










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencia forestal en México
versión impresa ISSN 1405-3586
Rev. Cien. For. Mex vol.34 no.106 México jul./dic. 2009
Artículo científico
Secuencia de arribo de coleópteros en árboles de Pinus montezumae Lamb. dañados por incendios
Juana Fonseca González1, Celina Llanderal Cázares2, David Cibrián Tovar3, Armando Equihua Martínez2 y Héctor Manuel de los Santos Posadas2
1 Instituto de Ciencias Agropecuarias, Universidad Autónoma del Estado de Hidalgo. Correo–e: fonsecaj@uaeh.edu.mx.
2 Colegio de Posgraduados, Campus Montecillo.
3 División de Ciencias Forestales, Universidad Autónoma Chapingo.
Fecha de recepción: 23 de junio de 2008.
Fecha de aceptación: 20 de noviembre de 2009.
RESUMEN
En los pinos dañados por incendios, varias especies de coleópteros son atraídas por los compuestos volátiles que desprenden los tejidos dañados. Algunos insectos dependen de este factor para su supervivencia debido a que los árboles siniestrados proveen el sustrato para su reproducción y ofrecen menos resistencia a su ataque, al tener presión osmótica inferior. El presente estudio se realizó con la finalidad de conocer la diversidad y la secuencia de llegada de coleópteros a Pinus montezumae afectados por incendios, que se capturaron mediante la aplicación de un pegamento en los troncos. El registro se hizo por fecha de recolecta, se calculó la abundancia relativa de cada grupo y se ajustó a una curva de regresión logística para determinar diferencias entre su patrón de acumulación. Se obtuvieron especímenes de las familias Curculionidae, Buprestidae, Cleridae, Salpingidae, Elateridae, Colydiidae, Bostrichidae y Staphylinidae, la tercera y la primera fueron las más abundantes, 47.5 y 29.2%, respectivamente. De acuerdo a sus hábitos alimentarios, se clasificaron en descortezadores primarios, descortezadores secundarios, barrenadores de madera, depredadores e insectos asociados. Sólo se observaron diferencias significativas entre el patrón de acumulación de descortezadores primarios y barrenadores, con respecto al conjunto de descortezadores, los cuales tienen su máxima acumulación a finales de marzo, mientras que los barrenadores, a principios de abril. Los depredadores acumularon una abundancia relativa muy semejante a la de sus presas, con un máximo a finales de marzo. Se cita una nueva especie para Lechríops como barrenador de madera.
Palabras clave: Chrysobothris inaequalis, Dendroctonus mexicanus, Enoclerus arachnodes, Enoclerus erro, Ips mexicanus, Lechriops.
ABSTRACT
Several beetle species are attracted by the volatile compounds that pines affected by forests fires emit. Some insects depend upon this factor to survive due to the fact that these trees provide the right substrate for their reproduction and offer less resistance to attacks because of a lower osmotic pressure. This study was aimed to evaluate the diversity and sequence of arrival of Coleoptera to Pinus montezumae wildfire damaged trees. Insects were captured with adhesive tangle traps applied directly to the stem. Relative abundance of each species group was calculated for each date of capture and data were fitted to a logistic curve, to differentiate accumulation patterns. The specimens collected included the families Curculionidae, Buprestidae, Cleridae, Salpingidae, Elateridae, Colydiidae, Bostrichidae y Staphylinidae, being Cleridae (47.5%) and Curculionidae (29%) the most abundant. Classified by their feeding habits the insects groups were: primary bark beetles, secondary bark beetles, wood borers, predators and others. Statistical significant differences were only found among the accumulation patterns of primary bark beetles and wood borers. While bark beetles peak in abundance at the end of March the wood borers did it at the beginning of April. Predators exhibit a highly associated pattern to their prey also peaking at the end of March. A new species of wood borer of genus Lechriops is also reported.
Key words: Chrysobothris inaequalis, Dendroctonus mexicanus, Enoclerus arachnodes, Enoclerus erro, Ips mexicanus, Lechriops.
INTRODUCCIÓN
Los incendios ocasionan la degradación de los recursos forestales y anualmente provocan cuantiosas pérdidas económicas y daños irreversibles a los ecosistemas. En México durante el período de 1988 a 1997, afectaron en promedio 246 mil ha al año (SEMARNAP–UACh, 1999), pero en algunos lapsos la superficie ha sido mucho mayor, como en 1998, cuando se quemaron 850 mil ha (Rodríguez, 2007). Los árboles que sobreviven a estos siniestros quedan con algún nivel de "chamuscado" ya sea en la copa o en el tronco, lo que los hace más susceptibles al ataque de plagas y enfermedades (Rodríguez, 1996).
En los pinos dañados por incendios forestales, varias especies de coleópteros son atraídas por los compuestos volátiles que desprenden los tejidos afectados (Suckling et al., 2001; Kelsey y Joseph, 2003). De hecho, algunos insectos dependen de los incendios para su supervivencia debido a que los árboles sometidos a esos eventos proveen el sustrato adecuado para su reproducción (McCullough et al., 1998) y ofrecen menos resistencia a su ataque por tener menor presión osmótica (Chararas, 1978). Se considera que esta atracción es mayor en las primeras semanas y desaparece al segundo año del siniestro (Sullivan et al., 2003); aunque Werner (2002) observó que en el caso de la familia Scolytidae, las poblaciones siguen altas hasta quince años después de la quema.
