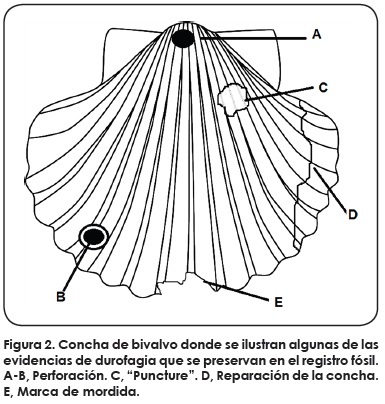
La mayoría de los estudios han utilizado la reparación y perforación de la concha como evidencia de durofagia. La comparación entre ambas clases de moluscos arroja como resultado que hay mayor número de publicaciones de conchas reparadas para gasterópodos (20 publicaciones) que para bivalvos (3 publicaciones); mientras que los registros de perforación son mayores en bivalvos (45 publicaciones) que en gasterópodos (33 publicaciones) (Fig. 3).
Respecto a la fragmentación por depredación, hay ocho publicaciones para gasterópodos y siete para bivalvos. De estas publicaciones, dos corresponden a revisiones sobre la fragmentación por depredación24,39, una corresponde a estudios en gasterópodos del Mesozoico12, otra estudia a bivalvos mesozoicos44 y una más, a bivalvos cenozoicos43; las cuatro restantes incluyen tanto a gasterópodos como a bivalvos del Mesozoico45,46 y Cenozoico47,48.
Las trazas tipo "puncture" son las marcas dejadas como consecuencia de la depredación por parte de crustáceos (decápodos y estomatópodos) que semejan punciones. Este tipo de evidencia se registró, en dos publicaciones, en conchas de gasterópodos del Neógeno49 y Cuaternario50. En el caso de los bivalvos, hay un registro de una posible "puncture", aunque se argumenta que la evidencia no es muy clara y que también podría tratarse de depredación por parte de un pez o de un reptil marino51.
Las evidencias de depredación de las que existe un menor número de publicaciones son de las mordidas y los coprolitos, que hasta el momento han sido identificadas únicamente para bivalvos cretácicos. Estas publicaciones corresponden a un registro de un coprolito de tortuga34 con un fragmento de bivalvo, y dos a marcas de mordidas, una atribuida a un tiburón52 y otra causada por un vertebrado no identificado53.
Paleozoico
Se recuperaron 25 publicaciones de depredación en gasterópodos y bivalvos paleozoicos y una más que incluye ejemplares paleozoicos, además de otras eras. Comparando el número de publicaciones entre periodos, hay mayor cantidad de publicaciones para ejemplares del Ordovícico en localidades de Canadá, Estados Unidos, República Checa y Suecia; en segundo lugar se encuentran ejemplares del Silúrico con publicaciones para localidades de Bélgica, Escocia, República Checa, Suecia y ejemplares de museos. El periodo con menor número de registros es el Cámbrico, con un único registro para Australia (Fig. 4).
En 74% de las publicaciones para la Era Paleozoica no se identifica el depredador potencial y sólo se menciona como depredador desconocido. En 8.7% de las publicaciones se propone más de un organismo como posible responsable de la depredación, considerando dentro de éstos a los ofiuroideos, nautiloideos, equinoideos y peces. Para 17.3% de las publicaciones se identifica con certeza al depredador y se mencionan entre éstos a los artrópodos, gasterópodos natícidos y nautiloideos ortocónicos.
Mesozoico
Se recuperaron 24 artículos sobre depredación en gasterópodos y bivalvos mesozoicos y seis más, que incluyen ejemplares del Mesozoico junto con otras eras. De las publicaciones analizadas, los trabajos de Vermeij11 y Tatsuo et al.46 consideran todo el Mesozoico, razón por la cual se contaron para los tres periodos de esta era. El primer trabajo corresponde a un estudio de ejemplares de museo; mientras que el segundo se realizó con ejemplares de localidades de Japón. Respecto al número de publicaciones sobre durofagia, por periodo, el mayor número corresponde a ejemplares del Cretácico, en localidades de Australia, Estados Unidos, India, Inglaterra, Nueva Zelanda y ejemplares de museo. En segundo lugar se encuentran las publicaciones de ejemplares del Triásico, en localidades de Inglaterra y Polonia y el menor número corresponde a ejemplares del Jurásico, en localidades de Japón (Fig. 5).
