










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkTIP. Revista especializada en ciencias químico-biológicas
versão impressa ISSN 1405-888X
TIP vol.17 no.2 Ciudad de México Dez. 2014
Artículo original
Análisis del papel de los caracteres foliares de Cnidoscolus (Euphorbiaceae) en la defensa contra herbívoros y patógenos
Assessing the role of leaf traits against herbivores and pathogens in Cnidoscolus (Euphorbiaceae)
Daniel Torres-González y Graciela García-Guzmán*
Lab. de Ecología de Poblaciones Vegetales, Depto. de Ecología Funcional, Instituto de Ecología, Universidad Nacional Autónoma de México. Ciudad Universitaria, Deleg. Coyoacán, C.P. 04510, México, D.F. E-mail: *mggarcia@ecologia.unam.mx
Artículo recibido el 08 de abril de 2014.
Aceptado el 15 de agosto de 2014.
Resumen
Se analizó el papel de algunos caracteres foliares (área específica, grosor, densidad de tricomas, contenido hídrico y producción de látex) de tres especies de Cnidoscolus (Euphorbiaceae) en la defensa contra insectos herbívoros y hongos patógenos. Se examinó el daño foliar en adultos de las especies arbóreas C. spinosus y C. megacanthus y de la especie herbácea C. urens, en poblaciones localizadas en la Reserva de la Biosfera de Chamela-Cuixmala (Jal.) y en la región de Huatulco-Zimatán (Oax.). También se analizó el daño en las plántulas de C. spinosus. Además se evaluaron los siguientes caracteres foliares: área específica, grosor, densidad de tricomas en el haz y en el envés, contenidos hídrico y de látex. En las tres especies el daño foliar causado por herbívoros forrajeros fue mayor que el causado por patógenos. La especie en la que se registró menor daño foliar fue C. spinosus, aunque el nivel de daño en plántulas fue mayor que en los adultos. Las especies arbóreas presentaron mayores niveles de daño por forrajeros que la herbácea. Sólo en las plántulas de C. spinosus se encontró que a mayor grosor foliar hubo más daño por herbívoros. Los resultados sugieren que los caracteres foliares evaluados son poco importantes en la defensa de las especies estudiadas.
Palabras Clave: Herbivoría, látex, patógenos, selva baja caducifolia, tricomas.
Abstract
The role of some leaf characters (specific area, thickness, trichome density, water content and latex production) on the defence against insect herbivores and fungal pathogens was analyzed in three species of Cnidoscolus (Euphorbiaceae). Foliar damage was examined in adults of the tree species C. spinosus and C. megacanthus, and the herbaceous species C. urens, in populations located in the Biosphere Biological Reserve of Chamela-Cuixmala (Jal.) and in the Huatulco-Zimatán region (Oax.), Mexico. Seedlings of C. spinosus were also analysed. We assessed the following leaf traits: specific area, thickness, density of trichomes on both sides of the leaf, as well as latex and hydric content. In the three Cnidoscolus species, levels of damage caused by grazer insects were higher than the ones caused by pathogens. The lowest level of foliar damage was detected in C. spinosus, but the levels of damage in seedlings were higher than those in the adults. The two tree species showed higher levels of damage by grazers than the herbaceous species. We only detected a direct significant relation between leaf thickness and herbivory levels in seedlings of C. spinousus. No other significant relations were found. This study suggests that the assessed leaf characters do not seem to play a key role in defence of these three Cnidoscolus species.
Key Words: Herbivory, latex, pathogens, seasonal dry forest, trichomes.
Introducción
Entre las interacciones bióticas que tienen un mayor impacto en las comunidades naturales se encuentra la herbivoría por insectos1, ya que los herbívoros consumen más del 20% de la productividad primaria neta anua12,3, pueden afectar la riqueza y la abundancia relativa de las especies de plantas, y promover la heterogeneidad espacial4. Entre los herbívoros resaltan los insectos por su gran importancia en el funcionamiento de los ecosistemas y por su diversidad1. Por su parte, las plantas poseen mecanismos que les permiten eludir, repeler, retardar y restringir el daño causado por sus depredadores; estos mecanismos se han considerado como un factor clave para la codiversificación de los insectos herbívoros y las plantas5. Las plantas responden al ataque de sus depredadores mediante dos estrategias defensivas: la resistencia y la tolerancia. La resistencia involucra características encaminadas a limitar el daño que le provocan herbívoros y microorganismos patógenos, en tanto que la tolerancia se refiere a la capacidad que tienen éstas de reducir las pérdidas en su adecuación, aunque no necesariamente las tasas de encuentro con sus depredadores6,7.