Los descortezadores son el grupo de coleópteros de mayor importancia por las pérdidas que pueden provocar en el manejo forestal, principalmente el género Dendroctonus (Cibrián et al., 1995). Miller y Patterson (1927) demostraron que Dendroctonus ponderosae Hopkins es atraído a los árboles de Pinus ponderosa Douglas ex C. Lawson dañados por incendio desde una distancia de dos o tres millas. El peligro potencial es que los descortezadores logren acumularse en árboles dañados o muertos, con el consecuente aumento de sus poblaciones, hasta ser capaces de matar individuos sanos adyacentes (Miller y Patterson, 1927; Furniss, 1965). Santoro et al. (2001) registraron que en bosques de Pinus resinosa Aiton, Ips pini Say se incrementó al doble cuatro semanas posteriores a un incendio, pero a los tres meses regresó a los niveles previos al disturbio; mientras que las poblaciones de I. grandicollis Eichh. e I. perroti Swaine se mantuvieron estables. A pesar de que los descortezadores del género Ips se consideran como secundarios, están involucrados en el manchado de la madera y la transmisión de patógenos (Cibrián et al., 1995).
Algunos barrenadores de madera de las familias Buprestidae y Cerambycidae responden directamente al humo (Rogers, 1996), e incluso al resplandor del fuego, como es el caso del género Melanophila (Evans, 1966; citado por McCullough et al., 1998). Estos organismos también buscan los sustratos producidos por el incendio para ovipositar, lo que demostraron Suckling et al. (2001) en la hembra de Arhopalus tristis F. (Cerambycidae) sobre Pinus radiata D. Don. McHugh et al. (2003) determinaron que los bupréstidos y los cerambícidos fueron los insectos más frecuentes en bosques de Pinus ponderosa impactados por el fuego en el sureste de Estados Unidos.
Se ha relacionado la presencia de los depredadores de descortezadores con el nivel de "chamuscado" de los árboles. Kelsey y Joseph (2003) obtuvieron una relación directa entre el nivel de "chamuscado" de Pinus ponderosa y la cantidad de insectos de las familias Cleridae y Trogositidae. Una situación semejante citan Santoro et al. (2001) para Thanasimus dubius F. en rodales quemados de Pinus resinosa.
La colonización de árboles por los escolítidos es seguida por un conjunto de insectos, principalmente depredadores y parasitoides; Stephen y Dahlsten (1976) demostraron que más de 100 especies están asociadas a Dendroctonus brevicomis LeConte y que la diversidad aumenta conforme avanza el ciclo de vida del descortezador. Por lo tanto, el arribo diferenciado de los insectos puede describirse a partir de modelar la acumulación total mediante un modelo logístico.
En México se carece de estudios sobre los principales grupos de insectos que son atraídos hacia coníferas con algún nivel de "chamuscado", así como de su secuencia de llegada. Por ello se plantearon los siguientes objetivos:
a) Identificar los principales descortezadores, barrenadores de madera y depredadores, que llegan a los individuos de Pinus montezumae sometidos a un incendio forestal reciente.
b) Registrar la secuencia de llegada de los diferentes grupos de insectos a los árboles.
MATERIALES Y MÉTODOS
Área de estudio
El trabajo se realizó en el ejido San Rafael Amolucan, municipio Singuilucan, Hgo., en donde a mediados de enero de 2005 ocurrió un incendio forestal en una superficie aproximada de 2 ha. La altitud promedio en el lugar es de 2,330 m y tiene un clima semiseco con lluvias en verano y temperatura anual promedio de 15.7°C (García, 1988). El sitio está asentado en un bosque bajo manejo y corresponde a un área de regeneración de Pinus montezumae, por lo que las características de los árboles son muy homogéneas, con una edad de 16 años, un promedio de 7 m de altura y 20 cm de diámetro normal.
Recolecta e identificación de insectos
El 29 de enero de 2005 se seleccionaron 10 ejemplares con la totalidad del fuste y la copa "chamuscados", pero con yemas terminales vivas, ya que los árboles en esta condición según McHugh et al. (2003) y Wallin et al. (2003) son los más atractivos para Dendroctonus e Ips. Se les aplicó pegamento Stickem®, que ha sido empleado en trampas pegajosas para monitorear la actividad de descortezadores (Stephen y Dahlsten, 1976; Byers et al., 1989) y arácnidos en bosques (Hodge, 1987). El adhesivo se colocó en un área del tronco de 30 x 20 cm, a una altura aproximada de 1.30 m, la que se eligió por ser una dimensión estandarizada y debido a que los ataques de Dendroctonus mexicanus, principal descortezador presente en Hidalgo, se ubican desde la base hasta la punta, en individuos con diámetros menores de 40 cm (Cibrián et al., 1995). Previamente se eliminó la corteza sin llegar al floema, con la finalidad de no aumentar la atracción de los insectos hacia los árboles por la salida de resina. En la superficie tratada se verificó que no hubiera evidencias de ataques previos por descortezadores o barrenadores.