Respecto a los potenciales depredadores durófagos durante esta era, en 22% de las publicaciones no se identifica o se menciona como desconocido, en 39% de los artículos la depredación se adjudica a gasterópodos carnívoros, 28% considera la depredación por vertebrados y el 11% restante menciona que no hay evidencia suficiente para distinguir si la depredación fue por crustáceos, peces o reptiles.
Cenozoico
Se recuperaron 54 artículos sobre depredación en moluscos cenozoicos. El mayor número de artículos se enfocó en el estudio de ejemplares del Neógeno, en localidades de Alemania, Australia, Austria, Bélgica, Eslovaquia, Estados Unidos, Francia, Hungría, India, Países Bajos y Polonia. El menor número de publicaciones corresponde a ejemplares del Paleógeno en localidades de Canadá, Estados Unidos, Gales, los océanos Atlántico y Pacífico y ejemplares de museo (Fig. 6).
Durante el Cenozoico, los principales depredadores malacófagos marinos identificados en el registro fósil fueron anélidos, gasterópodos carnívoros, crustáceos, peces y mamíferos.
El 72% de las publicaciones adjudica la depredación a gasterópodos de las familias Naticidae y Muricidae; 10% considera una depredación por crustáceos (decápodos y estomatópodos); en 4 % se reconoce la depredación por parte de peces; 2% reportan depredación por anélidos poliquetos; otro 2% registra la depredación por mamíferos pinnípedos; y en el 10% restante de las publicaciones no se identifica al posible depredador.
Comparando el número de publicaciones disponible para las diferentes eras, la mayor parte de los artículos tiene como objeto de estudio la depredación de bivalvos y gasterópodos cenozoicos; mientras que el menor número de registros corresponde a ejemplares del Mesozoico.
Respecto a las zonas geográficas, existen publicaciones sobre durofagia en gasterópodos y bivalvos marinos fósiles para localidades de Alemania, el Océano Atlántico, Australia, Austria, Bélgica, Brasil, Canadá, Costa Rica, Escocia, Eslovaquia, España, Estados Unidos, Francia, Hungría, India, Inglaterra, Islas Fiji, Italia, Japón, Nigeria, Nueva Zelanda, el Océano Pacífico, Países Bajos, Panamá, Polonia, República Checa y Suecia. Las publicaciones para localidades de Estados Unidos representan 38.6 % del total; mientras que para localidades de Alemania, Costa Rica, Escocia, Las Islas Fiji, Japón, Nueva Zelanda, Panamá y Suecia existe sólo una publicación, lo que representa 1% del total para cada país (Fig. 7).
Discusión
Gran parte de las publicaciones sobre depredación en moluscos, a través del Fanerozoico, están basadas en hipótesis que tratan de inferir el grado de depredación para cada era y cómo esta interacción ha ido evolucionando en el tiempo geológico. El principal modelo macroevolutivo a este respecto corresponde a la "Hipótesis de la Evolución Escalada" de Vermeij2, 11, donde se propone una carrera armamentista entre presas y depredadores.
Los trabajos de revisión sobre depredación hacen referencia a una gran cantidad de estudios sobre durofagia en el Reciente, tanto en acumulaciones naturales como en estudios experimentales. Además, en muchos casos, la posible conducta depredadora de los organismos y sus presas potenciales se infiere a través de evidencia indirecta, analizando las adaptaciones morfológicas de presas y depredadores.
Las publicaciones basadas en evidencia física sobre depredación en moluscos, aunque han aumentado considerablemente a partir de la década de 1980, son escasas. Además de que la mayoría no considera la relación entre los tipos de hábitats, las especies y el intervalo de tiempo estratigráfico.
La escasa evidencia de durofagia puede deberse, en parte, a que la fragmentación, perforación, mordidas y "punctures" disminuyen el potencial de preservación del esqueleto de la presa en el registro fósil, lo que resulta en un sesgo en las estimaciones cuantitativas basadas en la frecuencia de las huellas21. Además de que la relativamente escasa publicación de artículos sobre este tema puede estar relacionada con la dificultad de la identificación de la evidencia de durofagia en las conchas de los moluscos, pues algunos de los proxies que se utilizan son controversiales. En el caso de las conchas reparadas, para que se consideren como evidencia de durofagia, tiene que descartarse la posibilidad de un daño mecánico o la acción de otros factores destructivos7.