Entre las principales características de las plantas que afectan a los depredadores y parásitos se encuentran las defensas constitutivas como las espinas, tricomas y metabolitos secundarios que afectan el desempeño y la adecuación de sus enemigos naturales8. Pero las plantas también pueden activar su sistema inmunológico y disparar defensas inducidas cuando los receptores de reconocimiento de las plantas captan patrones moleculares asociados a patógenos u otros depredadores, como la flagelina, lipopolisacáridos, peptidoglicano, β-glucano y quitina9-11
Las interacciones entre plantas, herbívoros y patógenos son complejas, ya que en ellas intervienen una gran cantidad de factores bióticos y abióticos12. La cantidad y tipo de daño ocasionado por los herbívoros y patógenos varía en función de la edad de la planta, su historia de vida, el tipo de vegetación y las condiciones macro y microambientales13-15. Estas variables pueden ocasionar diferentes patrones de virulencia y resistencia en las poblaciones16.
En las selvas bajas caducifolias durante una gran parte del año la gran mayoría de las plantas pierden las hojas, provocando que el daño que reciben por parte de los depredadores se concentre en la temporada de lluvias17. El género Cnidoscolus resulta interesante debido a que sus especies poseen tricomas altamente urticantes y látex, características que podrían actuar como mecanismos defensivos; además, en este género se registran especies con diferentes formas de crecimiento (arbóreas, arbustivas y herbáceas)17, lo cual posibilita el estudio del papel que juegan los tricomas, el látex y otros posibles mecanismos de defensa en especies con distintas historias de vida. Actualmente hay pocos estudios que hayan evaluado la interacción entre los caracteres defensivos de las plantas y el daño ocasionado por herbívoros y patógenos en este tipo de vegetación, por ello en este estudio se explora la relación entre la magnitud del daño provocado por insectos herbívoros y hongos patógenos con algunos caracteres defensivos (tricomas urticantes y látex), y con caracteres foliares que se encuentran relacionados con la calidad nutricional de la hoja (contenido hídrico, grosor y área foliar específica) en tres especies del género Cnidoscolus: C. megacanthus, C. spinosus y C. urens. Adicionalmente, en C. spinosus se hace la comparación de estos rasgos entre etapas ontogenéticas (plántula y adulto).
Metodología
El estudio se realizó en la reserva de la Biosfera Chamela-Cuixmala, Jalisco y en la región de Huatulco y Zimatán, Oaxaca. En la Reserva de la Biosfera Chamela-Cuixmala se estudió C. spinosus y en la región de Huatulco y Zimatán, se analizaron C. megacanthus y C. urens.
El género Cnidoscolus (Euphorbiaceae) es endémico de América y contiene a unas 50 especies, principalmente distribuidas en México y Brasil, aunque también en el Caribe y Sudamérica18. Uno de los caracteres más evidentes en Cnidoscolus es la presencia de tricomas urticantes, que pueden llegar a medir hasta 1.3 cm; éstos parecen ser un mecanismo de defensa, ya que al contacto con la piel producen una muy fuerte irritación19,20.
Cnidoscolus megacanthus21 Breckon es un árbol o arbusto, de 8 a 14 cm de diámetro a la altura del pecho y de 3-8 m de altura; el tronco y las ramas presentan abundantes acúleos de 5 a 12 mm rematados en una seta urticante. Las hojas presentan un peciolo glabro y cilíndrico, el haz es ligeramente cóncavo, armado con tricomas urticantes abundantes o esparcidos, de entre 2.5 y 5 mm, sostenidos sobre un pedúnculo cilíndrico.
Cnidoscolus spinosus22 Lundell es un arbusto arborescente o árbol pequeño de hasta 5 m de altura; las ramas son largas, glabras y están armadas con tricomas rígidos tipo espina de hasta 1.3 cm. El peciolo es grueso, largo y con pocos tricomas urticantes, con una glándula larga y carnosa en el ápice del peciolo. La lámina foliar mide entre 9-20 cm de largo y de 12-29 cm de ancho, con 3-5 lobos primarios, las hojas jóvenes poseen gran cantidad de tricomas urticantes en el haz, las hojas maduras contienen menores cantidades, o no los poseen.
Cnidoscolus urens23 (L.) Arthur es una herbácea anual que puede alcanzar hasta 2 m de alto, a veces leñosas, con tricomas urticantes tanto en las hojas como en las ramas, pero sin espinas leñosas.