Los insectos atrapados se recolectaron cada dos semanas, del 12 de febrero al 21 de mayo de 2005. En cada ocasión de recolecta se colocaron todos los ejemplares de los 10 árboles en un frasco con thinner como solvente para eliminar los residuos del pegamento y se conservaron en una solución de alcohol etílico al 70% para su posterior registro; además, se contó el número de insectos atrapados por especie o género.
La identificación de los escolítidos se realizó de acuerdo a Wood (1982), la de los cléridos con base en Romero (1993), los estafilínidos por el Dr. Juan Márquez Luna, del Centro de Investigaciones Biológicas de la Universidad Autónoma del estado de Hidalgo (CIB–UAEH), los curculiónidos por el M. en C. Raúl Muñiz Vélez del Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria (SENASICA), SAGARPA y el bupréstido por Richard Wescott del Departamento de Agricultura de Oregon. Los ejemplares se depositaron en la Colección de Coleópteros de la citada institución en el Estado de Hidalgo.
Análisis de datos
Los insectos recolectados se agruparon de acuerdo a sus hábitos alimentarios e importancia en: descortezadores primarios (Grupo 1), descortezadores secundarios (Grupo 2), barrenadores de madera (Grupo 3), depredadores (Grupo 4) e insectos asociados (Grupo 5). Al realizar el primer análisis y no observar diferencias entre los patrones de llegada de descortezadores primarios y secundarios, se decidió integrarlos en un solo grupo (Grupo 6: Grupo 1 + Grupo 2).
Para cada grupo se calculó una abundancia relativa acumulada por fecha de recolecta, de la siguiente manera:

Donde:
Abi = abundancia relativa observada del grupo b en la fecha de colecta i,
ni = número de insectos del grupo b recolectado en la fecha i;
nt = número total de insectos recolectados para el grupo.
La abundancia relativa se comparó entre grupos para conocer si existían diferencias en su patrón de acumulación, para lo cual se buscó una función que lo describiera. Se utilizó un modelo de regresión logística limitado a 1, el cual tiene, entre otras, la ventaja de que al principio de las recolectas (tiempo 0) asume un porcentaje de población inicial base existente antes del evento que dispara su aumento.
La abundancia de cada grupo se ajustó a una curva logística y el modelo utilizado fue:

Donde:
(t) = abundancia relativa calculada de cada grupo de insectos
exp = base de los logaritmos naturales
β1 = coeficientes de regresión
t = fechas de recolecta.
Para establecer si los patrones de acumulación entre grupos son estadísticamente diferentes, se reformuló el modelo 2 de la siguiente forma general:

Donde:
Ij = variable indicadora, que toma el valor 1 si se presenta el j–ésimo grupo y 0 en el caso contrario
Así, y con base en la abundancia del Grupo 6, se pretendió diferenciar la distribución de los grupos de insectos por su abundancia relativa. Donde cada β1j y β2j representa la adicionalidad debida a los otros grupos.
Este modelo se redujo hasta lograr que sólo los parámetros estadísticamente diferentes de cero quedaran incluidos (4).
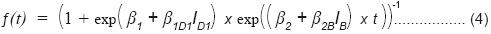
Donde:
β1D1 = parámetro de adicionalidad para el Grupo 1 (descortezadores primarios)
ID1 = variable indicadora para el Grupo 1
β2B = parámetro de adicionalidad para el Grupo 3 (barrenadores de madera)
IB = variable indicadora para el Grupo 3.
Para precisar la fecha en la que se consignó la máxima acumulación de insectos, así como el momento en que ésta empezó a disminuir, se generó la primera derivada del modelo, que de manera general es como se indica:
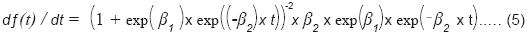
A fin de determinar el patrón de acumulación de los depredadores con respecto a sus presas (descortezadores primarios y secundarios), únicamente se consideraron estos dos grupos en el modelo, y se obtuvo la primera derivada para ubicar los momentos en los que se dieron las mayores acumulaciones de ambas poblaciones.
RESULTADOS Y DISCUSIÓN
Diversidad de Coleópteros
Se capturaron 360 insectos pertenecientes a 20 taxa (Cuadro 1). Estuvieron presentes las familias Cleridae (47.5%), Curculionidae (29.2%) Salpingidae (14.2%), Buprestidae (3.6%) Staphylinidae (2.8%), Bostrichidae (1.1%), Colydiidae (0.8%) y Elateridae (0.8%), que comúnmente se observan en rodales de coníferas afectados por incendios (Werner, 2002; Sullivan et al., 2003; Saint–Germain et al., 2004); aun cuando en esos reportes también se ha identificado a la familia Cerambycidae, en el presente estudio no se registró, posiblemente, debido a que son ejemplares de mayor tamaño y que pudieron escapar de la trampa. Se identificaron cinco grupos de acuerdo a su función en el ecosistema: descortezadores primarios (5.6%), descortezadores secundarios (12.2%), barrenadores de madera (13.9%), depredadores (62.5%) e insectos asociados (5.8%).