En el caso de las conchas fragmentadas como evidencia de durofagia en el registro fósil, la problemática que se presenta para su identificación es mayor, pues las conchas pueden romperse debido a la energía del ambiente y/o a retrabajo físico y sólo es posible reconocer la depredación analizando los tipos de bordes que presentan los fragmentos43.
Los proxies más confiables son las perforaciones, las marcas de mordidas y los coprolitos; las perforaciones de gasterópodos carnívoros sobre conchas de otros gasterópodos y bivalvos, son abundantes y fáciles de reconocer; sin embargo, las marcas de mordidas en las conchas y los coprolitos con restos orgánicos son relativamente infrecuentes en el registro fósil.
También hay que considerar que las evidencias deben interpretarse de acuerdo con las eras, pues Hoffmeister et al.54 estiman que los registros de perforación en bivalvos para el Paleozoico son controversiales; pero para el Cenozoico se cuenta con registros muy confiables e incluso con estudios cuantitativos a gran escala.
Respecto al tiempo geológico y la depredación durófaga, se considera que durante el Paleozoico temprano (Cámbrico-Ordovícico) los organismos durófagos estaban poco especializados y por tanto, eran poco eficientes para fragmentar las conchas2. Y no fue sino hasta el Devónico (Paleozoico Medio), cuando aparecieron los peces mandibulados, que se adquirió una mayor capacidad para depredar a organismos con esqueletos duros30.
Signor y Brett30 estiman que durante el Paleozoico Medio se dio una gran diversificación de depredadores, que derivó en la evolución y diversificación de las comunidades bentónicas55 y que sentó las bases de la denominada "Revolución Marina Mesozoica".
De acuerdo con la evidencia fósil, durante el Paleozoico inferior (Ordovícico56) y superior (Carbonífero57), la frecuencia de reparación de las conchas en los gasterópodos era baja, lo que pudo ser el resultado de la deficiencia de sus depredadores.
Aunque los gasterópodos y bivalvos marinos se diversificaron a principios del Mesozoico, los registros de durofagia para esta era geológica son muy escasos, esto puede estar relacionado con el hecho de que los durófagos mesozoicos se alimentaban de cualquier presa con concha dura y no eran depredadores especialistas de moluscos21. Durante el Triásico, las conchas de los gasterópodos presentaban una débil adaptación contra la durofagia, debido al poco desarrollo de las características morfológicas contra la fragmentación letal. Para el Jurásico, se dio un cambio en la morfología y la tendencia favoreció el desarrollo de las características que confirieron resistencia a la concha (ej. abertura estrecha y labio exterior grueso o dentado) y se disminuyeron las características que le conferían debilidad (ej. espira abierta y presencia de ombligo)2,11.
En el caso de las perforaciones se ha argumentado que su escasa evidencia en gasterópodos y bivalvos mesozoicos pudo ser el reflejo de una ineficiencia depredatoria durófaga durante el Triásico y de una evolución exitosa de ésta durante el Cretácico58. La evolución y especialización de los depredadores durófagos durante el Mesozoico Medio2,11,12 se ha interpretado por el aumento en el tamaño y grosor de las conchas en moluscos cretácicos y se vio reflejado en el registro fósil como un incremento de los registros de conchas reparadas.
En términos generales, puede considerarse que la durofagia fue mayor durante el Mesozoico y Cenozoico, de lo que fue en el Paleozoico2,11 y que fue durante el Mesozoico cuando ocurrió una importante diversificación de los depredadores de moluscos.
El aumento de la durofagia durante el Cenozoico puede deberse a un aumento en el número de depredadores malacófagos y también hay que tomar en cuenta el posible sesgo tafonómico, pues hay una mayor cantidad de fósiles para la era geológica más reciente, en tanto que los restos más antiguos tienden a destruirse debido a la dinámica del planeta.
El mayor número de publicaciones sobre durofagia para localidades de Estados Unidos muestra un mayor interés por parte de los investigadores de ese país en el estudio de la moluscivoría en el registro fósil. Hasta el momento no existe publicación alguna sobre durofagia en moluscos fósiles de localidades de México, por lo cual hay un campo por explorar dentro de la investigación paleontológica mexicana, el cual podría iniciarse revisando los ejemplares de museo.