Entre septiembre y octubre de 2011 se muestrearon hojas de las tres especies de plantas en su estado adulto, de C. spinosus también se incluyó el análisis de las plántulas. Para cada especie, o etapa ontogenética, se ubicaron tres poblaciones distantes entre sí. Las poblaciones de C. spinosus se localizaron en la reserva de la Biosfera Chamela-Cuixmala, Jalisco, en tanto que las de C. megacanthus y C. urens se ubicaron en la región de Huatulco y Zimatán, Oaxaca, todas las poblaciones se encontraron en selva baja caducifolia.
En cada población se ubicaron 30 individuos y de cada individuo se obtuvieron de 3 a 30 hojas maduras, dependiendo de la cantidad que presentara la planta. Se tomaron fotos de todas las hojas y las imágenes fueron procesadas con el programa Winfolia (Regent Instruments Canada Inc., 2007) para obtener el área foliar total, así como el área dañada por hongos patógenos e insectos herbívoros. El daño provocado por insectos herbívoros fue clasificado de la siguiente manera: la remoción de partes de la hoja fue considerada como un daño por forrajeros y el daño en el mesófilo de la hoja fue clasificado como daño por minadores).
A tres hojas de cada individuo se les midieron los siguientes caracteres: contenido hídrico, área foliar específica, grosor, contenido de látex y densidad de tricomas urticantes por cm2 en el haz y el envés. Para obtener el contenido hídrico, las hojas fueron pesadas en el campo (peso fresco) y posteriormente secadas y repesadas (peso seco) y se calculó como: 100*(1-(peso seco/peso fresco)). El área foliar específica (AFE) es una medida asociada de manera negativa con la rigidez de la hoja, así como con su densidad y grosor24,25, se calculó como: área de la hoja (cm2)/ peso seco (g). Para obtener la densidad de tricomas urticantes por cm2 en el haz y envés de las hojas, con un microscopio estereoscópico se contó la cantidad de tricomas en áreas de 1 a 4 cm2 en tres partes de la hoja: porción media del lobo central, y a ambos lados de la base de la lámina, estas medidas fueron escaladas a 1 cm2 y promediadas. El grosor de la hoja se encuentra relacionado de manera negativa con la tasa fotosintética y el crecimiento, y de manera positiva con el costo energético de la hoja24, este carácter se midió en la hoja fresca con un micrómetro digital Mitutoyo (mod. 293-330). Para medir el contenido de látex se realizó un procedimiento similar al usado por Agrawal y Fishbein26. Se utilizaron círculos de papel filtro de 1 cm de radio, estos fueron pesados en microgramo; posteriormente, en el campo, al realizar el corte de las primeras tres hojas el látex que escurría del peciolo se colectó en los círculos de papel filtro, los cuales fueron secados y repesados para obtener el peso del látex en miligramos.
Análisis estadístico
En cada especie se buscó probar la relación entre los distintos caracteres foliares y la magnitud (proporción de área foliar dañada) del daño por hongos patógenos e insectos herbívoros. Se realizaron regresiones logísticas múltiples, se usó el criterio de eliminación regresiva. Debido a la subdispersión de los residuales, los parámetros de la regresión se obtuvieron a partir de cuasi verosimilitud, se utilizó la función de enlace logit.
Adicionalmente, para la población de Zimatán, de la especie C. megacanthus, se probó si los caracteres foliares influían en la incidencia del daño (proporción de hojas dañadas por individuo) por hongos patógenos, para lo cual también se realizó una regresión logística múltiple usando el criterio de eliminación regresiva. Los parámetros del modelo se obtuvieron a partir de cuasi verosimilitud, se utilizó la función de enlace logit. Únicamente en esta población se probó dicha relación, debido a que en el resto hubo gran cantidad de individuos con porcentajes muy bajos (cero, o cercanos a cero) de área foliar dañada. En esta misma población se analizó si había una asociación entre la magnitud del daño causado por insectos herbívoros y el provocado por hongos patógenos por medio de una correlación de Spearman.
Se comprobó la diferencia en la magnitud de daño entre poblaciones de una misma especie y etapa ontogenética con la prueba de Kruskal-Wallis. Para verificar la diferencia entre adultos y plántulas de C. spinosus se realizó una prueba de Mann-Whitney-Wilcoxon.
Para describir la similitud de los caracteres foliares de las poblaciones se realizó un dendograma a partir de un cluster jerárquico obtenido por el método de Ward. Con el fin de controlar la covarianza entre los caracteres de las distintas especies, se realizó un análisis de componentes principales, el cluster se hizo con las distancias euclidianas entre los scores obtenidos de los primeros tres componentes, que en conjunto contenían el 92.4% de la varianza. Debido a que los ejes obtenidos en el análisis de componentes principales son ortogonales, se puede calcular la matriz distancias euclidianas entre los puntos. Además, a través de este procedimiento es posible obtener un cluster más robusto, ya que se elimina una porción de la varianza de los datos27.