Entre los descortezadores, Dendroctonus mexicanus Hopkins es capaz de producir la muerte de árboles sanos, por lo que contribuye al cambio de uso del suelo en los bosques del centro del país (Cibrián et al., 1995). Cabe mencionar que aunque no se colectó a D. valens LeConte, se observaron sus orificios de entrada en tres árboles adyacentes a los muestreados, también afectados por el incendio. Es un insecto común en pinos con daño por fuego (Bradley y Tueller, 2001; Santoro et al., 2001; Kelsey y Joseph, 2003). Otro descortezador presente en el área de estudio, que se considera importante, fue Ips mexicanus Hopkins, ya que infesta tanto árboles vivos, como en proceso de muerte (Furniss y Carolin, 1977; Cibrián et al., 1995). Según Coulson y Witter (1990) este género coloniza y mata árboles sanos cuando tiene niveles altos de población y su presencia confirma su capacidad para atacar individuos lesionados por incendio. Pityophthorus, un descortezador secundario, apareció desde el principio de la recolecta, conun incremento importante en el mes de abril. Algunas especies de Pityophthorus acompañan a descortezadores primarios tales como Dendroctonus, Ips o Scolytus, además colonizan ejemplares derribados, debilitados por sequía o por competencia con otros árboles (Cibrián et al., 1995); así mismo, son atraídos por aquellos que fueron dañados por incendios.
Se capturaron siete ejemplares de Hylastes fulgidus Blackman, la mayoría de ellos en abril y mayo; para Hylurgops incomptus Blandford se recolectó un solo individuo. Ambas especies colonizan tocones o árboles moribundos (Furniss y Carolin, 1977). Apigian et al. (2006) mencionan que estos géneros muestran una marcada preferencia por las áreas sujetas a tratamientos silvícolas en las que se realizan de manera conjunta aclareos y quemas, lo cual es confirmado por el estudio de Kelsey y Joseph (2003), quienes asocian la presencia de Hylurgops porosus LeConte con la severidad del "chamuscado" de Pinus ponderosa, la mayor cantidad se colectó en árboles con la copa completamente quemada, condición que prevaleció en el presente estudio. Estos insectos, considerados secundarios, no provocan un daño inmediato, pero al asociarse con hongos favorecen el debilitamiento gradual de la planta, causándole la muerte en años posteriores al incendio, como lo consignan Sullivan et al. (2003) para Hylastes spp. sobre Pinus palustris Mill.
Del grupo de barrenadores de madera se recolectaron los curculiónidos Gnathotrichus sp., Euplatypus pini Hopkins, una nueva especie del género Lechriops y el bupréstido Chrysobothris inaequalis Waterhouse. El primero, que se observó en el área desde febrero hasta mayo pertenece al grupo de los barrenadores ambrosiales, porque inocula esporas de hongos en las paredes de las galerías lo que produce grandes pérdidas económicas al inducir el manchado de la madera de trozas recién cortadas (Deglow y Borden, 1998); aunque también coloniza árboles moribundos (Wood, 1982). Chénier y Philogène (1989) mencionan que Gnathotrichus es atraído por el etanol que resulta de la fermentación de los tejidos de individuos dañados, por lo que es probable que la llegada del insecto a los árboles "chamuscados" del ejido San Rafael, sea una respuesta a la liberación de este compuesto a la atmósfera. De acuerdo con Kelsey y Joseph (2003), los árboles con la totalidad de la copa quemada incrementan la cantidad de etanol presente, tanto en el floema, como en la albura. Además se recolectó a Euplatypus pini, género que Zanuncio et al. (2002) detectaron en árboles quemados de pino.
Chrysobothris inaequalis fue el único bupréstido registrado con 13 especímenes, tuvo una frecuencia importante (26% del grupo de los barrenadores), lo que coincide con las citas de diversos autores, en el sentido de que esta familia reúne coleópteros considerados pirófilos (Barr, 1985; Dajoz, 2001; Werner, 2002; Sullivan et al., 2003; Saint–Germain et al., 2004). Es trascendente señalar que Ch. inaequalis se registra por primera vez para el estado de Hidalgo. Westcott (1998) menciona que lo recolectó en áreas quemadas del Cofre de Perote a una altitud de 3,000 m, y Westcott y Mudge (2003) lo observaron en el volcán Nevado de Colima a 3,350 msnm, aunque no lograron recolectarlo.
Se obtuvieron ejemplares de una especie nueva del género Lechriops, la cual está en proceso de determinación y en agosto de 2004 emergieron de trozas de árboles pequeños de Pinus montezumae muertos por incendio en un área adyacente al sitio de estudio (Fonseca, 2007). En el presente trabajo representó al 50% de los insectos barrenadores, por lo que se considera que es un barrenador de este pino que acompaña las colonizaciones de descortezadores. Stephen y Dahlsten (1976) registran a Lechriops californicus Le Conte arribando a pinos infestados por Dendroctonus brevicomis Le Conte, cuando ya estaban en desarrollo las larvas del descortezador. Para el estado de Hidalgo sólo se citan a Lechriops aurita Schöenherr y L. lebasii Boheman (Morrone et al., 2002), sin mencionar el hospedante; pero según Hespenheide (2003), los individuos de este género son barrenadores de coníferas, las larvas y pupas se desarrollan bajo la corteza de pinos y un grupo pequeño de especies infesta a Pseudotsuga menziesii (Mirb.) Franco.