Con respecto a la zona latitudinal, la durofagia mantiene la misma tendencia que se observa en la actualidad27, siendo mayor en la zona tropical y disminuyendo hacia la zona de los polos. Por ejemplo, el registro de durofagia para Australia corresponde al Cámbrico5, periodo durante el cual, paleogeográficamente, Australia se localizaba en la zona ecuatorial59.
Las interacciones bióticas que se establecieron entre los gasterópodos y bivalvos marinos y sus depredadores no es muy clara durante el Paleozoico pues la mayor parte de las trazas de depredación no se adjudican a algún organismo en particular, e incluso en varios artículos sólo se estudia la depredación y se omite el tratar de identificar al posible depredador. Algunos de los depredadores que sí se identifican para esta era son nautiloideos ortocónicos y, probablemente ofiuroideos juveniles, equinoideos y peces teleósteos como depredadores de gasterópodos; así mismo, se ha considerado a los gasterópodos natícidos y artrópodos como depredadores de bivalvos.
Durante el Mesozoico, las relaciones depredador-presa son más claras y aunque también hay publicaciones que mencionan depredadores desconocidos, hay evidencia de que los gasterópodos y bivalvos marinos fueron depredados por gasterópodos carnívoros (en algunos casos se identifica a los miembros de la familia Naticidae), crustáceos y peces teleósteos y cartilaginosos. En el caso de los bivalvos, también se tienen registros de depredación por parte de reptiles marinos.
Durante el Cenozoico, la depredación de gasterópodos y bivalvos fue dominada por gasterópodos natícidos y murícidos y también hay evidencia de depredación por parte de crustáceos (decápodos y estomatópodos) y peces teleósteos. Los gasterópodos, además, fueron depredados por anélidos poliquetos y los bivalvos, por parte de mamíferos pinnípedos (morsas).
Conclusiones
Las publicaciones sobre durofagia en gasterópodos y bivalvos marinos fósiles son escasas, aunque han aumentado considerablemente a partir de la década de 1980 y se basan en seis tipos de evidencia de depredación: la perforación, la reparación y fragmentación de las conchas, las marcas de mordidas e icnitas tipo "puncture" y coprolitos. El proxy de durofagia del que existe mayor número de publicaciones es la perforación, tanto en gasterópodos como en bivalvos y en segundo lugar se encuentra la reparación de las conchas. De acuerdo a la bibliografía consultada, se considera que el fenómeno de depredación fue menor durante el Paleozoico, de acuerdo con las evidencias que se observaron en los ejemplares de esa Era y aumentó, junto con la diversificación de los depredadores, durante el Mesozoico y Cenozoico. Paleogeográficamente, la mayoría de las localidades con registros de durofagia se ubican en la zona tropical y disminuyen hacia la zona boreal, siendo incluso nulas las publicaciones para la zona polar.
La evidencia fósil muestra que los gasterópodos y bivalvos marinos han sido depredados principalmente por gasterópodos carnívoros, artrópodos y peces. Los artículos sobre depredación de ejemplares paleozoicos no identifican, en la mayoría de los casos, al depredador potencial; para ejemplares del Mesozoico, aunque también hay depredadores no identificados, se reconoce la depredación por parte de gasterópodos, artrópodos, peces y reptiles y para el Cenozoico, se muestra una dominancia de depredación por parte de gasterópodos carnívoros.
Agradecimientos
Los autores agradecen a los revisores anónimos sus sugerencias, comentarios y recomendaciones para mejorar este trabajo.