Todos los análisis se realizaron en el programa R28. Para el análisis de cluster se utilizaron las medias por población de los caracteres foliares y los niveles de daño, para el resto de los análisis se utilizaron las medias por individuo.
Resultados
Daño por hongos patógenos
Las tres especies fueron afectadas por manchas necróticas, obscuras y pequeñas, de las que fueron aislados hongos que no han sido identificados. Cnidoscolus spinosus fue la especie que presentó una menor magnitud de ataque (Fig. 1), tanto los individuos adultos ( =0.05% del área foliar dañada por planta, e.e.=0.0017, n=86) como las plántulas ( =0.02%, e.e.=0.0004, n=88) presentaron niveles de daño muy bajos. No se encontraron diferencias estadísticamente significativas entre las poblaciones de las plántulas (Kruskal-Wallis, χ2=1, ν=2, P=0.6), pero sí entre las poblaciones de los adultos (Kruskal-Wallis, χ2=24.97, ν=2, P < 0.001). Cnidoscolus megacanthus fue la especie que presentó la mayor magnitud de daño por hongos patógenos (Fig. 1), aunque hubo una gran variación entre poblaciones (Kruskal-Wallis, χ2=32.3, ν=2, P < 0.001), una de ellas (Zimatán) presentó un daño bastante superior al resto ( =1.53%, e.e.=0.05, n=30). Cnidoscolus urens presentó niveles de daño promedio bajos y similares entre localidades (Fig. 1; Kruskal-Wallis, χ2=2.1, ν=2, P =0.34).
=0.05% del área foliar dañada por planta, e.e.=0.0017, n=86) como las plántulas ( =0.02%, e.e.=0.0004, n=88) presentaron niveles de daño muy bajos. No se encontraron diferencias estadísticamente significativas entre las poblaciones de las plántulas (Kruskal-Wallis, χ2=1, ν=2, P=0.6), pero sí entre las poblaciones de los adultos (Kruskal-Wallis, χ2=24.97, ν=2, P < 0.001). Cnidoscolus megacanthus fue la especie que presentó la mayor magnitud de daño por hongos patógenos (Fig. 1), aunque hubo una gran variación entre poblaciones (Kruskal-Wallis, χ2=32.3, ν=2, P < 0.001), una de ellas (Zimatán) presentó un daño bastante superior al resto ( =1.53%, e.e.=0.05, n=30). Cnidoscolus urens presentó niveles de daño promedio bajos y similares entre localidades (Fig. 1; Kruskal-Wallis, χ2=2.1, ν=2, P =0.34).
Daño por insectos herbívoros
Todas las especies tuvieron un nivel de daño por herbívoros bajo (Fig. 2). La especie que presentó menor magnitud de daño fue C. urens ( =0.94% de área foliar dañada por planta, e.e.=0.02, n=90 plantas), gran cantidad de los individuos de esta especie (51%) no mostraron ningún daño por herbívoros. En esta especie no se encontraron diferencias estadísticamente significativas entre las poblaciones (Kruskal-Wallis, χ2=0.74, ν=2, P=0.68). Las poblaciones de C. megacanthus presentaron niveles de daño similares (Kruskal-Wallis, χ2=0.57, ν=2, P=0.75), y en general superiores a los de las otras especies ( =5.64%, e.e.=0.04, n=90). Las poblaciones de los adultos de C. spinosus presentaron niveles de daño superiores al de las plántulas (W=2340, P<0.001), aunque la magnitud del daño fue por lo general menor al 10% (Fig. 2). En los adultos de esta especie se encontraron diferencias estadísticamente significativas en los niveles de daño por forrajeros entre las poblaciones (Kruskal-Wallis χ2=12.46, ν=2, P=0.002), pero no en las plántulas (Kruskal-Wallis, χ2=1.75, ν=2, P=0.41). En las plántulas de esta especie 35% de los individuos no presentaron daño, sin embargo, aquí también encontramos cinco individuos con un daño promedio mayor al 20%.
Tanto el porcentaje de área foliar dañada, como el de hojas atacadas por minadores fue muy bajo. En el caso de los adultos de C. spinosus se encontró que sólo el 0.54% de las hojas muestreadas (N=1669) estuvo dañado por este tipo de herbívoro. Igualmente, sólo el 0.20% de las hojas analizadas en las plántulas (N= 499) presentó daño por minador. En C. megacanthus se detectó que el 6% de las hojas (N=1808) presentó este tipo de daño, y de este porcentaje, el 65% de las hojas presentaron entre 0.01 y 0.02% del área foliar dañada. Por último, de C. urens se revisaron 670 hojas y en ninguna de ellas se observó ataque por minador.