Respecto a los depredadores de escolítidos, se reunió a insectos de las familias Cleridae, Salpingidae, Colydiidae y Elateridae. Los cléridos: Enoclerus erro Wolcott y E. arachnodes Klug. representaron 75% del total. Cibrián (1988) consigna sólo a E. arachnodes para el estado de Hidalgo, mientras que a E. erro lo identifica para Jalisco, Michoacán y Tlaxcala. Este último es un depredador de Ips y Dendroctonus mexicanus, su asociación fue demostrada por Romero (1993). Al género Elacatis sp. (Salpingidae), que se observó desde el primer mes de recolecta, Cibrián (1987) señala su presencia en árboles recién infestados por Dendroctonus adjunctus Blandford, pero Rivera y Macías (2001) mencionan que depreda a Ips grandicollis en rodales de Pinus oocarpa Schiede, en Chiapas.
En cuanto a las familias Colydiidae y Elateridae, poco se sabe de su influencia sobre las poblaciones de descortezadores; en el área de estudio sólo constituyeron 2.7% del grupo de depredadores. Sin embargo, en otras localidades parece que los colítidos son importantes depredadores; Cibrián (1987) obtuvo larvas alimentándose de larvas de Dendroctonus adjunctus. Otras especies en el estado larval son ectoparásitos de las larvas y pupas de varios barrenadores de madera (Triplehorn y Johnson, 2005).
Los insectos que se consideraron como asociados pertenecen a las familias Bostrichidae y Staphylinidae; además del curculiónido Amphidees acuminatus Champion. La familia Staphylinidae estuvo representada por las subfamilias Aleocharinae y Staphylininae, que incluyen géneros que se comportan como depredadores (Navarrete et al., 2002).
Secuencia de arribo de coleópteros
Dado que no todos los grupos de insectos tuvieron diferencias en su patrón de acumulación a lo largo del período de colecta, se fueron eliminando los parámetros no significativos hasta lograr diferencias en el patrón de acumulación entre los descortezadores primarios (Grupo 1) y barrenadores de madera (Grupo 3), con respecto al conjunto de descortezadores (Grupo 6) (Cuadro 2).

Al graficar el patrón de acumulación ajustado de estos grupos, se observó que al principio, el conjunto de descortezadores tiene una tasa de acumulación más rápida que los barrenadores de madera, pero después de abril disminuye (Figura 1).

Al calcular la derivada del modelo se determinó que los barrenadores de madera alcanzan su máxima acumulación a principios de abril, al igual que los descortezadores primarios, mientras que el conjunto de descortezadores lo hace a finales de marzo (Figura 2), lo cual coincide con los resultados de Stephen y Dahlsten (1976), quienes observaron que los barrenadores de la familia Curculionidae arriban a los árboles infestados por Dendroctonus brevicomis cuando las larvas del descortezador tienen un desarrollo avanzado. Su llegada está muy relacionada con el bajo vigor de los árboles e incluso con un proceso de descomposición del hospedante. Dajoz (2001) menciona que los descortezadores infestan individuos con una presión de resina de 6 a 8 atmósferas, pero los barrenadores invaden los árboles cuando los valores son más bajos (1.6 atmósferas); la presión normal es de 10 atmósferas y la muerte ocurre a las 4 atmósferas de presión.

Los árboles del área de estudio mostraron un daño severo como lo constata la totalidad de la copa "chamuscada"; según Rodríguez (1996), tal condición causa la muerte a los árboles y los que no mueren adolecen de una ostensible disminución en su crecimiento. Esta situación fue aprovechada, en primera instancia, por los descortezadores que tuvieron una tasa de acumulación más rápida que los barrenadores en los primeros dos meses después del incendio, hecho que coincide con lo señalado por Chararas (1978), en el sentido de que uno de los indicadores más importantes del vigor de un árbol es la presión osmótica del plasma celular y que su incremento a niveles inusuales (producto de sequías), coincide con ataques de coleópteros, tanto escolítidos como bupréstidos.
Sobre el particular, Santoro et al. (2001) mencionan que después de un incendio o quema prescrita, el flujo de resina disminuye, lo cual crea una ventana de oportunidad para la colonización por coleópteros. Al final del periodo de colecta, los descortezadores primarios experimentaron una notoria acumulación debido probablemente a que los árboles tuvieron la capacidad de emitir nuevo follaje y empezaron a restablecer sus funciones fisiológicas a los niveles adecuados y sólo Dendroctonus mexicanus e Ips mexicanus pudieron colonizarlos.
Dendroctonus se considera el género más agresivo porque la mayoría de las especies necesitan colonizar individuos vivos para que su progenie se desarrolle con éxito, al contrario de lo que sucede con Ips que, por lo general, requiere hospedantes recién muertos o que están muy dañados, lo que es común ya que varias de sus especies arriban a los árboles ya colonizados por Dendroctonus, una vez que éste ha superado la resistencia (Coulson y Witter, 1990).