Referencias
1. Begon, M., Townsend, C.R. & Harper, J.L. Ecology from individuals to ecosystems. (Wiley-Blackwell, United Kingdom, 2006). [ Links ]
2. Vermeij, G.J. Evolution and escalation, an ecological history of life. (Princeton University Press, Princeton, 1987). [ Links ]
3. Morris, S.C. & Bengtson, S. Cambrian predators: possible evidence from boreholes. Journal of Paleontology 68, 1-23 (1994). [ Links ]
4. Kowalewski, M. & Kelley, P.H. The fossil record of predation. The Paleontology Society Papers 8, 1–398 (2002). [ Links ]
5. Skovsted, C.B., et al. Early Cambrian record of failed durophagy and shell repair in an epibenthic mollusk. Biology Letters 3, 314-317 (2007). [ Links ]
6. Walker, S.E. & Brett, C.E. The fossil record of shell boring by snails Post Paleozoic patterns in marine predation: was there a Mesozoic and Cenozoic marine predatory revolution?. Paleontological Society Papers 8, 119-194 (2002). [ Links ]
7. Aronson, R.B. in Palaeobiology II (ed. Briggs, D.E.G. & Crowther P.R.) 393–397 (Blackwell Science, Oxford, 2001). [ Links ]
8. Marshall, C.R. Explaining the Cambrian "explosion" of animals. Annual Review of Earth Planetary Sciences 34, 355–384 (2006). [ Links ]
9. Bottjer, D.J., Hagadorn, J.W. & Dornbos, S.Q. The Cambrian substrate revolution. GSA Today 10, 1–7 (2000). [ Links ]
10. Bengtson, S. & Yue, Z. Predatorial borings in Late Precambrian mineralized exoskeletons. Science 257, 367–369 (1992). [ Links ]
11. Vermeij, G.J. The Mesozoic marine revolution: Evidence from snails, predators, and grazers. Paleobiology 3, 245–258 (1977). [ Links ]
12. Vermeij, G.J. Traces and trends of predation, with special reference to bivalve animals. Palaeontology 26, 455-465 (1983). [ Links ]
13. Galle, A. & Mikulas, R. Evidence of predation on the rugose coral Calceola sandalina (Devonian, Czech Republic). Ichnos 10, 41-45 (2003). [ Links ]
14. Mapes, R.H., Fahrer, T.R. & Babcock, L.E. Sublethal and lethal injuries of Pennsylvanian conularids from Oklahoma. Journal of Paleontology 63, 34–37 (1989). [ Links ]
15. Martinell, J., Kowalewski, M. & Domenech, R. Drilling predation on serpulid polychaetes (Ditrupa arietina) from the Pliocene of the Cope Basin, Murcia Region, Southeastern Spain. PLoS ONE 7, e34576 (2012). [ Links ]
16. Hoffmeister, A.P., Kowalewski, M., Bambach, R.K. & Baumiller, T.K. Intense drilling predation in the Carboniferous brachiopod Cardiarina cordata Cooper 1956. Lethaia 36, 107–118 (2003). [ Links ]
17. Baumiller, T.K. & Gahn, F.J. Reconstructing predation pressure on crinoids: estimating arm-loss rates from regenerating arms. Paleobiology 39, 40-51 (2012). [ Links ]
18. Klompmaker, A.A. Drilling and crushing predation on scaphopods from the Miocene of the Netherlands. Lethaia 44, 429-439 (2011). [ Links ]
19. Ruiz, F. et al. Predation on Neogene ostracods of Southwestern Spain. Rivista Italiana di Paleontologia e Stratigrafia 116, 253-260 (2010). [ Links ]
20. Huber, D.R., Eason, T.G., Hueter, R.E. & Motta, P.J. Analysis of the bite force and mechanical design of the feeding mechanism of the durophagous horn shark Heterodontus francisci. J. Exp. Biol. 208, 3553-3571 (2005). [ Links ]
21. Kowalewski, M. The fossil record of predation: an overview of analytical methods. Paleontological Society Papers 8, 3-41 (2002). [ Links ]
22. Lindstrom, A. & Peel, J.S. Repaired injuries shell form in some Paleozoic pleurotomarioid gastropods. Acta Palaeontologica Polonica 50, 697-704 (2005). [ Links ]
23. Harper, E.M. & Skelton, P.W. The Mesozoic marine revolution and epifaunal bivalves. Scripta Geologica Special Issue 2, 127-153 (1993). [ Links ]
24. Alexander, R. R. & Dietl, G. P. in Predator - prey interactions in the fossil record (ed. Kelley, P.H., Kowaleski, M. & Hansen, T.H.) 141- 176 (Kluwer Academic/Plenum Publishers, New York, 2003). [ Links ]
25. Lindstrom, A. & Peel, J.S. Shell repair and shell form in Jurassic pleurotomarioid gastropods from England. Bulletin of Geosciences 85, 541-550 (2010). [ Links ]