Relación entre los caracteres foliares y los niveles de daño
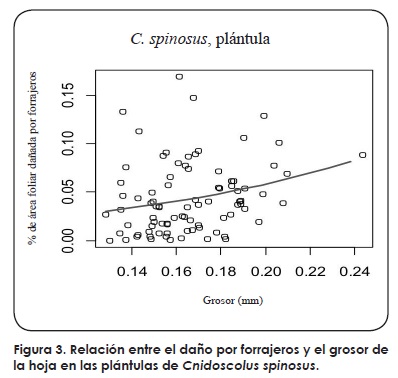
En general no se encontró relación entre los caracteres foliares y los niveles de daño por insectos herbívoros o patógenos. Únicamente en las plántulas de C. spinosus se encontró una relación positiva entre el grosor de la hoja y los niveles de daño por forrajero (logit(magnitud de daño por forrajero)= -4.68+9.49*(grosor); parámetro de dispersión (ф)=0.032; F=5.09, ν=85,86, P=0.026) (Fig. 3). No se encontró una dependencia entre la incidencia del daño por hongos patógenos y los caracteres foliares en la población de Zimatán de C. megacanthus (para todas las variables P>0.1).
Caracteres foliares
El grosor, el contenido hídrico, y el área foliar específica presentaron expresiones relativamente similares entre las especies (Figs. 4.c, d, e). Por otro lado, la densidad de tricomas en el haz y el envés, así como el contenido de látex fueron claramente distintos entre las especies. Las plántulas de C. spinosus y la herbácea C. urens presentaron densidades mayores de tricomas urticantes en el haz de la hoja, mientras que C. megacanthus y los adultos de C. spinosus presentaron densidades muy bajas (Fig. 4.b). En cuanto a las densidades de tricomas en el envés, C. urens fue la especie en la que se registró mayor densidad, en las plántulas de C. spinosus en general fue menor a 3 tricomas/cm2, mientras que C. megacanthus y los adultos de C. spinosus sumamente baja (Fig. 4.a). Se registró la mayor producción de látex en los adultos de C. spinosus y en C. megacanthus, por su parte, las plántulas de C. spinosus y la herbácea C. urens fue menor en proporción (Fig. 4.f).
Tanto en el análisis de cluster, como en el de componentes principales, las poblaciones se agruparon por especie. Las especies presentaron una variabilidad diferente en la expresión de los caracteres; destaca por un lado C. megacanthus, cuyas poblaciones están agrupadas a corta distancia entre sí, lo cual refleja la constancia de la expresión de los caracteres foliares; en el otro extremo se encuentra C. urens que presenta la mayor variabilidad entre poblaciones (Fig.5). En un grado intermedio se encuentran las poblaciones de C. spinosus, especie en la que las plántulas presentan menor variabilidad en los caracteres, mientras que los adultos presentan mayor variación en la expresión de los mismos, aunque menor a la de C. urens. Además, existe una fuerte correlación positiva entre la densidad de tricomas en el haz y la densidad de tricomas en el envés, así como una asociación negativa entre la densidad de tricomas y la cantidad de látex (Fig. 5).

El dendograma obtenido de los scores de los primeros tres componentes principales (Fig. 5) muestra con mayor claridad el grado de similitud entre las poblaciones, destaca la clasificación de las plántulas de C. spinosus y C. urens por un lado, y C. megacanthus y los adultos de C. spinosus por el otro, además de la clasificación de la población de Bocana de C. urens como perteneciente al grupo de las plántulas de C. spinosus.
Discusión
El daño causado por hongos patógenos en todos los casos fue menor al causado por forrajeros. En los ecosistemas tropicales el daño causado por hongos patógenos es bajo29,30. Particularmente, en la región de Chamela el 41% de 168 especies estudiadas, no presentó evidencias de ataque por hongos y sólo el 1% de las especies presentó más del 10% del área foliar dañada, además los niveles de daño se relacionaban de manera negativa con la apertura del dosel, temperatura y humedad (G. García-Guzmán, datos no publ.) lo cual podría explicar por qué los niveles de daño de C. meganathus fueron superiores en la población de Zimatán.