Patrón de acumulación de descortezadores y depredadores
Se confirma que ambos grupos tienen una sincronización en su llegada a los árboles (Figura 3), al parecer producto de la coevolución entre ellos (Birch, 1984; Romero, 1993); sin embargo, se pudo observar que a partir de los primeros días de abril, la abundancia relativa de los depredadores es más alta en comparación con la de los descortezadores.

Al obtener la derivada del modelo, se aprecia que a pesar de que la tendencia es muy similar, la acumulación de depredadores ocurre antes de la correspondiente a los descortezadores y declina más rápido (Figura 4). Esta es una abundancia relativa, ya que si se observan los números absolutos, es notorio que los depredadores superan a sus presas por cinco, lo que coincide con Santoro et al. (2001) en términos de que la abundancia del clérido Thanasimus dubius, depredador específico de Ips perroti e I. grandicollis, se incrementó de 30 a 90% a la siguiente semana de una quema controlada, aunque la población de su presa no aumentó.

Los cléridos son atraídos parcialmente por las feromonas de los descortezadores (Birch, 1984; Romero, 1993; Dahlsten et al., 2004) que al interpretar y utilizarlas, los depredadores tienen la ventaja de llegar en forma simultánea con sus presas a los árboles. Varios estudios han destacado que también usan los volátiles de los individuos arbóreos como las kairomonas para localizar a sus presas, de manera específica los componentes de la resina (Chénier y Philogène, 1989), llega a los hospederos en el transcurso de uno a dos días después de iniciada la infestación (Romero, 1993).
En el caso particular del presente trabajo, es posible que el arribo de los cléridos a P. montezumae haya tenido lugar como respuesta tanto a la feromonas de sus presas, como a los volátiles que emitieron los tejidos dañados de los árboles, a pesar de que en los descortezadores no había las condiciones adecuadas para que aumentaran su población; hecho que no fue posible comprobar. Hacia el final del período de recolecta, los árboles mostraron una recuperación importante, debido a que el incendio se verificó a mediados de febrero cuando las yemas de crecimiento no estaban activas y, según Rodríguez (2007), éstas son más sensibles en dicha etapa y hay mayor contenido de humedad en los tejidos. Además en presencia de P. montezumae, taxón adaptado al fuego, por su corteza gruesa, capacidad de rebrote y autopoda (Rodríguez y Fulé, 2003), razón por la cual y no obstante que el incendio "chamuscó" completamente los árboles, se recuperaron con las primeras lluvias.
Aunque la información sobre el género Elacatis es escasa, se concluyó que el género es un depredador significativo de Ips, ya que representó más del 22% de los depredadores recolectados; además, su abundancia estuvo sincronizada con la de los descortezadores, de la misma forma que los cléridos. De las familias Colydiidae y Elateridae sólo se recolectaron tres especímenes de cada una, por lo que se presume que no tuvieron una influencia relevante sobre la población de los descortezadores.
CONCLUSIONES
La colonización de árboles de Pinus montezumae dañados por incendios se da por un conjunto de coleópteros, de los cuales se identificaron diferentes grupos que explotan diversas partes del hábitat que se forma como consecuencia del siniestro. Los primeros insectos en llegar cambian las condiciones del sustrato, con lo que facilitan los subsiguientes arribos de insectos.
En la secuencia de llegada de los insectos, sólo se observaron diferencias en el patrón entre los grupos de descortezadores primarios y barrenadores de madera, con respecto al resto de los coleópteros. Los barrenadores necesitan colonizar árboles muy debilitados, por lo que preceden a la infestación de los descortezadores.
El patrón de acumulación de insectos depredadores se asoció a su secuencia. En términos absolutos, por cada descortezador se recolectaron cinco insectos depredadores. El uso de la abundancia relativa permite comparar los patrones de acumulación de los diferentes grupos de insectos, aun cuando las densidades absolutas variaron ampliamente.