26. Vermeij, G.J., Zipser, E. & Zardini, R. Breakage-induced shell repair in some gastropods from the Upper Triassic of Italy. Journal of Paleontology 56, 233-235 (1982). [ Links ]
27. Vermeij, G.J. in The fossil record of predation (ed. Kowalewsky, M. & Kelley, P.H.) 375–393 (Kluwer Academic/Plenum Publishers, New York, 2002). [ Links ]
28. Vermeij, G.J., Schindel, D.E. & Zipser, E. Predation through geological time: evidence from gastropod shell repair. Science 214, 1024-1026 (1981). [ Links ]
29. Conway, S. & Whittington, H.B. The animals of the Burgess Shale. Scientific American 241, 122-133 (1979). [ Links ]
30. Signor, P.W. & Brett, C.E. The Mid-Paleozoic precursor to the Mesozoic marine revolution. Paleobiology 10, 229-245, (1984). [ Links ]
31. Alexander, R.R. Resistance to and repair of shell breakage induced by durophages in Late Ordovician brachiopods. Journal of Paleontology 60, 273-285 (1986). [ Links ]
32. Harper, D.E. Trends in the spiny lobster commercial fishery of Florida, 1960-1990. Natural Marine Fishery MIN-91/92-01, 1-29, (1991). [ Links ]
33. Massare, J.A. Tooth morphology and prey preference of Mesozoic marine reptiles. Journal of Vertebrate Paleontology 7, 121-137 (1987). [ Links ]
34. Kear, B.P. First gut contents in a Cretaceous sea turtle. Biology Letters 2, 113–115 (2006). [ Links ]
35. Anderson, R.C. Octopus bites clam. The Festivus 26, 58-59 (1994). [ Links ]
36. Baluk, W. & Radwanski, A. Stomatopod predation upon gastropods from Korytnica basin, and from other classical Miocene localities in Europe. Acta Geologica Polonica 46, 79-304 (1996). [ Links ]
37. Vermeij, G.J. A natural history of shells (Princeton University Press, Princeton, 1993). [ Links ]
38. Carter, R.M. On the Biology and Paleontology of some predators of bivalved Mollusca. Palaeogeography, Paleoclimatology, Paleoecology 4, 29-65 (1968). [ Links ]
39. Schweitzer, C. E. & Feldmann, R.M. The decapoda (Crustacea) as predators on mollusca through geologic time. Palaios 25, 167-182 (2010). [ Links ]
40. Sohi, N.F. The fossil record of shell boring by snails. American Zoologist 9, 725-734 (1969). [ Links ]
41. Leighton, L.R. Inferring predation intensity in the marine fossil record. Paleobiology 28, 328-342 (2002). [ Links ]
42. Bucher, W.H. A shell-boring gastropod in a Dalmanella bed of Upper Cretaceous age. American Journal of Science 236, 1-7 (1938). [ Links ]
43. Mondal, S., Harries, P.J., Paul, S. & Herbert, G.S. Paleoecological significance of coupling metrics of successful and unsuccessful shell-breaking predation: examples using Neogene bivalve prey. Palaeogeography, Palaeoclimatology, Palaeoecology 399, 89-97 (2014). [ Links ]
44. Speden, I.G. Predation on New Zealand Cretaceous species of Inoceramus (Bivalvia) New Zealand. Journal of Geology and Geophysic 14, 56-60 (1971). [ Links ]
45. Zaton, M. & Salamon, M.A. Durophagous predation on Middle Jurassic molluscs as evidenced from shell fragmentation. Palaeontology 51, 63–70 (2008). [ Links ]
46. Tatsuo, O., Chigusa, O. & Takehiro, S. Increase of crushing predation recorded in fossil shell fragmentation. Paleobiology 29, 520-526 (2003). [ Links ]
47. Merle, D. Premiere etude taphonomique de la predation affectant de grands mollusques bentiques dans l'Eocene de Gan (Pyrenées-Atlantiques, France), Comptes Rendus de l' Academie des Sciences, series II. Sciencies de la Terre et des Planetes 330, 217-220 (2000). [ Links ]
48. Oji, T., Ogaya, C.V. & Sato, T. Increase in shell-crushing predation recorded in shell fragmentation. Paleobiology 29, 520-526 (2003). [ Links ]
49. Baluk, W. & Radwanski, A. Stomatopod predation upon gastropods from the Korytnica Basin, and from other classical Miocene localities in Europe. Acta Geologica Polonica 46, 279-304 (1996). [ Links ]
50. Geary, D.H., Howard, J.D. & Reaka-Kudla, J.S. Stomatopod predation on fossil gastropods from the Plio-Pleistoceno of Florida. Journal of Paleontology 65, 355-360 (1991). [ Links ]