El daño provocado por forrajeros y particularmente por minadores fue bajo en todas las poblaciones. Debido a que las plantas de este género son capaces de tolerar la pérdida de una gran cantidad de tejido foliar31,32, el efecto de los niveles de daño aquí observados sobre la adecuación o supervivencia de las plantas podría ser muy bajo.
Las plántulas de C. spinosus fueron menos atacadas que los adultos de esta especie. Estas diferencias pudieran deberse a que los herbívoros preferirán alimentarse de los adultos, que presentan mayor cantidad de hojas. Además, es posible que las plántulas posean una mayor concentración de metabolitos secundarios u otros caracteres defensivos. Otra posible explicación es que quizás la mayor abundancia de tricomas en la plántula impida no el ataque de insectos, sino la oviposición, como se ha observado en otras especies33,34. Además, se ha sugerido que las plántulas deberían estar más defendidas que los adultos13. En el mismo tenor, el estudio reveló que las especies arbóreas, particularmente C. megacanthus, tuvieron mayor cantidad de daño en comparación con la herbácea. Los estudios sugieren que debido a que las plantas de menor tamaño poseen menor cantidad de hojas, éstas debieran estar mejor protegidas13, más aún por que C. urens es una especie anual, por lo cual el daño foliar pudiera afectar de manera más directa su adecuación.
Conclusiones
Diversas investigaciones han mostrado el papel defensivo de los tricomas35,36. Sin embargo, los resultados aquí reportados no parecen favorecer esa conclusión, esto podría deberse a que los tricomas en Cnidoscolus son grandes en comparación al tamaño de los insectos herbívoros, por lo cual éstos no se verían afectados e incluso pudieran verse beneficiados37. Así, los tricomas pudieran jugar un papel defensivo contra grandes herbívoros, como ha sido sugerido para plantas de la familia Urticaceae38 aunque el impacto de los grandes herbívoros en las selvas bajas caducifolias es en general menor al de los insectos39. Además, se ha observado que existe una asociación negativa entre el contenido de látex y la presencia de tricomas26,40. Cnidoscolus urens presentó mayor densidad de tricomas y menor producción de látex, mientras que en C. spinosus y C. megacanthus se observa el caso contrario. El látex del género Cnidoscolus contiene compuestos tóxicos como linamarina, b-amyrina y glicósidos flavonoides37, sin embargo la calidad del látex como defensa depende tanto de su composición como de su cantidad, por consiguiente es posible que en las poblaciones estudiadas el látex no tenga las características suficientes como para actuar como defensa.
También observamos que ninguno de los caracteres foliares medidos afectó de manera negativa el daño por forrajeros. La relación positiva entre el grosor de la hoja y la herbivoría pudiera deberse al hecho de que este caracter no necesariamente se encuentra asociado a la defensa24. Por lo tanto, los caracteres aquí estudiados parecen ser poco importantes para los insectos y otros herbívoros pequeños que se alimentan de las plantas de este género.
Sin embargo, Cnidoscolus posee otros caracteres que pudieran jugar un papel en la defensa, como la presencia de nectarios en la base de la lámina foliar, que son visitados por hormigas (D. Torres-González, obs. pers.), así como la dureza de las hojas, o la interacción que sostienen las especies de este género con arañas del género Peucetia, cuya presencia se ha demostrado que reduce las tasas de herbivoría41. Además, se conoce que C. chayamansa y C. aconitifolius producen glicósidos cianogénicos42, reportados como compuestos antiherbívoros bastante potentes que pueden llegar a ser tóxicos incluso para los grandes mamíferos, sin embargo, la producción de estas sustancias en otras especies de este género es desconocida.
Para poder comprender las relaciones entre las plantas del género Cnidoscolus y sus depredadores resultaría relevante conocer el impacto que éstos tienen en su desempeño y adecuación, así como la composición química de las hojas y la diversidad de herbívoros y patógenos que consumen a esta planta.
Agradecimientos
Agradecemos a Irma Acosta-Calixto y a Irma Trejo su apoyo en la toma de datos de campo, y también a Guillermo Raúl Castillo Sánchez por su apoyo en los análisis estadísticos. Reconocemos a la Estación de Biología de Chamela y al Mtro. Álvaro Miranda García de la Fundación Ecológica Cuixmala A.C. por las facilidades otorgadas. El proyecto fue financiado por UNAM-PAPIIT (IN205012).