REFERENCIAS
Apigian, K. O., D. L. Dahlsten and S. L. Stephens. 2006. Fire and fire surrogate treatment effects on leaf litter arthropods in a Western Sierra Nevada mixed–conifer forest. Forest Ecology and Management 221 (1–3): 110–122. [ Links ]
Barr, W. F. 1985. An overview of the Mexican Buprestidae with a listing of Chrysobothris species (Coleoptera). In: Sociedad Mexicana de Entomología (Comp.): Memoria de los Simposia Nacionales de Parasitología Forestal II y III. Instituto Nacional de Investigaciones Forestales. Publicación Especial No. 46. Cuernavaca, Mor. México. pp. 69–75. [ Links ]
Birch, M. C. 1984. Aggregation in bark beetles. In: Bell, W. J. and R. T. Cardé (Eds.): Chemical Ecology of Insects. Chapman and Hall Ltd. London, UK. pp. 331–353. [ Links ]
Bradley, T. and P. Tueller. 2001. Effects of fire on bark beetle presence on Jeffrey pine in the Lake Tahoe Basin. Forest Ecology and Management 142 (1–3): 148–153. [ Links ]
Byers, J. A., A. Olle and J. Löfqvist. 1989. Effective attraction radius: a method for comparing species attractants and determining densities of flying insects. Journal of Chemical Ecology 15 (2): 749–765. [ Links ]
Chararas, C. 1978. La presión osmótica de las especies forestales y sus relaciones con los insectos xilófagos. In: Pesson, P. Ecología Forestal. El bosque: clima, suelo, árboles, fauna. Mundi–Prensa. Madrid, España. pp. 207–229. [ Links ]
Chénier J., V. R. and B. J. R. Philogène. 1989. Field responses of certain forest Coleoptera to conifer monoterpenes and ethanol. Journal of Chemical Ecology 15: 1729–1745. [ Links ]
Cibrián T., D. 1987. Estudios sobre la biología y disposición espacial del descortezador de pinos Dendroctonus adjunctus Blandf. (Coleóptera: Scolytidae). Tesis de Maestría en Ciencias. Centro de Entomología y Acarología, Colegio de Posgraduados. Montecillo, Edo. de Méx. México. pp. 67–70. [ Links ]
Cibrián T., D. 1988. Aspectos ecológicos del depredador Enoclerus arachnodes. In: Sociedad Mexicana de Entomología (Comp.): Memorias del XXIII Congreso Nacional de Entomología. ICI–BAYER–SARH–UMSNH. Morelia, Mich. México. 199 p. [ Links ]
Cibrián T., D., J. T. Méndez–Montiel, R. Campos–Bolaños, H. O. Yates III y J. Flores–Lara. 1995. Insectos Forestales de México/Forest Insects of Mexico. Universidad Autónoma Chapingo. SARH, Subsecretaría Forestal y de Fauna Silvestre México. USDA Forest Service, Natural Resources Canada. Comisión Forestal de América del Norte–FAO. Pub. # 6. Chapingo, Edo. de Méx., México. 453 p. [ Links ]
Coulson, R. N. y J. A. Witter. 1990. Entomología Forestal. Ecología y Control. Editorial LIMUSA. México, D. F. México. 751 p. [ Links ]
Dahlsten, E. L., D. L. Rowney, A. B. Lawson, N. Erbilgin and K. F. Raffa. 2004. Attraction of Ips pini (Coleoptera: Scolytinae) and its predators to natural attractants and synthetic semiochemicals in Northern California: Implications for population monitoring. Environmental Entomology 33 (6): 1554–1561. [ Links ]
Dajoz, R. 2001. Entomología Forestal. Los insectos y el bosque. Mundi–Prensa. Madrid, España. 548 p. [ Links ]
Deglow, E. K. and J. H. Borden. 1998. Green leaf volatiles disrupt and enhance response to aggregation pheromones by the ambrosia beetle, Gnathotrichus sulcatus (Coleoptera: Scolytidae). Canadian Journal of Forest Research 28: 1697–1705. [ Links ]
Fonseca G., J. 2007. Ocurrencia de insectos descortezadores en bosques dañados por incendios. Tesis de Doctorado en Ciencias. Instituto de Fitosanidad. Colegio de Postgraduados. Montecillo, Edo. de Méx. México. 62 p. [ Links ]
Furniss, M. M. 1965. Susceptibility of fire–injured Douglas–fir to bark beetle attack in Southern Idaho. Journal of Forestry 63: 8–11. [ Links ]
Furniss, R. L. and V. M. Carolin. 1977. Western Forest Insects. U. S. Department of Agriculture. Miscellaneous Publication No. 1339. Washington, DC. USA. 654 p. [ Links ]
García, E. 1988. Modificaciones al sistema de clasificación climática de Köppen (para adaptarla a las condiciones de la República Mexicana). 4a ed. Instituto de Geografía, Universidad Nacional Autónoma de México. México, D. F. México. 121 p. [ Links ]
Hespenheide, H. A. 2003. New Lechriops species for the United States (Coleoptera: Curculionidae: Conoderinae). The Colleopterist's Bulletin 57: 345–352. [ Links ]
Hodge, M. A. 1987. Macrohabitat selection by the orb weaving spider, Micrathena gracilis. Psyche 94: 347–361. [ Links ]
Kelsey, R. G. and G. Joseph. 2003. Ethanol in ponderosa pine as an indicator of physiological injury from fire and its relationship to secondary beetles. Canadian Journal of Forest Research 33 (5): 870–884. [ Links ]
McCullough, D. G., R. A. Werner and D. Neumann. 1998. Fire and insects in northern and boreal forest ecosystems of North America. Annual Review of Entomology 43: 107–127. [ Links ]
McHugh, C., T. E. Kolb and J. L. Wilson. 2003. Bark beetle attacks on ponderosa pine following fire in northern Arizona. Environmental Entomology 32: 510–522. [ Links ]