51. Phipps, K.J. Evidence of predation on Gryphaea (Bilobissa) lituola Lamarck, 1819 from the Oxford Clay Formation of South Cave. Proceedings of the Geologists Association 119, 277-285 (2008). [ Links ]
52. Kauffman, E.G. Ptychodus predation in a Cretaceous Inoceramus. Paleontology 15, 439–444 (1972). [ Links ]
53. Kear, B.P. Inferred vertebrate bite marks on an Early Cretaceous unionoid bivalve from Lightning Ridge, New South Wales, Australia. Alcheringa 32, 65-71 (2008). [ Links ]
54. Hoffmeister, A.P., Kowalewski, M., Baumiller, T.K. & Bambach, R.K. Drilling predation on Permian brachiopods and bivalves from the Glass Mountains, west Texas. Acta Palaeontologica Polonica 49, 443–454 (2004). [ Links ]
55. Bambach, R.K. Seafood through time: Changes in biomass, energetics, and productivity in the marine ecosystem. Paleobiology 19, 372-397 (1993). [ Links ]
56. Ebbestad, J.O.R. & Stott, C.A. Failed Predation in Late Ordovician gastropods (Mollusca) from Manitoulin Island, Ontario Canada. Canadian Journal of Earth Sciences 45, 231-241 (2008). [ Links ]
57. Schindel, D.E., Vermeij, G.J. & Zipser, E. Frequencies of repaired shell fractures among the Pennsylvanian gastropods of north central Texas. Journal of Paleontology 56, 729-740 (1982). [ Links ]
58. Kowalewski, M., Dulai, A. & Fürsich, F.T. A fossil record full of holes: The Phanerozoic history of drilling predation. Geology 26, 1091-1094 (1998). [ Links ]
59. Scotese, C.R. PaleomapProject. http://www.scotese.com/earth.htm. Consultado junio 2014. (2010). [ Links ]
Información sobre los autores
Catalina Gómez-Espinosa
Catalina Gómez-Espinosa estudió la Licenciatura en Biología en la Facultad de Ciencias de la Universidad Nacional Autónoma de México y la Maestría y Doctorado en Ciencias Biológicas en el Instituto de Geología de la misma universidad. Realizó estancias posdoctorares en el Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México en México, D.F. y en el Instituto Argentino de Oceanografía en Bahía Blanca, Argentina. Ha participado en proyectos nacionales e internacionales y ha escrito artículos científicos y de divulgación, así como capítulos de libros; es coautora de un libro de apoyo docente. Actualmente realiza una estancia posdoctoral en el Instituto de Ciencias Nucleares de la Universidad Nacional Autónoma de México y es profesora de Asignatura en la Facultad de Ciencias y en la Facultad de Ingeniería de la Universidad Nacional Autónoma de México. Sus estudios se han enfocado en invertebrados bentónicos marinos, principalmente moluscos, desde una perspectiva tafonómica y de tafonomía actualística.
Raúl Gío-Argáez
Raúl Gío-Argáez es Doctor en Ciencias, profesor de Paleobiología e investigador en la Unidad de Procesos Oceánicos y Costeros del Instituto de Ciencias del Mar y Limnología, todo lo anterior en la UNAM. Ha publicado 115 trabajos entre libros, artículos para revistas indizadas/ arbitradas, capítulos de libros, resúmenes en congresos y artículos de divulgación científica. En 2007, la Real Sociedad Española de Historia Natural lo designó Socio Honorario como reconocimiento a sus relevantes contribuciones a las Ciencias Naturales; en 2006 obtuvo la "Cátedra Patrimonial Enrique Beltrán Castillo" de la División de Ciencias Biológicas y de la Salud en el Departamento del Hombre y su Ambiente de la UAM-Xochimilco y recibió Diploma y Medalla "Alfonso L. Herrera" al mérito en Ecología y Recursos Naturales que otorga el Instituto Mexicano de Recursos Naturales Renovables, A. C, en 1992.
Miriam Carreño De la Vega
Miriam Carreño de la Vega es egresada de la Licenciatura en Biología por parte de la Universidad Nacional Autónoma de México. Durante sus estudios de licenciatura incursionó en la investigación de las evidencias de depredación en moluscos marinos fósiles.
Su interés por el tema de la durofagia en el registro fósil surgió al llevar a cabo la investigación correspondiente, para obtener el título en la licenciatura de Biología. Se graduó en 2014 y actualmente labora en la Comisión Nacional de Arbitraje Médico en la Dirección General de Calidad e Informática.