Referencias
1. Agrawal, A.A. Current trends in the evolutionary ecology of plant defence. Funct. Ecol. 25, 420-432 (2011). [ Links ]
2. Coupe, M.D. & Cahill, J.F. Effects of insects on primary production in temperate herbaceous communities: a meta-analysis. Ecol. Entomol. 28, 511-521 (2003). [ Links ]
3. Rasmann, S. & Agrawal, A.A. In defense of roots: a research agenda for studying plant resistance to belowground herbivory. Plant Physiol. 146, 875-880 (2008). [ Links ]
4. Crawley, M.J. Insect herbivores and plant population dynamics. Annu. Rev. Entomol. 34, 531-564 (1989). [ Links ]
5. Ehrlich, P.R. & Raven, P.H. Butterflies and plants: a study in coevolution. Evolution 18, 586-608 (1964). [ Links ]
6. Núñez-Farfán J., Fornoni, J. & Valverde, P.L. The evolution of resistance and tolerance to herbivores. Annu. Rev. Ecol. Evol. S. 38, 541-566 (2007). [ Links ]
7. Oliver T.H., Leather, S.R. & Cook, J.M. Tolerance traits and the stability of mutualism. Oikos 118, 346-352 (2009). [ Links ]
8. Heil, M. Plastic defence expression in plants. Evol. Ecol. 24, 555-569 (2010). [ Links ]
9. Jones, J.D.G. & Dangl, J.L. The plant immune system. Nature 444, 323-329 (2006). [ Links ]
10. Pel, M.J. & Pieterse, C.M. Microbial recognition and evasion of host immunity. J. Exp. Bot. 64, 1237-1248 (2013). [ Links ]
11. Ballaré, C.L. Light Regulation of Plant Defense. Annu. Rev. Plant Biol. (0) (2014). [ Links ]
12. Alexander, H.M. Plant populations, communities, and ecosystems: insights into ecological and evolutionary processes. Plant Disease 94, 492-503 (2010). [ Links ]
13. Coley, P.D. & Barone J.A. Herbivory and plant defenses in tropical forests. Annu. Rev. Ecol. Evol. S 27, 305-335 (1996). [ Links ]
14. Schoonhoven, L.M., van Loon J.J.A. & Dicke, M. Insect-plant biology (Oxford Univ. Press, Nueva York, 2005). [ Links ]
15. García-Guzmán, G. & Morales, E. Life-history strategies of plant pathogens: distribution patterns and phylogenetic analysis. Ecology 88, 589-596 (2007). [ Links ]
16. Burdon, J.J. & Thrall, P.H. Coevolution of plants and their pathogens in natural habitats. Science 324, 755-756 (2009). [ Links ]
17. Bullock, S.H. en Historia Natural de Chamela (ed. F.A., Vega-Rivera, J.H., García-Aldrete, Quesada-Avendaño, M.) 491-498 (Instituto de Biología UNAM, México DF, 2002). [ Links ] Webster, G.L. Cnidoscolus Pohl. Consultado el 2 de febrero de 2013 en Missouri Botanical Garden, URL http://www.tropicos.org (2009). [ Links ]
18. Lutz, O. The poisonous nature of the stinging hairs of Jatropha urens. Science New Series 40, 609-610 (1914). [ Links ]
19. Scheman, A.J. & Conde, A. Contact dermatitis from Cnidoscolus angustidens. Contact Dermatitis 45, 39 (2001). [ Links ]
20. Fernández-Casas, F.J. Cnidoscolorum notulæ (Euphorbiaceaæ). Fontqueria, 55, 481-514 (2008). [ Links ]
21. Lundell, C.L. The genus Cnidoscolus in Mexico: new species and critical notes. B. Torrey Bot. Club 72, 319-334 (1945). [ Links ]
22. Missouri Botanical Garden. Cnidoscolus urens (L.) Arthur. Consultado el 4 de febrero de 2013 en Missouri Botanical Garden, URL http://www.tropicos.org (2009). [ Links ]
23. Vile, D.,É. et al. Specific leaf area and dry matter content estimate thickness in laminar leaves. Ann. Bot. 96, 1129-1136 (2005). [ Links ]
24. Moles, A.T., et al. Putting plant resistance traits on the map: a test of the idea that plants are better defended at lower latitudes. New Phytol. 191, 777-788 (2011). [ Links ]
25. Agrawal, A.A. & Fishbein, M. Plant defense syndromes. Ecology 87, S132-S149 (2006). [ Links ]
26. Husson, F, Josse, J. & Pagès, J. Principal component methods -hierarchical clustering- partitional clustering: why would we need to choose for visualizing data? Technical report, Agrocampus, Applied Mathematics Department. Consultado el 4 de febrero de 2013 en Agro Campus Ouest, URL http://www.agrocampus-ouest.fr (2010). [ Links ]
27. R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org/ (2012). [ Links ]