Miller, J. M. and J. E. Patterson. 1927. Preliminary studies on the relation of fire injury to bark–beetle attack in western yellow pine. Journal of Agricultural Research 31 (7): 597–613. [ Links ]
Morrone, J. J., R. Muñiz, J. Asiain y J. Márquez. 2002. Lista de las especies de Curculionoidea (Insecta: Coleoptera) depositadas en la colección del Museo de Zoología "Alfonso L. Herrera", Facultad de Ciencias (MZFC), Universidad Nacional Autónoma de México. Acta Zoológica Mexicana (nueva serie) 87: 147–165. [ Links ]
Navarrete H., J. L., A. F. Newton, M. K. Thayer, J. S. Ashe y D. S. Chandler. 2002. Guía ilustrada para los géneros de Staphylinidae (Coleóptera) de México. Universidad de Guadalajara/CONABIO. México, D. F. México. 401 p. [ Links ]
Rivera G., M. L. y J. E. Macías S. 2001. Respuesta en condiciones de campo de Dendroctonus frontalis, Ips grandicollis (Coleóptera: Scolytidae) y sus depredadores a diferentes semioquímicos en el sureste de México. Manejo Integrado de Plagas 61: 85–86. [ Links ]
Rodríguez T., D. A. 1996. Incendios Forestales. Universidad Autónoma Chapingo y Mundi–Prensa. México, D. F. México. 630 p. [ Links ]
Rodríguez T., D. A. 2007. Factores de predisposición. Fuego. In: Cibrián–Tovar, D., D. Alvarado–Rosales y S. E. García–Díaz (Eds.): Enfermedades Forestales de México/ Forest Diseases in México. Universidad Autónoma Chapingo. Comisión Nacional Forestal. International Forestry Staff, Forest Service. Natural Resources Canada. México, D. F. México. pp. 42–50. [ Links ]
Rodríguez T., D. A. and Z. Fulé. 2003. Fire ecology of Mexican pines and a fire management proposal. International Journal of Wildland Fire 12: 23–37. [ Links ]
Rogers,T. J. 1996. Effects of fire on forest insects in western forest ecosystems. In: R. F. Billings and T. E. Nebeker (Eds.). Proceedings: North American Forest Insect Work Conference. Forest Entomology: Vision 20:21. La Grange, TX, USA. pp. 136–137. [ Links ]
Romero, L. 1993. Estudio de depredadores de la familia Cleridae y su relación con Dendroctonus mexicanus (Hopk.). Tesis de Maestría. División de Ciencias Forestales. Universidad Autónoma Chapingo. Chapingo, Edo. de Méx. México. 140 p. [ Links ]
Saint–Germain, M., P. Drapeau and C. Hébert. 2004. Xilophagous insect species composition and patterns of substratum use on fire–killed black spruce in central Quebec. Canadian Journal of Forest Research 34: 677–685 [ Links ]
Santoro, A. E., M. J. Lombardero, M. P. Ayres and J. J. Ruel. 2001. Interactions between fire and bark beetles in an old growth pine forest. Forest Ecology and Management 144 (1/3): 245–254. [ Links ]
Secretaría del Medio Ambiente, Recursos Naturales y Pesca (SEMARNAP) y Universidad Autónoma Chapingo (UACh). 1999. Atlas Forestal de México. México, D. F. México. 102 p. [ Links ]
Stephen, J. A. and D. L. Dahlsten. 1976. The arrival sequence of the Arthropod complex following attack by Dendroctonus brevicomis (Coleoptera: Scolytidae) in Ponderosa pine. Can. Ent. 108: 283–304. [ Links ]
Suckling, D. M., A. R. Gibb, J. M. Daly, X. Chen and E. G. Brockerhoff. 2001. Behavioral and electrophysiological responses of Arhopalus tristis to burnt pine and other stimuli. Journal of Chemical Ecology 27 (6): 1091–1104. [ Links ]
Sullivan, B. T., C. J. Fetting, W. J. Otrosina, M. J. Dalusky and C. W. Berisford. 2003. Association between severity of prescribed burns and subsequent activity of conifer infesting beetles in stands of longleaf pine. Forest Ecology and Management 185 (3): 327–340. [ Links ]
Triplehorn, Ch. A. and N. F. Johnson (Eds.). 2005. Borror and DeLong's introduction to the study of insects. Thomson Brooks/Cole 7th edition. Belmont, CA. USA. pp. 365–467. [ Links ]
Wallin, K. F., T. E. Kolb, K. R. Skov and M. R. Wagner. 2003. Effects of crown scorch on ponderosa pine resistance to bark beetles in Northern Arizona. Environmental Entomology 32 (3): 652–661. [ Links ]
Werner, R. A. 2002. Effect of ecosystem disturbance on diversity of bark and wood–boring beetles (Coleoptera: Scolytidae, Buprestidae, Cerambycidae) in white spruce (Picea glauca (Moench) Voss) ecosystems of Alaska. Research Paper PNW–RP–546. U. S. Departament of Agriculture, Forest Service, Pacific Northwest Research Station. Portland, OR. USA.15 p. [ Links ]
Westcott, R. 1998. Mexico in spring. Buprestis Report No. 36: 5–6. [ Links ]
Westcott, R. and A. Mudge. 2003. The Buprestidae of Mexico. Buprestis Report No. 41: 3–5. [ Links ]
Wood, S. L. 1982. The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae), a taxonomic monograph. Great Basin Naturalist Memoirs 6: 1– 1359 p. [ Links ]
Zanuncio J. C., M. F. Sossai, L. Couto and R. Pinto. 2002. Ocurrence of Euplatypus parallelus, Euplatypus sp. (Col.: Euplatypodidae) and Xyleborus affinis (Col.: Scolytidae) in Pinus sp. in Ribas do Río Pardo, Mato Grosso do Sul, Brazil. Revista Árvore 26 (3): 387–389. [ Links ]