28. García-Guzmán, G. & Dirzo, R. Patterns of leaf-pathogen infection in the understory of a Mexican rain forest: incidence, spatiotemporal variation, and mechanisms of infection. Am. J. Bot. 88, 634-645 (2001). [ Links ]
29. Gilbert, G.S. Evolutionary ecology of plant disease in natural ecosystems. Annu. Rev. Phytopathol. 40, 13-43 (2002). [ Links ]
30. Parra-Tabla, V., Rico-Gray V. & Carbajal, M. Effect of defoliation on leaf growth, sexual expression and reproductive success of Cnidoscolus aconitifolius (Euphorbiaceae). Plant Ecol. 173, 153-160 (2004). [ Links ]
31. Arceo-Gómez, G., Parra-Tabla, V. & Navarro, J. Changes in sexual expression as result of defoliation and evironment in a monoecious shrub in Mexico: implications for pollination. Biotropica 41: 435-441 (2009). [ Links ]
32. Levin, D.A. The role of trichomes in plant defense. Q. Rev. Biol. 48, 3-15 (1973). [ Links ]
33. Malakar, R. & Tingey, W. M. Glandular trichomes of Solanum berthaultii and its hybrids with potato deter oviposition and impair growth of potato tuber moth. Entomol. Exp. Appl. 92, 249-257 (2003). [ Links ]
34. Lookadoo, S.E. & Pollard, A.J. Chemical contents of stinging trichomes of Cnidoscolus texanus. J. Chem. Ecol. 17, 1909-1916 (1991). [ Links ]
35. Abdala-Roberts, L. & Parra-Tabla, V. Artificial defoliation induces trichome production in the tropical shrub Cnidoscolus aconitifolius (Euphorbiaceae). Biotropica, 37, 251-257 (2005). [ Links ]
36. Stidham, J.A. & Stidham, T.A. Bull nettle (Cnidoscolus texanus) as enemy free space for Orthopterans in Texas, U.S.A. Entomol. News 119, 102-104 (2008). [ Links ]
37. Tuberville, T.D., Dudley, P.G. & Pollard, A.J. Responses of invertebrate herbivores to stinging trichomes of Urtica dioica and Laportea canadensis. Oikos 75, 83-88 (1996). [ Links ]
38. Janzen, D.H. Patterns of herbivory in a tropical deciduous forest. Biotropica 13, 271-282 (1981). [ Links ]
39. Da Silva, D.M. & Batalha, M.A. Defense syndromes against herbivory in a cerrado plant community. Plant Ecol. 212, 181-193 (2011). [ Links ]
40. Arango, A.M., López-Portillo, J., Parra-Tabla, V., Hernández-Salazar, L.T., Morales-Mávil, J.E. & Rico-Gray, V. Effect of the spider Peucetia viridans (Oxyopidae) on floral visitors and seed set of Cnidoscolus multilobus (Euphorbiaceae). Acta Bot. Mex. 100, 1-14 (2012). [ Links ]
41. Kuti, J.O. & Konoru, H.B. Cyanogenic glycosides content in two edible leaves of tree spinach (Cnidoscolus spp.). J. Food Compos. Anal. 19, 556-561 (2006). [ Links ]
42. Vetter, J. Plant cyanogenic glycosides. Toxicon 38, 11-36 (2000). [ Links ]
Información sobre los autores
Daniel Torres-González
Daniel Torres-González realizó sus estudios de Licenciatura en la Facultad de Ciencias, UNAM y se graduó en 2013. Su interés por estudiar los mecanismos defensivos del género Cnidoscolus surgió durante sus primeras visitas a la selva baja caducifolia de la Reserva de la Biosfera de Chamela-Cuixmala, Jal. Actualmente lleva a cabo estudios de Especialización en Estadística Aplicada en la UNAM.
Graciela García-Guzmán
Graciela García-Guzmán realizó estudios de Licenciatura y Maestría en Biología en la Facultad de Ciencias de la UNAM y realizó sus estudios de doctorado en Ecología y Evolución en la Universidad Nacional Australiana (Canberra, Australia). Actualmente labora en el Instituto de Ecología de la UNAM. Sus líneas de investigación se centran en estudiar la ecología y evolución de las interacciones entre hongos patógenos e insectos herbívoros y sus plantas hospederas. Particularmente está interesada en investigar la evolución de los mecanismos defensivos de las plantas contra sus enemigos naturales y el papel que juegan las historias de vida de las plantas y de sus hongos patógenos en los resultados ecológicos y evolutivos de las asociaciones que establecen. Cuenta con varias publicaciones en revistas internacionales de alto nivel sobre estos temas.

